
Electrophysiological Characteristics of Inhibitive Control for Adults with Different Physiological or Psychological Obesity
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Materials
2.2.1. Self-Measurements
- (1)
- Hunger, Thirst, and Desire to Eat
- (2)
- Dutch Eating Behavior Questionnaire at Restraint Subscale (DEBQ-RS)
- (3)
- Negative-Physical-Self Scale at Fatness Subscale (NPSS-F)
- (4)
- Positive Affect and Negative Affect Schedule (PANAS)
2.2.2. Experimental Procedure
2.2.3. Data Recording and Analysis
- (1)
- Behavior analyses
- (2)
- Heart rate variability analyses
- (3)
- EEG analyses
- (4)
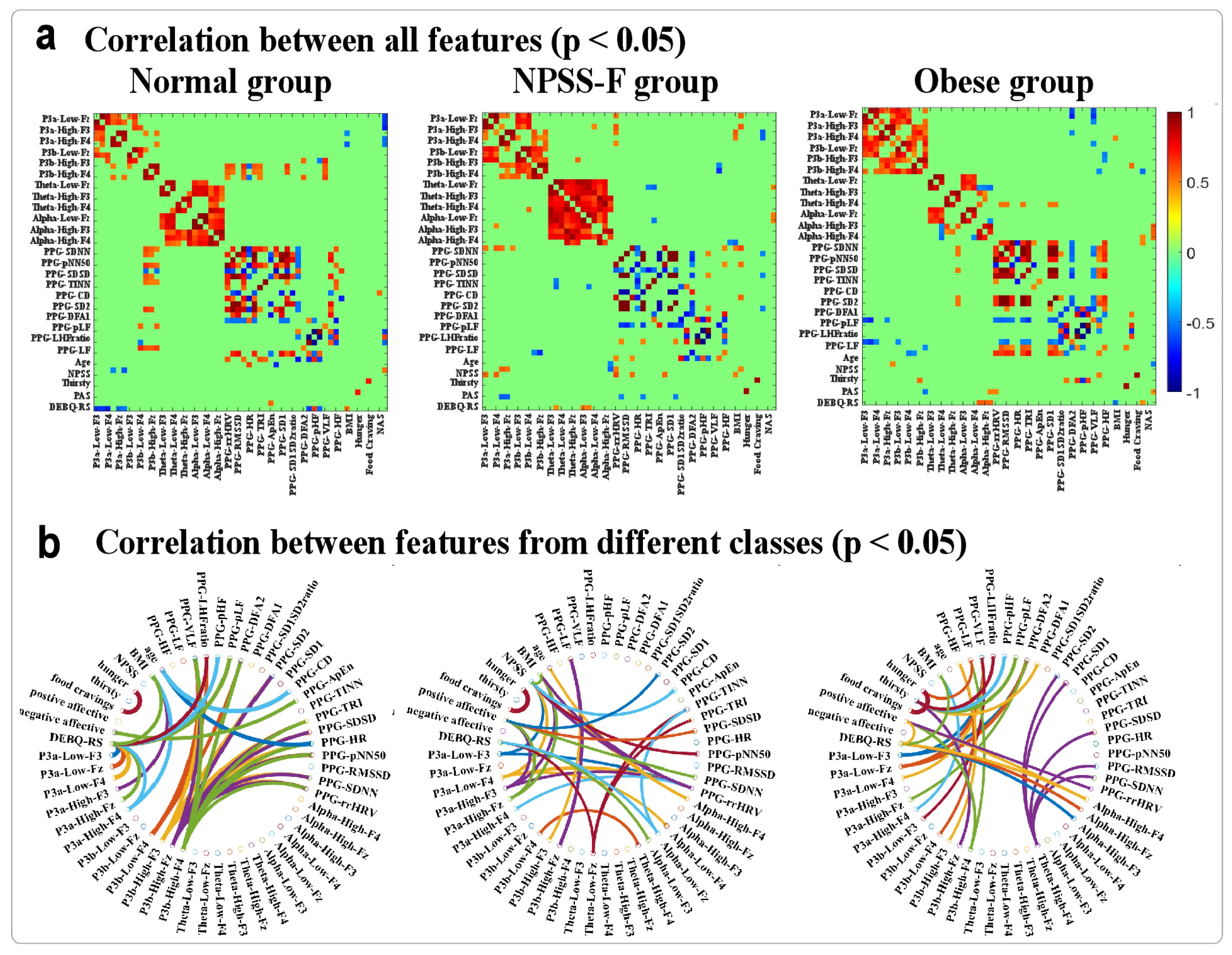
- Correlation analysis
3. Results
3.1. Self-Reported Results
3.2. HRV Results
3.3. EEG Results
4. Discussion
4.1. Inhibitive Control and Working Memory of Obesity
4.2. Brain–Heart Axis
4.3. Limitations and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diamond, A. Executive functions. Annu. Rev. Neurosci. 2013, 64, 135–168. [Google Scholar] [CrossRef] [PubMed]
- Miyake, A.; Friedman, N.P. The nature and organization of individual differences in executive functions: Four general conclusions. Curr. Dir. Psychol. 2012, 21, 8–14. [Google Scholar] [CrossRef]
- Coppin, G.; Nolan-Poupart, S.; Jones-Gotman, M.; Small, D.M. Working memory and reward association learning impairments in obesity. Neuropsychologia 2014, 65, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shields, G.S.; Wu, Q.; Liu, Y.; Chen, H.; Guo, C. The association between obesity and lower working memory is mediated by inflammation: Findings from a nationally representative dataset of U.S. Adults. Brain Behav. Immun. 2020, 84, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Uddin, L.Q. Cognitive and behavioural flexibility: Neural mechanisms and clinical considerations. Nat. Rev. Neurosci. 2021, 22, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, J.; Zhang, X.; Gao, X.; Xu, W.; Chen, H. Overweight adults are more impulsive than normal weight adults: Evidence from erps during a chocolate-related delayed discounting task. Neuropsychologia 2019, 133, 107181. [Google Scholar] [CrossRef]
- Huo, S.; Li, J.; Guo, J.; Yan, H.; Deng, X.; Liu, Y.; Zhao, J. Young adults with negative body image at fatness subscale are more restrained than normal adults during a chocolate discounting task. Int. J. Environ. Res. Public Health 2023, 20, 6122. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Zhao, J.; Li, R.; Gao, Y.; Gao, X. Higher visceral adipose tissue is associated with decreased memory suppression ability on food-related thoughts: A 1-year prospective erp study. Appetite 2023, 191, 107048. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.; Gilbert, S.; Serpell, L. Systematic review: Are overweight and obese individuals impaired on behavioural tasks of executive functioning? Neuropsychol. Rev. 2013, 23, 138–156. [Google Scholar] [CrossRef]
- Liang, J.; Matheson, B.E.; Kaye, W.H.; Boutelle, K.N. Neurocognitive correlates of obesity and obesity-related behaviors in children and adolescents. Int. J. Obes. 2014, 38, 494–506. [Google Scholar] [CrossRef]
- Werthmann, J.; Roefs, A.; Nederkoorn, C.; Mogg, K.; Bradley, B.P.; Jansen, A. Can(not) take my eyes off it: Attention bias for food in overweight participants. Health Psychol. 2011, 30, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Li, Q.; Jiang, Y.; Liu, Y.; Xu, A.; Liu, X.; Chen, H. Do overweight people have worse cognitive flexibility? Cues-triggered food craving may have a greater impact. Nutrients 2022, 14, 240. [Google Scholar] [CrossRef] [PubMed]
- Strack, F.; Deutsch, R. Reflective and impulsive determinants of social behavior. Pers. Soc. Psychol. Rev. 2004, 8, 220–247. [Google Scholar] [CrossRef] [PubMed]
- Meule, A.; von Rezori, V.; Blechert, J. Food addiction and bulimia nervosa. Eur. Eat. Disord. Rev. 2014, 22, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Price, M.; Lee, M.; Higgs, S. Food-specific response inhibition, dietary restraint and snack intake in lean and overweight/obese adults: A moderated-mediation model. Int. J. Obes. 2016, 40, 877–882. [Google Scholar] [CrossRef]
- Batterink, L.; Yokum, S.; Stice, E. Body mass correlates inversely with inhibitory control in response to food among adolescent girls: An fmri study. NeuroImage 2010, 52, 1696–1703. [Google Scholar] [CrossRef] [PubMed]
- Gerdan, G.; Kurt, M. Response inhibition according to the stimulus and food type in exogenous obesity. Appetite 2020, 150, 104651. [Google Scholar] [CrossRef] [PubMed]
- Raji, C.A.; Ho, A.J.; Parikshak, N.N.; Becker, J.T.; Lopez, O.L.; Kuller, L.H.; Hua, X.; Leow, A.D.; Toga, A.W.; Thompson, P.M. Brain structure and obesity. Hum. Brain Mapp. 2009, 31, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, I.A.; Jansen, P.R.; Lamb, H.J. Obesity, brain volume, and white matter microstructure at mri: A cross-sectional UK biobank study. Radiology 2019, 291, 763–771. [Google Scholar] [CrossRef]
- Dong, D.; Chen, X.; Li, W.; Gao, X.; Wang, Y.; Zhou, F.; Eickhoff, S.B.; Chen, H. Opposite changes in morphometric similarity of medial reward and lateral non-reward orbitofrontal cortex circuits in obesity. NeuroImage 2024, 290, 120574. [Google Scholar] [CrossRef]
- Zhao, J.; Long, Z.; Li, Y.; Qin, Y.; Liu, Y. Alteration of regional heterogeneity and functional connectivity for obese undergraduates: Evidence from resting-state fmri. Brain Imaging Behav. 2022, 16, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, Y.; Zhang, W.; Wang, J.; Ji, W.; Manza, P.; Volkow, N.D.; Zhang, Y.; Wang, G.-J. Brain functional and structural magnetic resonance imaging of obesity and weight loss interventions. Mol. Psychiatry 2023, 28, 1466–1479. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jackson, T.; Huang, X. The negative physical self scale: Initial development and validation in samples of chinese adolescents and young adults. Body Image 2006, 3, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Sang, H.-B.; Huang, C.-Z.; Zhou, A.-B. Effect of body-related information on food attentional bias in women with body weight dissatisfaction. Sci. Rep. 2023, 13, 16736. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Luo, Y.; Zhao, J.; Cui, Y.; Chen, H. Peer teasing and restrained eating among chinese college students: The chain mediating role of negative coping styles and negative physical self. Nutrients 2024, 16, 163. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Valentini, E.; Zhang, Z.G.; Liang, M.; Iannetti, G.D. The primary somatosensory cortex contributes to the latest part of the cortical response elicited by nociceptive somatosensory stimuli in humans. NeuroImage 2014, 84, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.E.; Rau, H.; Lutzenberger, W.; Birbaumer, N. Mapping p300 waves onto inhibition: Go/no go discrimination. Electroencephalogr. Clin. Neurophysiol. 1994, 92, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Xu, M.; Zhao, J.; Zhu, Y.; Yang, D. Trust violations affect the emotional working memory updating: An event-related brain potential study. Int. J. Psychophysiol. 2022, 179, 67–76. [Google Scholar] [CrossRef]
- Fjell, A.M.; Walhovd, K.B.; Fischl, B.; Reinvang, I. Cognitive function, p3a/p3b brain potentials, and cortical thickness in aging. Hum. Brain Mapp. 2007, 28, 1098–1116. [Google Scholar] [CrossRef]
- Kok, A. On the utility of p3 amplitude as a measure of processing capacity. Psychophysiology 2001, 38, 557–577. [Google Scholar] [CrossRef]
- Friedman, D.; Squires-Wheeler, E. Event-related potentials (erps) as indicators of risk for schizophrenia. Schizophr. Bull. 1994, 20, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Iacono, W.G.; Carlson, S.R.; Malone, S.M.; McGue, M. P3 event-related potential amplitude and the risk for disinhibitory disorders in adolescent boys. Arch. Gen. Psychiatry 2002, 59, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.L.; Wang, W.; Porjesz, B.; Begleiter, H. Auditory p300 in young alcoholics: Regional response characteristics. Alcohol. Clin. Exp. Res. 1995, 19, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Saint Amour di Chanaz, L.; Pérez-Bellido, A.; Wu, X.; Lozano-Soldevilla, D.; Pacheco-Estefan, D.; Lehongre, K.; Conde-Blanco, E.; Roldan, P.; Adam, C.; Lambrecq, V.; et al. Gamma amplitude is coupled to opposed hippocampal theta-phase states during the encoding and retrieval of episodic memories in humans. Curr. Biol. 2023, 33, 1836–1843.e6. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, J.; Zhou, Y.; Yang, R.; Han, B.; Zhao, Y.; Pang, Y.; Yuan, H.; Chen, H. High-calorie food-cues impair conflict control: Eeg evidence from a food-related stroop task. Nutrients 2022, 14, 4593. [Google Scholar] [CrossRef] [PubMed]
- Bagherzadeh, Y.; Baldauf, D.; Pantazis, D.; Desimone, R. Alpha synchrony and the neurofeedback control of spatial attention. Neuron 2020, 105, 577–587.e5. [Google Scholar] [CrossRef]
- Alamia, A.; Terral, L.; D’Ambra, M.R.; VanRullen, R. Distinct roles of forward and backward alpha-band waves in spatial visual attention. eLife 2023, 12, e85035. [Google Scholar] [CrossRef]
- Palva, S.; Palva, J.M. New vistas for alpha-frequency band oscillations. Trends Neurosci. 2007, 30, 150–158. [Google Scholar] [CrossRef]
- Shaffer, F.; Ginsberg, J.P. An overview of heart rate variability metrics and norms. Front. Public Health 2017, 5, 258. [Google Scholar] [CrossRef]
- Huynh, K. Heartbeat-induced pressure pulsations in cerebral arteries modulate neuronal activity. Nat. Rev. Cardiol. 2024, 21, 218. [Google Scholar] [CrossRef]
- Jammal Salameh, L.; Bitzenhofer, S.H.; Hanganu-Opatz, I.L.; Dutschmann, M.; Egger, V. Blood pressure pulsations modulate central neuronal activity via mechanosensitive ion channels. Science 2024, 383, eadk8511. [Google Scholar] [CrossRef]
- Birch, S.L.; Duncan, M.J.; Franklin, C. Overweight and reduced heart rate variability in british children: An exploratory study. Prev. Med. 2012, 55, 430–432. [Google Scholar] [CrossRef]
- Pavel, A.M.; Mathieson, S.R.; Livingstone, V.; O’Toole, J.M.; Pressler, R.M.; de Vries, L.S.; Rennie, J.M.; Mitra, S.; Dempsey, E.M.; Murray, D.M.; et al. Heart rate variability analysis for the prediction of eeg grade in infants with hypoxic ischaemic encephalopathy within the first 12 h of birth. Front. Pediatr. 2023, 10, 1016211. [Google Scholar] [CrossRef]
- Mather, M.; Thayer, J.F. How heart rate variability affects emotion regulation brain networks. Curr. Opin. Behav. Sci. 2018, 19, 98–104. [Google Scholar] [CrossRef]
- Breitling-Ziegler, C.; Tegelbeckers, J.; Flechtner, H.-H.; Krauel, K. Economical assessment of working memory and response inhibition in adhd using a combined n-back/nogo paradigm: An erp study. Front. Hum. Neurosci. 2020, 14, 322. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.F.; Wang, L.; Pan, A. Epidemiology and determinants of obesity in China. Lancet Diabetes Endocrinol. 2021, 9, 373–392. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Shen, Z.; Gu, W.; Lyu, Z.; Qi, X.; Mu, Y.; Ning, Y.; Meinian Investigator Group. Prevalence of obesity and associated complications in China: A cross-sectional, real-world study in 15.8 million adults. Diabetes Obes. Metab. 2023, 25, 3390–3399. [Google Scholar] [CrossRef]
- van Strien, T.; Frijters, J.E.R.; van Staveren, W.A.; Defares, P.B.; Deurenberg, P. The predictive validity of the dutch restrained eating scale. Int. J. Eat. Disord. 1986, 5, 747–755. [Google Scholar] [CrossRef]
- Bohrer, B.K.; Forbush, K.T.; Hunt, T.K. Are common measures of dietary restraint and disinhibited eating reliable and valid in obese persons? Appetite 2015, 87, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Loughnan, S.A.; Mulgrew, K.E.; Lane, B.R. Attention bias modification produces no changes to appearance-related bias, state or trait body dissatisfaction in nonclinical women. Health Psychol. Open 2015, 2, 2055102915614310. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and validation of brief measures of positive and negative affect: The panas scales. J. Pers. Soc. Psychol. 1988, 54, 1063–1070. [Google Scholar] [CrossRef]
- Vollmer, M. A robust, simple and reliable measure of heart rate variability using relative rr intervals. In Proceedings of the 2015 Computing in Cardiology Conference (CinC), Nice, France, 6–9 September 2015. [Google Scholar]
- Vollmer, M. Hrvtool—An open-source matlab toolbox for analyzing heart rate variability. In Proceedings of the 2019 Computing in Cardiology (CinC), Singapore, 8–11 September 2019. [Google Scholar]
- Delorme, A.; Makeig, S. Eeglab: An open source toolbox for analysis of single-trial eeg dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Y.; Qin, Y.; Liu, Y.; Chen, Q.; Zhao, K.; Long, Z. Electroencephalographic oscillations of alpha and beta rhythms during phrase-guessing procedure. Cogn. Neurodynamics 2023, 17, 1345–1355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.G.; Hu, L.; Hung, Y.S.; Mouraux, A.; Iannetti, G.D. Gamma-band oscillations in the primary somatosensory cortex—A direct and obligatory correlate of subjective pain intensity. J. Neurosci. 2012, 32, 7429–7438. [Google Scholar] [CrossRef]
- Johnson, R. A triarchic model of p300 amplitude. Psychophysiology 1986, 23, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Huang, D.; Ao, H.; Wang, X.; Gao, X. Food cue recruits increased reward processing and decreased inhibitory control processing in the obese/overweight: An activation likelihood estimation meta-analysis of fmri studies. Obes. Res. Clin. Pract. 2020, 14, 127–135. [Google Scholar] [CrossRef]
- Liu, X.; Turel, O.; Xiao, Z.; Lv, C.; He, Q. Neural differences of food-specific inhibitory control in people with healthy vs higher bmi. Appetite 2023, 188, 106759. [Google Scholar] [CrossRef]
- Reyes, S.; Peirano, P.; Peigneux, P.; Lozoff, B.; Algarin, C. Inhibitory control in otherwise healthy overweight 10-year-old children. Int. J. Obes. 2015, 39, 1230–1235. [Google Scholar] [CrossRef]
- Nijs, I.M.T.; Franken, I.H.A. Attentional processing of food cues in overweight and obese individuals. Curr. Obes. Rep. 2012, 1, 106–113. [Google Scholar] [CrossRef]
- Drewnowski, A.; Henderson, S.A.; Shore, A.B. Taste responses to naringin, a flavonoid, and the acceptance of grapefruit juice are related to genetic sensitivity to 6-n-propylthiouracil. Am. J. Clin. Nutr. 1997, 66, 391–397. [Google Scholar] [CrossRef]
- Kakoschke, N.; Hawker, C.; Castine, B.; de Courten, B.; Verdejo-Garcia, A. Smartphone-based cognitive bias modification training improves healthy food choice in obesity: A pilot study. Eur. Eat. Disord. Rev. 2018, 26, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Tahsili-Fahadan, P.; Geocadin, R.G. Heart–brain axis. Circ. Res. 2017, 120, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Strüven, A.; Holzapfel, C.; Stremmel, C.; Brunner, S. Obesity, nutrition and heart rate variability. Int. J. Mol. Sci. 2021, 22, 4215. [Google Scholar] [CrossRef]
- Young, H.A.; Benton, D. Heart-rate variability: A biomarker to study the influence of nutrition on physiological and psychological health? Behav. Pharmacol. 2018, 29, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Eckberg, D.L. Sympathovagal balance. Circulation 1997, 96, 3224–3232. [Google Scholar] [CrossRef] [PubMed]
- Chuangchai, W.; Pothisiri, W. Postural changes on heart rate variability among older population: A preliminary study. Curr. Gerontol. Geriatr. Res. 2021, 2021, 6611479. [Google Scholar] [CrossRef]
- Kumar, P.; Das, A.K.; Prachita; Halder, S. Time-domain hrv analysis of ecg signal under different body postures. Procedia Comput. Sci. 2020, 167, 1705–1710. [Google Scholar] [CrossRef]
- Maggioni, M.A.; Rundfeldt, L.C.; Gunga, H.-C.; Joerres, M.; Merati, G.; Steinach, M. The advantage of supine and standing heart rate variability analysis to assess training status and performance in a walking ultramarathon. Front. Physiol. 2020, 11, 731. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Control Group (M ± SD) N = 16 | NPSS-F Group (M ± SD) N = 17 | Obese Group (M ± SD) N = 18 | F | p |
|---|---|---|---|---|---|
| Age | 19.38 (1.67) | 19.82 (1.47) | 19.78 (1.35) | 0.448 | 0.64 |
| DEBQ-RS *** | 2.41 (0.81) | 3.69 (0.68) | 3.30 (0.82) | 11.86 | <0.001 |
| NPSS-F *** | 1.31 (0.21) | 2.96 (0.33) | 2.97 (0.38) | 151.6 | <0.001 |
| Hunger | 20.63 (16.52) | 32.35 (15.62) | 27.22 (17.76) | 2.04 | 0.14 |
| Thirst | 41.25 (18.57) | 36.47 (10.57) | 32.22 (13.09) | 1.68 | 0.198 |
| Desire to eat | 25.00 (18.26) | 30.59 (17.13) | 24.44 (19.17) | 0.594 | 0.56 |
| PAS | 2.98 (0.73) | 2.72 (0.58) | 3.00 (0.61) | 0.969 | 0.387 |
| NAS | 1.84 (0.72) | 1.96 (0.70) | 1.86 (0.63) | 0.149 | 0.862 |
| BMI *** | 20.30 (1.38) | 21. 40 (1.44) | 26.82 (1.88) | 83.13 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Wan, X.; Lian, J.; Ma, H.; Dong, D.; Liu, Y.; Zhao, J. Electrophysiological Characteristics of Inhibitive Control for Adults with Different Physiological or Psychological Obesity. Nutrients 2024, 16, 1252. https://doi.org/10.3390/nu16091252
Guo J, Wan X, Lian J, Ma H, Dong D, Liu Y, Zhao J. Electrophysiological Characteristics of Inhibitive Control for Adults with Different Physiological or Psychological Obesity. Nutrients. 2024; 16(9):1252. https://doi.org/10.3390/nu16091252
Chicago/Turabian StyleGuo, Jiaqi, Xiaofang Wan, Junwei Lian, Hanqing Ma, Debo Dong, Yong Liu, and Jia Zhao. 2024. "Electrophysiological Characteristics of Inhibitive Control for Adults with Different Physiological or Psychological Obesity" Nutrients 16, no. 9: 1252. https://doi.org/10.3390/nu16091252