
Association between Food or Nutrients and Gut Microbiota in Healthy and Helminth-Infected Women of Reproductive Age from Zanzibar, Tanzania

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethic Statements
2.2. Study Design and Recruitment of Participants
2.3. Faecal Sample Collection and Parasitological Analysis
2.4. Recording and Evaluation of Food and Nutrient Intake
2.5. DNA Extraction, PCR, and Sequencing
- Pro 341F:
- 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNBGCASCAG-3′
- Pro 805R:
- 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACNVGGGTATCTAATCC-3′
2.6. Bioinformatic and Data Analysis
2.7. Statistical Analysis
3. Results
3.1. Characteristics of Participants
3.2. Structure of the Diet of Women of Reproductive Age from Pemba
3.3. Analysis of Nutrient Intakes for Women in Pemba and Unguja
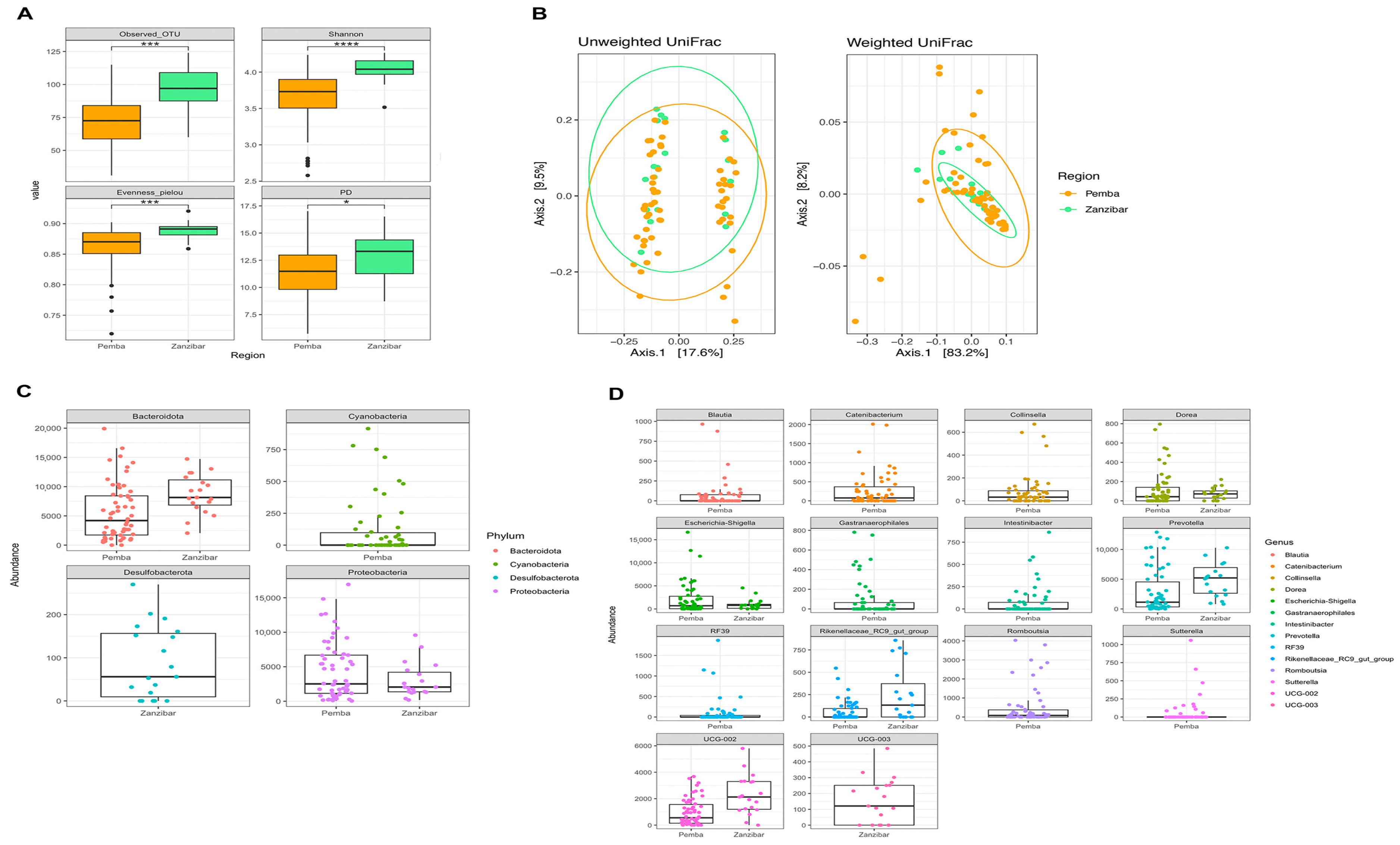
3.4. Women from Pemba and Unguja Harbour Different Gut Microbiota
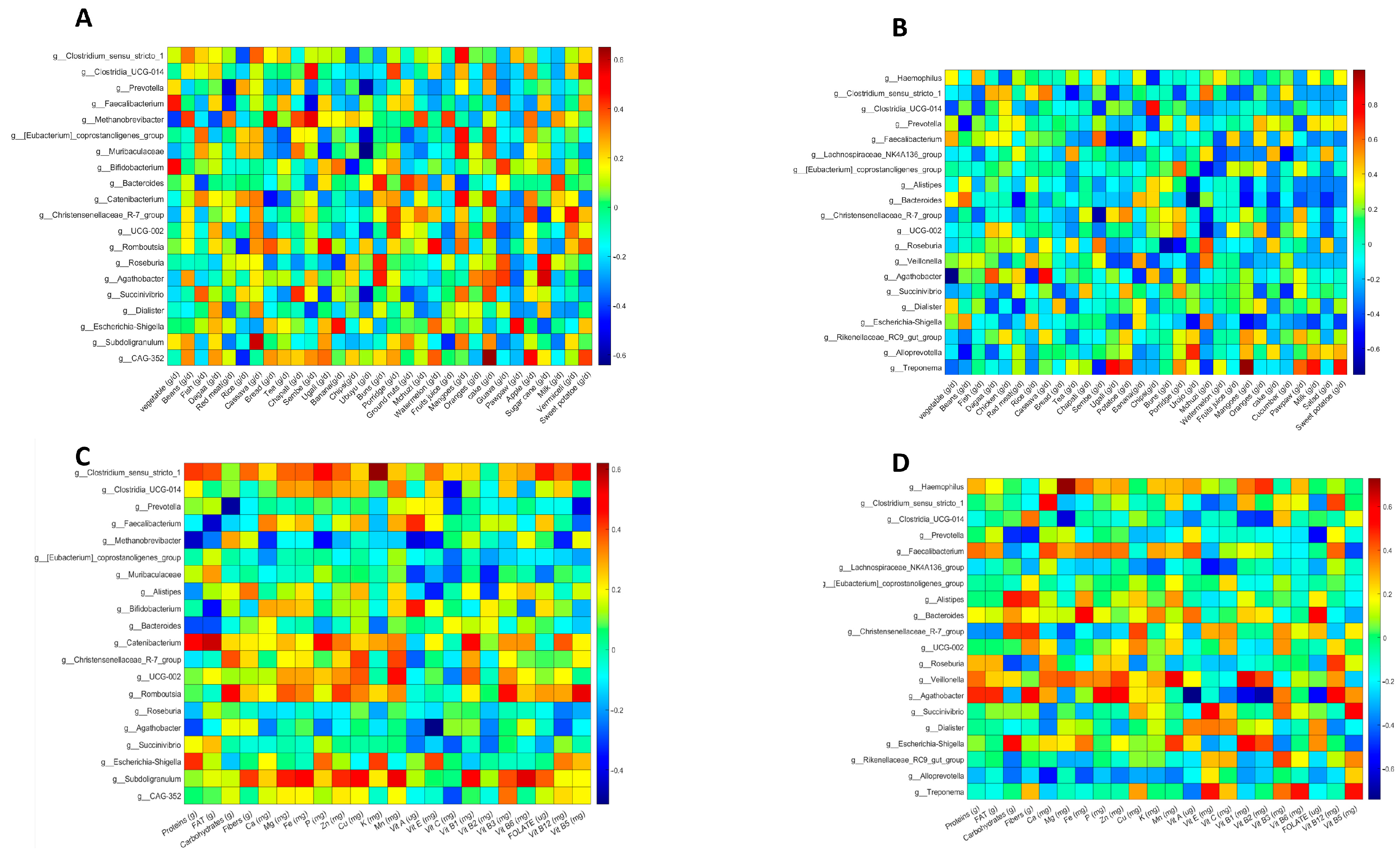
3.5. Correlation Analysis of Foods and Gut Microbiota in Non-Pregnant and Healthy Women from Pemba and Unguja
3.6. Nutrient and Gut Microbiota Correlations in Non-Pregnant and Healthy Women from Pemba and Unguja
3.7. Correlation Analysis of Foods and Gut Microbes in Helminth-Infected Participants
3.8. Correlation Analysis of Nutrients and Gut Microbes in Helminth-Infected Participants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Region | Age (Years) | Weight (kg) | Height (cm) | BMI (kg/m2) | Helminth | Ascaris | Trichuris | EPG |
|---|---|---|---|---|---|---|---|---|---|
| 01M_11A_1 | Pemba | 32 | 52.6 | 152.5 | 22.5 | No | No | No | 0 |
| 01M_11B_1 | Pemba | 32 | 52.6 | 152.5 | 22.5 | No | No | No | 0 |
| 02M_10A_1 | Pemba | 30 | 48 | 150 | 21.33 | No | No | No | 0 |
| 02M_10B_1 | Pemba | 30 | 48 | 150 | 21.33 | No | No | No | 0 |
| 02M_11A_1 | Pemba | 24 | 47 | 156 | 19.31 | No | No | No | 0 |
| 02M_11B_1 | Pemba | 24 | 47 | 156 | 19.31 | No | No | No | 0 |
| 02M_19B_1 | Pemba | 35 | 73.4 | 153.2 | 31.18 | No | No | No | 0 |
| 03M_16A_1 | Pemba | 42 | 67 | 155.4 | 27.74 | No | No | No | 0 |
| 03M_16B_1 | Pemba | 42 | 67 | 155.4 | 27.74 | No | No | No | 0 |
| 03M_3B_1 | Pemba | 23 | 60 | 155.7 | 24.75 | No | No | No | 0 |
| 03M_15B_1 | Pemba | 38 | 83.7 | 151 | 36.7 | No | No | No | 0 |
| 01M1B | Pemba | 30 | 75 | 158 | 30.04 | TT | No | Yes | 60 |
| 01M3A | Pemba | 26 | 45.3 | 158 | 18.14 | No | No | No | 0 |
| 01M4B | Pemba | 34 | 75 | 149 | 33.78 | No | No | No | 0 |
| 01M5B | Pemba | 23 | 63.3 | 163 | 23.82 | No | No | No | 0 |
| 01M7B | Pemba | 20 | 68.3 | 168 | 24.19 | TT | No | Yes | 50 |
| 01M8B | Pemba | 37 | 65 | 154.2 | 27.4 | AS | Yes | No | 40 |
| 01M9B | Pemba | 32 | 68 | 162.5 | 25.91 | No | No | No | 0 |
| 01M10B | Pemba | 27 | 67 | 157 | 27.18 | No | No | No | 0 |
| 01M12B | Pemba | 35 | 67 | 167 | 24.02 | No | No | No | 0 |
| 01M13B | Pemba | 25 | 45.4 | 146.6 | 21.11 | No | No | No | 0 |
| 01M15B | Pemba | 40 | 68 | 151 | 29.82 | AS | Yes | No | 70 |
| 01M16B | Pemba | 27 | 43 | 151.2 | 18.85 | No | No | No | 0 |
| 01M17B | Pemba | 27 | 43 | 155 | 17.89 | No | No | No | 0 |
| 01M21A | Pemba | 27 | 46 | 147 | 21.28 | No | No | No | 0 |
| 01M22B | Pemba | 20 | 60.3 | 162 | 24 | No | No | No | 0 |
| 01M23B | Pemba | 38 | 76 | 158 | 30.44 | No | No | No | 0 |
| 01M25B | Pemba | 25 | 52 | 153 | 22.21 | No | No | No | 0 |
| 01M26B | Pemba | 25 | 67 | 163 | 25.21 | No | No | No | 0 |
| 01M27B | Pemba | 23 | 61 | 158.1 | 24.43 | No | No | No | 0 |
| 02M2A | Pemba | 33 | 82 | 168 | 31.24 | AS/TT | No | No | 60 and 100 |
| 02M3A | Pemba | 35 | 75 | 153 | 32.03 | AS | Yes | No | 110 |
| 02M5A | Pemba | 31 | 45 | 146 | 21.11 | AS | Yes | No | 90 |
| 02M6B | Pemba | 21 | 60 | 154 | 25.3 | TT | No | Yes | 30 |
| 02M7B | Pemba | 23 | 86 | 156 | 35.33 | No | No | No | 0 |
| 02M12B | Pemba | 32 | 49 | 161.6 | 18.9 | AS/TT | No | No | 20 and 40 |
| 02M13B | Pemba | 30 | 56.3 | 156.3 | 23.01 | No | No | No | 0 |
| 02M14B | Pemba | 28 | 77 | 157.3 | 31.23 | No | No | No | 0 |
| 02M15B | Pemba | 30 | 43 | 151 | 18.85 | No | No | No | 0 |
| 02M17B | Pemba | 32 | 50 | 158 | 20.02 | No | No | No | 0 |
| 02M18B | Pemba | 38 | 53 | 153 | 22.64 | No | No | No | 0 |
| 02M20B | Pemba | 40 | 57 | 155.3 | 23.72 | No | No | No | 0 |
| 02M22A | Pemba | 28 | 47 | 153.4 | 20.07 | No | No | No | 0 |
| 02M23B | Pemba | 37 | 42 | 146.6 | 19.43 | No | No | No | 0 |
| 02M24A | Pemba | 27 | 86 | 162.5 | 32.58 | TT | No | Yes | 50 |
| 02M26A | Pemba | 30 | 69 | 160 | 26.95 | AS | Yes | No | 150 |
| 02M27A | Pemba | 22 | 50 | 160 | 19.53 | TT | No | Yes | 180 |
| 02M28A | Pemba | 25 | 54 | 158 | 21.63 | No | No | No | 0 |
| 03M1B | Pemba | 40 | NA | NA | NA | No | No | No | 0 |
| 03M2B | Pemba | 53 | 155.5 | 21.91 | No | No | No | 0 | |
| 03M4B | Pemba | 28 | 79 | 166 | 28.66 | No | No | No | 0 |
| 03M6B | Pemba | 37 | 40 | 147 | 18.51 | AS | Yes | No | 360 |
| 03M7B | Pemba | 45 | 76.5 | 166 | 27.76 | No | No | No | 0 |
| 03M10B | Pemba | 22 | 50 | 162 | 19.05 | No | No | No | 0 |
| 03M11B | Pemba | 27 | 65 | 163 | 24.46 | TT | No | Yes | 170 |
| 03M12A | Pemba | 28 | 38 | 141 | 19.11 | No | No | No | 0 |
| 03M13A | Pemba | 20 | 39.1 | 151 | 17.14 | No | No | No | 0 |
| 03M14A | Pemba | 30 | 56 | 155 | 23.3 | No | No | No | 0 |
| Code | Region | Age (Years) | Weight (kg) | Height (cm) | BMI (kg/m2) | Helminth | Ascaris | Trichuris | EPG |
|---|---|---|---|---|---|---|---|---|---|
| 1M_2 | Unguja | 26 | 44.5 | 148 | 20.31 | No | No | No | 0 |
| 2M_2 | Unguja | 32 | 73 | 158 | 29.24 | No | No | No | 0 |
| 3M_2 | Unguja | 36 | 60.5 | 135 | 33.19 | No | No | No | 0 |
| 4M_2 | Unguja | 24 | 61.7 | 120 | 42.85 | No | No | No | 0 |
| 6M_2 | Unguja | 20 | 35.1 | 136 | 18.98 | No | No | No | 0 |
| 7M_2 | Unguja | 36 | 60.5 | 139 | 31.31 | No | No | No | 0 |
| 8M_2 | Unguja | 23 | 52 | 119 | 36.72 | No | No | No | 0 |
| 9M_2 | Unguja | 27 | 76 | 120 | 52.78 | No | No | No | 0 |
| 11M_2 | Unguja | 45 | 55 | 120 | 38.19 | Yes | No | Yes | Nd |
| 12M_2 | Unguja | 38 | 72 | 121 | 49.18 | No | No | No | 0 |
| 13M_2 | Unguja | 27 | 55 | 126 | 34.64 | Yes | No | Yes | Nd |
| 14M_2 | Unguja | 25 | 58.4 | 137 | 31.12 | No | No | No | 0 |
| 15M_2 | Unguja | 25 | 60 | 154 | 25.3 | No | No | No | 0 |
| 16M_2 | Unguja | 45 | 80 | 159 | 31.4 | No | No | No | 0 |
| 18M_2 | Unguja | 32 | 54 | 153 | 23.07 | No | No | No | 0 |
| 19M_2 | Unguja | 22 | 74 | 174 | 24.44 | No | No | No | 0 |
| 20M_2 | Unguja | 24 | 56 | 134 | 29.84 | No | No | No | 0 |
| Nutrients. Recommended Intakes | % PEMBA | % UNGUJA | % TOTAL | |
|---|---|---|---|---|
| Proteins (g) 50–75 | Normal | 48.07 | 29.41 | 43.48 |
| Above | 36.53 | 58.82 | 40.02 | |
| Below | 15.38 | 11.76 | 14.50 | |
| Fats (g) 33.33–66.66 | Normal | 42.30 | 47.06 | 43.48 |
| Above | 53.84 | 52.94 | 53.62 | |
| Below | 3.85 | 0 | 2.90 | |
| Carbohydrates (g) 275–375 | Normal | 69.23 | 52.94 | 65.21 |
| Above | 5.77 | 5.88 | 5.79 | |
| Below | 25 | 41.18 | 29 |
References
- Morelli, L.; Garbagna, N.; Rizzello, F.; Zonenschain, D.; Grossi, E. In vivo association to human colon of Lactobacillus paracasei B21060: Map from biopsies. Dig. Liver Dis. 2006, 38, 894–898. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Li, S.; Gan, R.-Y.; Zhou, T.; Xu, D.-P.; Li, H.-B. Impacts of Gut Bacteria on Human Health and Diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Tseng, C.H.; Wu, C.Y. The gut microbiome in obesity. J. Formos. Med. Assoc. 2019, 118, S3–S9. [Google Scholar] [CrossRef]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet–microbiota interactions and personalized nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef]
- De Filippis, F.; Vitaglione, P.; Cuomo, R.; Canani, R.B.; Ercolini, D. Dietary Interventions to Modulate the Gut Microbiome—How Far Away Are We From Precision Medicine. Inflamm. Bowel Dis. 2018, 24, 2142–2154. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Seel, W.; Reiners, S.; Kipp, K.; Simon, M.-C.; Dawczynski, C. Role of Dietary Fiber and Energy Intake on Gut Microbiome in Vegans, Vegetarians, and Flexitarians in Comparison to Omnivores—Insights from the Nutritional Evaluation (NuEva) Study. Nutrients 2023, 15, 1914. [Google Scholar] [CrossRef]
- Wu, G.D.; Compher, C.; Chen, E.Z.; Smith, S.A.; Shah, R.D.; Bittinger, K.; Chehoud, C.; Albenberg, L.G.; Nessel, L.; Gilroy, E.; et al. Comparative metabolomics in vegans and omnivores reveal constraints on diet-dependent gut microbiota metabolite production. Gut 2016, 65, 63–72. [Google Scholar] [CrossRef]
- Galié, S.; García-Gavilán, J.; Papandreou, C.; Camacho-Barcía, L.; Arcelin, P.; Palau-Galindo, A.; Rabassa, A.; Bulló, M. Effects of Mediterranean Diet on plasma metabolites and their relationship with insulin resistance and gut microbiota composition in a crossover randomized clinical trial. Clin. Nutr. 2021, 40, 3798–3806. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut Microbiota 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- García-Vega, S.; Corrales-Agudelo, V.; Reyes, A.; Escobar, J.S. Diet Quality, Food Groups and Nutrients Associated with the Gut Microbiota in a Nonwestern Population. Nutrients 2020, 12, 2938. [Google Scholar] [CrossRef]
- Jang, H.B.; Choi, M.-K.; Kang, J.H.; Park, S.I.; Lee, H.-J. Association of dietary patterns with the fecal microbiota in Korean adolescents. BMC Nutr. 2017, 3, 20. [Google Scholar] [CrossRef]
- Maukonen, J.; Saarela, M. Human gut microbiota: Does diet matter? Proc. Nutr. Soc. 2015, 74, 23–36. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Laghi, L.; Gobbetti, M.; Ercolini, D. Unusual sub-genus associations of faecal Prevotella and Bacteroides with specific dietary patterns. Microbiome 2016, 4, 57. [Google Scholar] [CrossRef]
- Tsigalou, C.; Paraschaki, A.; Karvelas, A.; Kantartzi, K.; Gagali, K.; Tsairidis, D.; Bezirtzoglou, E. Gut microbiome and Mediterranean diet in the context of obesity. Current knowledge, perspectives and potential therapeutic targets. Metab. Open 2021, 9, 100081. [Google Scholar] [CrossRef]
- Wisniewski, P.J.; Dowden, R.A.; Campbell, S.C. Role of Dietary Lipids in Modulating Inflammation through the Gut Microbiota. Nutrients 2019, 11, 117. [Google Scholar] [CrossRef]
- Masangcay, D.U.; Amado, A.J.Y.; Bulalas, A.R.; Ciudadano, S.R.; Fernandez, J.D.; Sastrillo, S.M.; Mabaggu, R.M. Association of Soil-transmitted Helminth Infection and Micronutrient Malnutrition: A Narrative Review. Asian J. Biol. Life Sci. 2021, 10, 317–324. [Google Scholar] [CrossRef]
- Levecke, B.; Montresor, A.; Albonico, M.; Ame, S.M.; Behnke, J.M.; Bethony, J.M.; Noumedem, C.D.; Engels, D.; Guillard, B.; Kotze, A.C.; et al. Assessment of Anthelmintic Efficacy of Mebendazole in School Children in Six Countries Where Soil-Transmitted Helminths Are Endemic. PLoS Neglected Trop. Dis. 2014, 8, e3204. [Google Scholar] [CrossRef]
- Ame, S.; Kabole, F.; Nanai, A.M.; Mwinzi, P.; Mupfasoni, D.; Ali, S.M.; Montresor, A. Impact of preventive chemotherapy on transmission of soil-transmitted helminth infections in Pemba Island, United Republic of Tanzania, 1994–2021. PLoS Neglected Trop. Dis. 2022, 16, e0010477. [Google Scholar] [CrossRef]
- Schneeberger, P.H.H.; Gueuning, M.; Welsche, S.; Hürlimann, E.; Dommann, J.; Häberli, C.; Frey, J.E.; Sayasone, S.; Keiser, J. Different gut microbial communities correlate with efficacy of albendazole-ivermectin against soil-transmitted helminthiases. Nat. Commun. 2022, 13, 1063. [Google Scholar] [CrossRef]
- Lawson, M.A.E.; Roberts, I.S.; Grencis, R.K. The interplay between Trichuris and the microbiota. Parasitology 2021, 148, 1806–1813. [Google Scholar] [CrossRef]
- Chen, H.; Mozzicafreddo, M.; Pierella, E.; Carletti, V.; Piersanti, A.; Ali, S.M.; Ame, S.M.; Wang, C.; Miceli, C. Dissection of the gut microbiota in mothers and children with chronic Trichuris trichiura infection in Pemba Island, Tanzania. Parasites Vectors 2021, 14, 62. [Google Scholar] [CrossRef]
- Goodwin, P.; Garrett, D.; Galal, O. Women and Family Health: The Role of Mothers in Promoting Family and Child Health. Int. J. Glob. Health Health Disparities 2005, 4, 30. Available online: https://scholarworks.uni.edu/ijghhdAvailableat:https://scholarworks.uni.edu/ijghhd/vol4/iss1/4 (accessed on 18 December 2023).
- Jian, C.; Carpén, N.; Helve, O.; de Vos, W.M.; Korpela, K.; Salonen, A. Early-life gut microbiota and its connection to metabolic health in children: Perspective on ecological drivers and need for quantitative approach. EBioMedicine 2021, 69, 103475. [Google Scholar] [CrossRef]
- Lukmanji, Z.; Hertzmark, E.; Mlingi, N.; Assey, V.; Ndossi, G.; Fawzi, W. Tanzania Food Composition Tables; MUHAS, TFNC, HSPH: Dar es Salaam, Tanzania, 2008. [Google Scholar]
- Nutrient Profile Model for the WHO African Region: A Tool for Implementing WHO Recommendations on the Marketing of Foods and Non-Alcoholic Beverages to Children; Licence: CC BY-NC-SA 3.0 IGO; WHO Regional Office for Africa: Brazzaville, Congo, 2018.
- EFSA (European Food Safety Authority). Dietary Reference Values for nutrients. Summary Report. EFSA Support. Publ. 2017, 98, e15121. [Google Scholar] [CrossRef]
- Górska-Warsewicz, H.; Rejman, K.; Laskowski, W.; Czeczotko, M. Milk and Dairy Products and Their Nutritional Contribution to the Average Polish Diet. Nutrients 2019, 11, 1771. [Google Scholar] [CrossRef]
- Muehlhoff, E.; Bennett, A.; Mcmahon, D. Dairy Products in Human Nutrition Dairy Products; Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2013. [Google Scholar]
- Uebanso, T.; Shimohata, T.; Mawatari, K.; Takahashi, A. Functional Roles of B-Vitamins in the Gut and Gut Microbiome. Mol. Nutr. Food Res. 2020, 64, 2000426. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Arzamasov, A.A.; Khoroshkin, M.S.; Iablokov, S.N.; Leyn, S.A.; Peterson, S.N.; Novichkov, P.S.; Osterman, A.L. Micronutrient Requirements and Sharing Capabilities of the Human Gut Microbiome. Front. Microbiol. 2019, 10, 1316. [Google Scholar] [CrossRef]
- Ministry of Health, Community Development, Gender, Elderly and Children (MoHCDGEC) [Tanzania Mainland]; Ministry of Health (MoH) [Zanzibar]; Tanzania Food and Nutrition Centre (TFNC); National Bureau of Statistics (NBS); Office of the Chief Government Statistician (OCGS) [Zanzibar]; UNICEF. Tanzania National Nutrition Survey Using SMART Methodology (TNNS); MoHCDGEC, MoH, TFNC, NBS, OCGS, and UNICEF: Dar es Salaam, Tanzania, 2018.
- Rusu, I.G.; Suharoschi, R.; Vodnar, D.C.; Pop, C.R.; Socaci, S.A.; Vulturar, R.; Istrati, M.; Moroșan, I.; Fărcaș, A.C.; Kerezsi, A.D.; et al. Iron Supplementation Influence on the Gut Microbiota and Probiotic Intake Effect in Iron Deficiency—A Literature-Based Review. Nutrients 2020, 12, 1993. [Google Scholar] [CrossRef]
- Dostal, A.; Chassard, C.; Hilty, F.M.; Zimmermann, M.B.; Jaeggi, T.; Rossi, S.; Lacroix, C. Iron Depletion and Repletion with Ferrous Sulfate or Electrolytic Iron Modifies the Composition and Metabolic Activity of the Gut Microbiota in Rats3. J. Nutr. 2012, 142, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium supplementation modulates gut microbiota in a prebiotic manner in dietary obese mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Whisner, C.M.; Martin, B.R.; Schoterman, M.H.C.; Nakatsu, C.H.; McCabe, L.D.; McCabe, G.P.; Wastney, M.E.; Heuvel, E.G.H.M.V.D.; Weaver, C.M. Galacto-oligosaccharides increase calcium absorption and gut bifidobacteria in young girls: A double-blind cross-over trial. Br. J. Nutr. 2013, 110, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Mehta, O.; Inbaraj, L.R.; Astbury, S.; Grove, J.I.; Norman, G.; Aithal, G.P.; Valdes, A.M.; Vijay, A. Gut Microbial Profile Is Associated With Residential Settings and Not Nutritional Status in Adults in Karnataka, India. Front. Nutr. 2021, 8, 595756. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Hayes, R.B. Implications for Noncommunicable Disease. Annu. Rev. Public Health 2021, 47, 277–292. [Google Scholar] [CrossRef]
- Martinez, K.B.; Leone, V.; Chang, E.B. Western diets, gut dysbiosis, and metabolic diseases: Are they linked? Gut Microbes 2017, 8, 130–142. [Google Scholar] [CrossRef]
- Elsherbiny, N.M.; Ramadan, M.; Abu Faddan, N.H.; Hassan, E.A.; E Ali, M.; El-Rehim, A.S.E.-D.A.; A Abbas, W.; Abozaid, M.A.; Hassanin, E.; A Mohamed, G.; et al. Impact of Geographical Location on the Gut Microbiota Profile in Egyptian Children with Type 1 Diabetes Mellitus: A Pilot Study. Int. J. Gen. Med. 2022, 15, 6173–6187. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2011, 5, 220–230. [Google Scholar] [CrossRef]
- De Angelis, M.; Ferrocino, I.; Calabrese, F.M.; De Filippis, F.; Cavallo, N.; Siragusa, S.; Rampelli, S.; Di Cagno, R.; Rantsiou, K.; Vannini, L.; et al. Diet influences the functions of the human intestinal microbiome. Sci. Rep. 2020, 10, 4247. [Google Scholar] [CrossRef]
- Rosa, B.A.; Snowden, C.; Martin, J.; Fischer, K.; Kupritz, J.; Beshah, E.; Supali, T.; Gankpala, L.; Fischer, P.U.; Urban, J.F.; et al. Whipworm-Associated Intestinal Microbiome Members Consistent Across Both Human and Mouse Hosts. Front. Cell. Infect. Microbiol. 2021, 11, 637570. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.; Walker, A.W.; Reyes, J.; Chico, M.; Salter, S.J.; Vaca, M.; Parkhill, J. Patent Human Infections with the Whipworm, Trichuris trichiura, Are Not Associated with Alterations in the Faecal Microbiota. PLoS ONE 2013, 8, e76573. [Google Scholar] [CrossRef] [PubMed]
- Skrypnik, K.; Suliburska, J. Association between the gut microbiota and mineral metabolism. J. Sci. Food Agric. 2018, 98, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Duncan, S.H.; Stams, A.J.; Van Dijl, J.M.; Flint, H.J.; Harmsen, H.J.M. The gut anaerobe Faecalibacterium prausnitzii uses an extracellular electron shuttle to grow at oxic–anoxic interphases. ISME J. 2012, 6, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- White, E.C.; Houlden, A.; Bancroft, A.J.; Hayes, K.S.; Goldrick, M.; Grencis, R.K.; Roberts, I.S. Manipulation of host and parasite microbiotas: Survival strategies during chronic nematode infection. Sci. Adv. 2018, 4, eaap7399. [Google Scholar] [CrossRef]
- Llinás-Caballero, K.; Caraballo, L. Helminths and Bacterial Microbiota: The Interactions of Two of Humans’ “Old Friends”. Int. J. Mol. Sci. 2022, 23, 13358. [Google Scholar] [CrossRef]
- Rosa, B.A.; Supali, T.; Gankpala, L.; Djuardi, Y.; Sartono, E.; Zhou, Y.; Fischer, K.; Martin, J.; Tyagi, R.; Bolay, F.K.; et al. Differential human gut microbiome assemblages during soil-transmitted helminth infections in Indonesia and Liberia. Microbiome 2018, 6, 33. [Google Scholar] [CrossRef]
- Williams, A.R.; Myhill, L.J.; Stolzenbach, S.; Nejsum, P.; Mejer, H.; Nielsen, D.S.; Thamsborg, S.M. Emerging interactions between diet, gastrointestinal helminth infection, and the gut microbiota in livestock. BMC Vet. Res. 2021, 17, 33. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
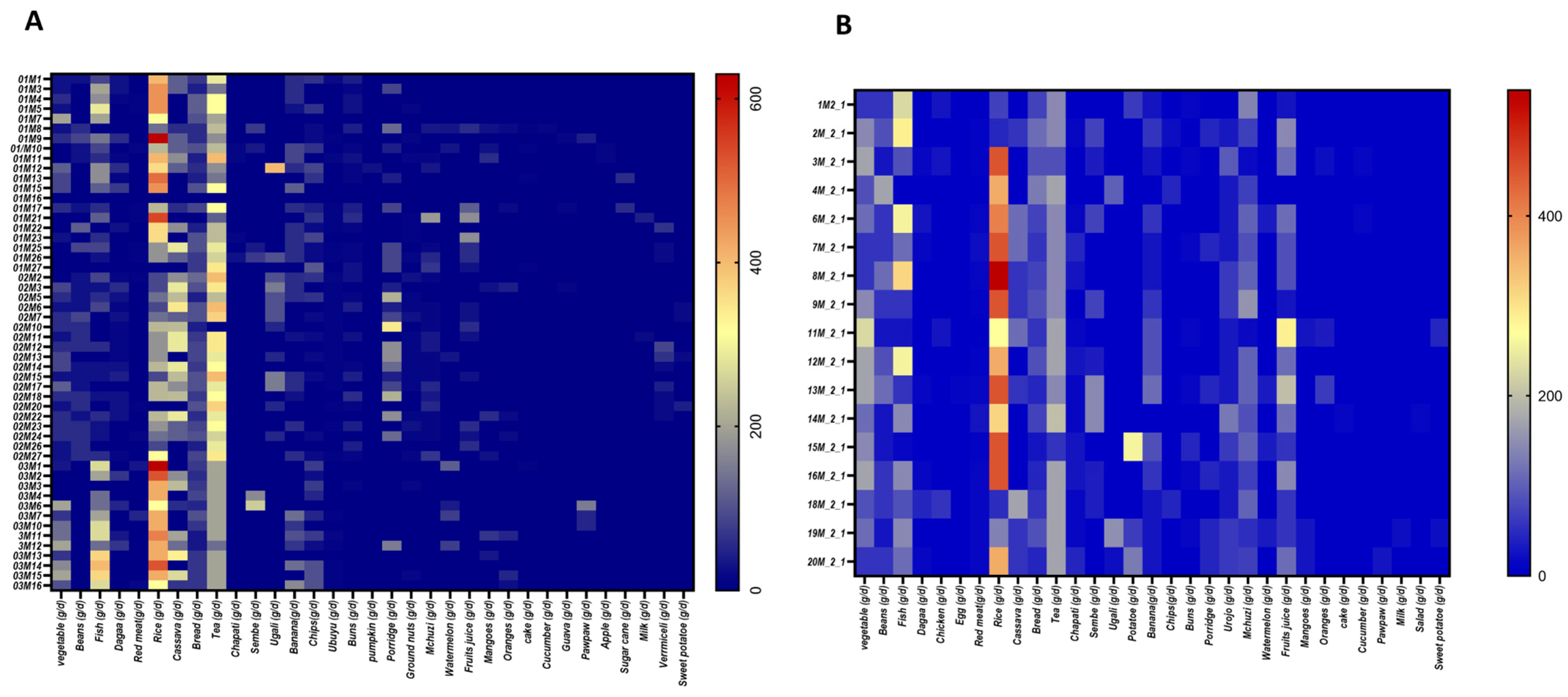

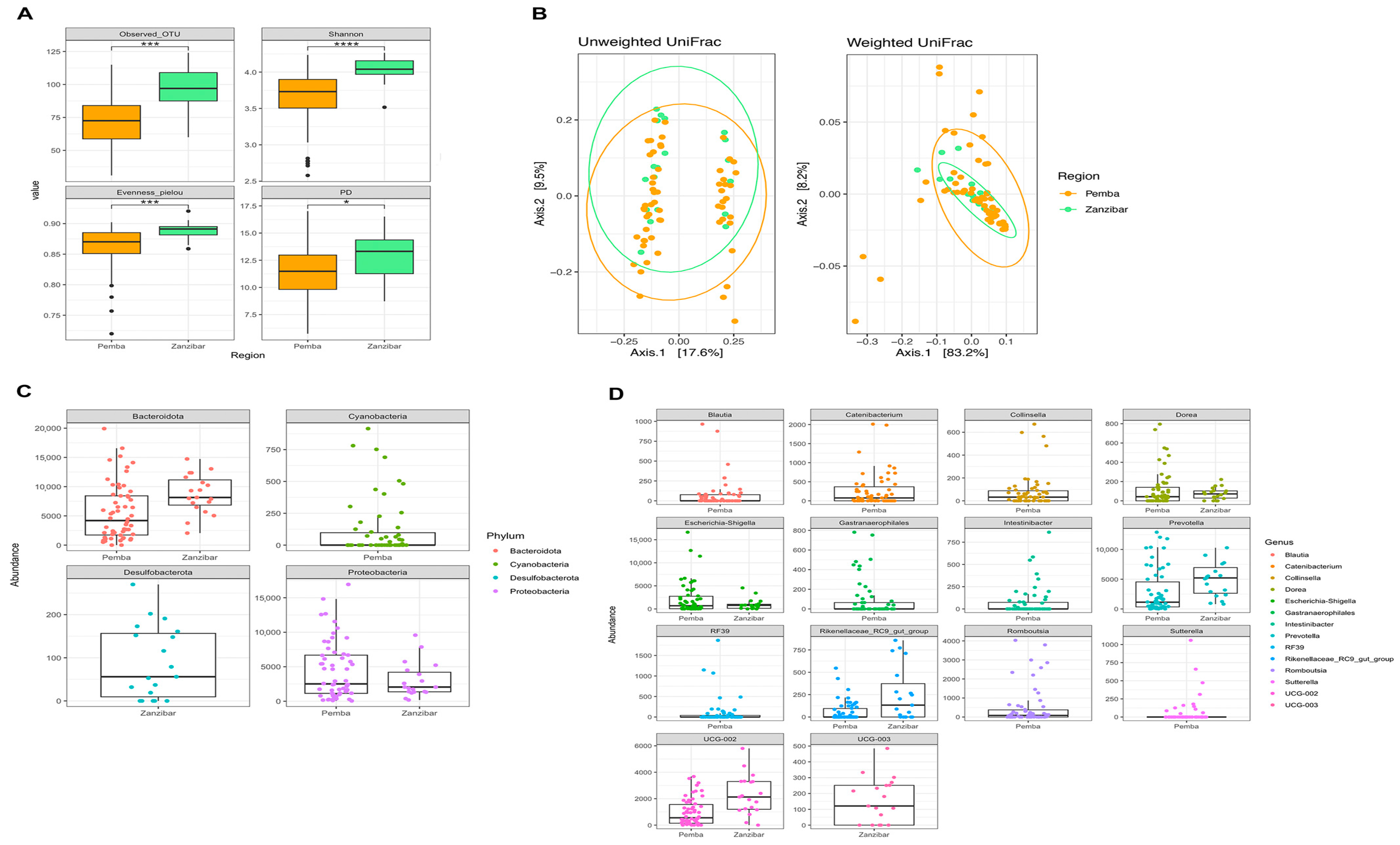

| Pemba (Mean ± Std) | Unguja (Mean ± Std) | |
|---|---|---|
| Age (years) | 30.04 ± 6.31 | 29.79 ± 7.36 |
| Weight (kg) | 59.29 ± 13.30 | 59.8 ± 11.19 |
| Height (cm) | 155.89 ± 5.95 | 138.52 ± 16.46 |
| BMI (kg/m2) | 24.34 ± 4.88 | 32.22 ± 9.13 |
| Nutrient | Intake (Pemba) | Intake (Unguja) | RNI | % of Women Reaching the RNI (Pemba) | % of Women Reaching the RNI (Unguja) |
|---|---|---|---|---|---|
| Protein (g) | 67.59 ± 21.19 | 76.87 ± 19.46 | 50–75 | 84.61 | 88.24 |
| Fat (g) | 72.59 ± 26.95 | 71.16 ± 18.68 | 33.33–66.66 | 96.15 | 88.24 |
| Carbohydrates (g) | 290.84 ± 52.85 | 274 ± 60.23 | 275–375 | 75 | 58.82 |
| Fibre (g) | 18.94 ± 7.82 | 21.60 ± 5.29 | 25 | 37.20 | 17 |
| Vitamin A (μg) | 499.89 ± 467.43 | 1124.12 ± 449.41 | 650 | 28.30 | 76.47 |
| Vitamin E (mg) | 4.4 ± 1.77 | 6.91 ± 4.40 | 11 | 0.0 | 11.76 |
| Vitamin C (mg) | 85.23 ± 44.28 | 91.54 ± 41.30 | 95 | 43.39 | 52.94 |
| Vitamin B1 (Thiamine) (mg) | 0.91 ± 0.23 | 1.08 ± 0.21 | 0.83 | 69.81 | 94.11 |
| Vitamin B2 (riboflavin) (mg) | 0.78 ± 0.22 | 1.04 ± 0.23 | 1.6 | 0.0 | 0.0 |
| Vitamin B3 (Niacin) (μg) | 8.34 ± 3.13 | 10.17 ± 2.69 | 14 | 7.54 | 11.76 |
| Vitamin B6 (pyridoxine) (mg) | 1.18 ± 0.36 | 1.55 ± 0.47 | 1.6 | 11.32 | 29.41 |
| Folate (μg) | 264.79 ± 109.07 | 428.21 ± 95.90 | 330 | 37.73 | 88.23 |
| Vitamin B12 (Cobalamin) (μg) | 2.76 ± 2.53 | 2.55 ± 1.61 | 2.4 | 41.50 | 41.17 |
| Vitamin B5 (pantothenic acid) (mg) | 2.51 ± 0.83 | 4.25 ± 2.75 | 5 | 0.0 | 17.64 |
| Calcium (mg) | 512.34 ± 360.05 | 599.45± 219.83 | 950 | 13.20 | 11.78 |
| Magnesium (mg) | 323.51 ± 112.05 | 367.85 ± 68.07 | 300 | 49.05 | 94.11 |
| Iron (mg) | 10.81 ± 5.01 | 12.25 ± 2.57 | 16 | 13.20 | 5.88 |
| Phosphorus (mg) | 1258 ± 451.03 | 1253.49 ± 336.45 | 550 | 94 | 100 |
| Zinc (mg) | 7.12 ± 2.26 | 7.64 ± 1.39 | 7.5 | 41.50 | 47.05 |
| Copper (mg) | 1.46 ± 0.60 | 1.49 ± 0.39 | 1.3 | 50.94 | 70.64 |
| Potassium (mg) | 2574.25 ± 825 | 2912.70 ± 545.41 | 3500 | 15 | 17.64 |
| Manganese (mg) | 5.15 ± 2.12 | 4.58 ± 1.11 | 3.0 | 88.67 | 94.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguélé, A.T.; Carrara, C.; Mozzicafreddo, M.; Chen, H.; Piersanti, A.; Salum, S.S.; Ali, S.M.; Miceli, C. Association between Food or Nutrients and Gut Microbiota in Healthy and Helminth-Infected Women of Reproductive Age from Zanzibar, Tanzania. Nutrients 2024, 16, 1266. https://doi.org/10.3390/nu16091266
Nguélé AT, Carrara C, Mozzicafreddo M, Chen H, Piersanti A, Salum SS, Ali SM, Miceli C. Association between Food or Nutrients and Gut Microbiota in Healthy and Helminth-Infected Women of Reproductive Age from Zanzibar, Tanzania. Nutrients. 2024; 16(9):1266. https://doi.org/10.3390/nu16091266
Chicago/Turabian StyleNguélé, Aristide Toussaint, Chiara Carrara, Matteo Mozzicafreddo, Hongliang Chen, Angela Piersanti, Salum Seif Salum, Said M. Ali, and Cristina Miceli. 2024. "Association between Food or Nutrients and Gut Microbiota in Healthy and Helminth-Infected Women of Reproductive Age from Zanzibar, Tanzania" Nutrients 16, no. 9: 1266. https://doi.org/10.3390/nu16091266