IGF-1, the Cross Road of the Nutritional, Inflammatory and Hormonal Pathways to Frailty


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Insulin-Like Growth Factor
2.1. Insulin-Like Growth Factor Anabolic Hormone
2.2. Insulin-Like Growth Factor and Inflammation
2.3. Insulin-Like Growth Factor: A Sensitive Nutritional Marker in the Elderly
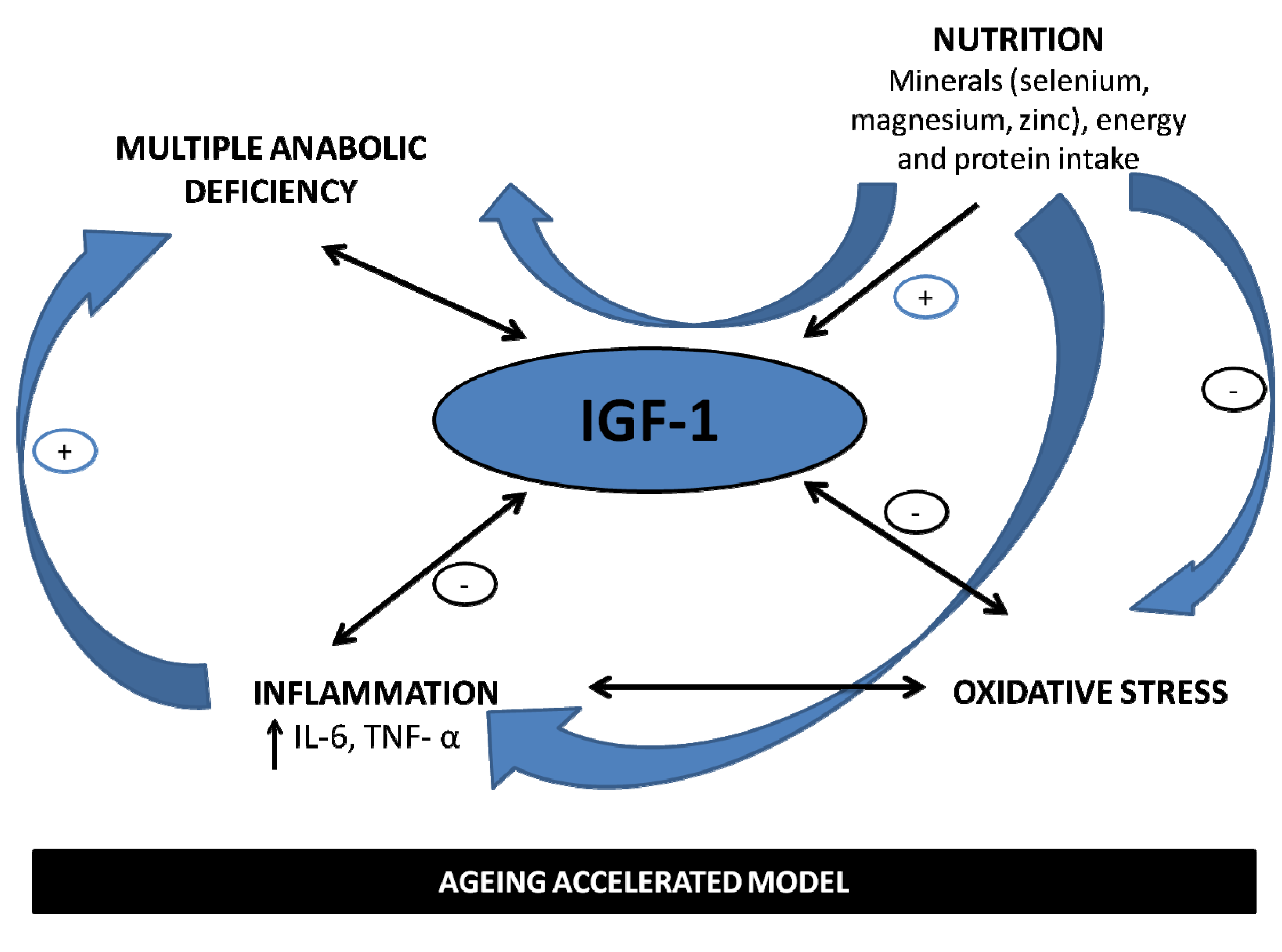
3. Minerals and IGF-1 in the Fire of Frailty

3.1. Magnesium and IGF-1
3.2. Selenium and IGF-1
3.3. Zinc and IGF-1
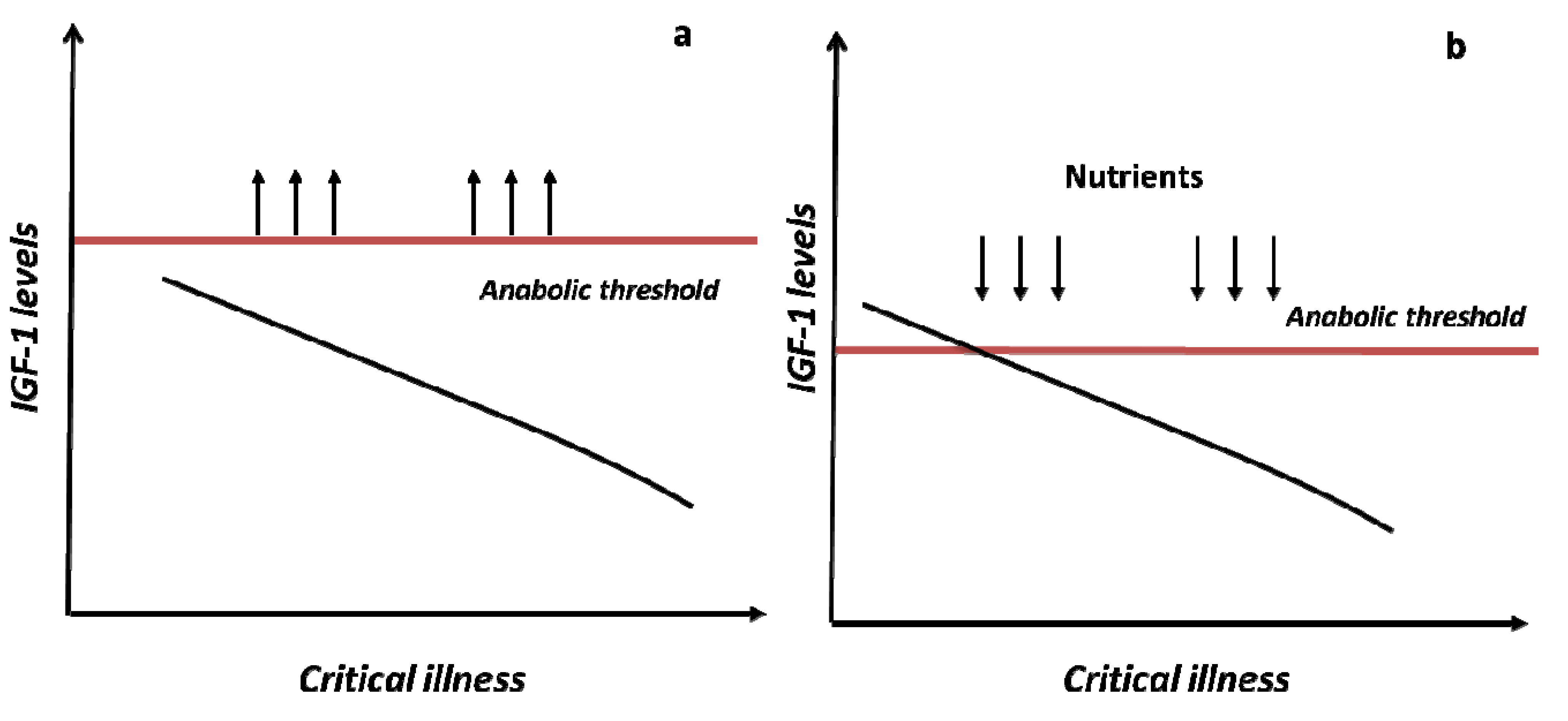
4. Perspectives

5. Conclusions
Conflicts of Interest
References
- Delmonico, M.J.; Harris, T.B.; Lee, J.S.; Visser, M.; Nevitt, M.; Kritchevsky, S.B.; Tylavsky, F.A.; Newman, A.B. Health, aging and body composition study. Alternative definitions of sarcopenia, lower extremity performance, and functional impairment with aging in older men and women. J. Am. Geriatr. Soc. 2007, 55, 769–774. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; Simonsick, E.M.; Tylavsky, F.A.; Visser, M.; Newman, A.B. The loss of skeletal muscle strength, mass, and quality in older adults: The health, aging and body composition study. J. Gerontol. 2006, 61, 1059–1064. [Google Scholar] [CrossRef]
- Fried, L.P.; Tangen, C.M.; Walston, J.; Newman, A.B.; Hirsch, C.; Gottdiener, J.; Seeman, T.; Tracy, R.; Kop, W.J.; Burke, G.; et al. Frailty in older adults: Evidence for a phenotype. J. Gerontol. 2001, 56, M146–M156. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauuer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 1, 412–423. [Google Scholar]
- Rosenberg, I.H. Sarcopenia: Origins and clinical relevance. J. Nutr. 1997, 127, 990S–991S. [Google Scholar]
- Maggio, M.; Lauretani, F.; Ceda, G.P.; Bandinelli, S.; Ling, S.M.; Metter, E.J.; Artoni, A.; Carassale, L.; Cazzato, A.; Ceresini, G.; et al. Relationship between low levels of anabolic hormones and 6-year mortality in older men: The aging in the Chianti Area (InCHIANTI) study. Arch. Intern. Med. 2007, 167, 2249–2254. [Google Scholar] [CrossRef]
- Maggio, M.; Dall’Aglio, E.; Lauretani, F.; Cattabiani, C.; Ceresini, G.; Caffarra, P.; Valenti, G.; Volpi, R.; Vignali, A; Schiavi, G.; et al. The hormonal pathway to cognitive impairment in older men. J. Nutr. Health Aging 2012, 16, 40–54. [Google Scholar] [CrossRef]
- Arvat, E.; Broglio, F.; Ghigo, E. Insulin-like growth factor I: Implications in aging. Drugs Aging 2000, 16, 29–40. [Google Scholar] [CrossRef]
- Rosen, C.J. Serum insulin-like grow factor and insulin like grow factor-binding proteins: Clinical implication. Clin. Chem. 1999, 45, 1384–1390. [Google Scholar]
- Estívariz, C.F.; Ziegler, T.R. Nutrition and the insulin-like growth factor system. Endocrine 1997, 7, 65–71. [Google Scholar] [CrossRef]
- Sherlock, M.; Toogood, A.A. Aging and the growth hormone/insulin like growth factor-I axis. Pituitary 2007, 10, 189–203. [Google Scholar] [CrossRef]
- Lee, P.D.; Giudice, L.C.; Conover, C.A.; Powell, D.R. Insulin-like growth factor binding protein 1: Recent findings and new directions. Proc. Soc. Exp. Biol. Med. 1997, 216, 319–357. [Google Scholar] [CrossRef]
- Franklin, S.L.; Ferry, R.J.; Cohen, P. Rapid insulin-like growth factor (IGF)-independent effects of IGF binding protein-3 on endothelial cell survival. J. Clin. Endocrinol. Metab. 2003, 88, 900–907. [Google Scholar] [CrossRef]
- Hoffman, A.R.; Pyka, G.; Liecerman, S.A.; Ceda, G.P.; Marcus, R. The Somatopause. In Growth Hormone and Somatomedins during Lifespan; Muller, E.E., Cocchi, D., Locatelli, V., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 265–274. [Google Scholar]
- De Marinis, L.; Mancini, A.; Giampietro, A.; Gentilella, R.; Bianchi, A.; Perrelli, M.; Vezzosi, C.; Milardi, D.; Fusco, A.; Valle, D.; et al. GH deficiency syndrome in elderly patients. J. Endocrinol.Investig. 2002, 25, 40–41. [Google Scholar]
- Cappola, A.R.; Bandeen-Roche, K.; Wand, G.S.; Volpato, S.; Fried, L.P. Association of IGF-I levels with muscle strength and mobility in older women. J. Clin. Endocrinol. Metab. 2001, 86, 4139–4146. [Google Scholar] [CrossRef]
- Colbert, L.H.; Rosen, C.J.; Goodpaster, B.H.; Newman, A.B.; Kritchevsky, S.B.; Satterfield, S.; Kanaya, A.M.; Taaffe, D.R.; Harris, T.B. Insulin-like growth factor-1. J. Am. Geriatr. Soc. 2004, 52, 1962–1963. [Google Scholar] [CrossRef]
- Ferrucci, L.; Corsi, A.; Lauretani, F.; Bandinelli, S.; Bartali, B.; Taub, D.D.; Guralnik, J.M.; Longo, D.L. The origins of age-related proinflammatory state. Blood 2005, 105, 2294–2299. [Google Scholar] [CrossRef]
- Elosua, R.; Bartali, B.; Ordovas, J.M.; Corsi, A.M.; Lauretani, F.; Ferrucci, L. Association between physical activity, physical performance, and inflammatory biomarkers in an elderly population: The InCHIANTI study. J. Gerontol. 2005, 60, 760–767. [Google Scholar] [CrossRef]
- Cesari, M.; Penninx, B.W.; Pahor, M.; Lauretani, F.; Corsi, A.M.; Rhys Williams, G.; Guralnik, J.M.; Ferrucci, L. Inflammatory markers and physical performance in older persons: The InCHIANTI study. J. Gerontol. 2004, 59, 242–248. [Google Scholar] [CrossRef]
- Ferrucci, L.; Harris, T.; Guralnik, J.; Tracy, R.P.; Corti, M.C.; Cohen, H.J.; Cohen, B.; Penninx, B.; Pahor, M.; Wallace, R.; et al. Serum IL-6 level and the development of disability in older persons. J. Am. Geriatr. Soc. 1999, 47, 662–668. [Google Scholar]
- Lazarus, D.D.; Moldawer, L.L.; Lowry, S.F. Insulin like growth factor-1 activity is inhibited by interleukine alpha, tumor necrosis factor-alpha, and interleukin-6. Lymphokine Cytokine Res. 1993, 12, 219–223. [Google Scholar]
- Barbieri, M.; Ferrucci, L.; Ragno, E.; Corsi, A.; Bandinelli, S.; Bonafè, M.; Olivieri, F.; Giovagnetti, S.; Franceschi, C.; Guralnik, J.M.; et al. Chronic inflammation and the effect of IGF-I on muscle strength and power in older persons. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E481–E487. [Google Scholar]
- Stenholm, S.; Maggio, M.; Lauretani, F.; Bandinelli, S.; Ceda, G.P.; Di Iorio, A.; Giallauria, F.; Guralnik, J.M.; Ferrucci, L. Anabolic and catabolic biomarkers as predictors of muscle strength decline: The InCHIANTI study. Rejuvenation Res. 2010, 13, 3–11. [Google Scholar] [CrossRef]
- Cappola, A.R.; Xue, Q.L.; Ferrucci, L.; Guralnik, J.M.; Volpato, S.; Fried, L.P. Insulin-like growth factor I and interleukin-6 contribute synergistically to disability and mortality in older women. J. Clin. Endocrinol. Metab. 2003, 88, 2019–2025. [Google Scholar] [CrossRef]
- Roubenoff, R.; Parise, H.; Payette, H.A.; Abad, L.W.; D’Agostino, R.; Jacques, P.F.; Wilson, P.W.; Dinarello, C.A.; Harris, T.B. Cytokines, insulin-like growth factor 1, sarcopenia, and mortality in very old community-dwelling men and women: The Framingham Heart Study. Am. J. Med. 2003, 115, 429–435. [Google Scholar] [CrossRef]
- Fernandez, S.; Fernandez, A.M.; Lopez-Lopez, C.; Torres-Aleman, I. Emerging roles of insulin-like growth factor 1 in the adult brain. Growth Horm. IGF Res. 2007, 17, 89–95. [Google Scholar] [CrossRef]
- Yanase, S.; Ishii, N. Hyperoxia exposure induced hormesis decreases mithocondrial superoxide radical levels via Ins/IGF-1 signaling pathway in a long-lived age-1 mutant on Caenorhabditis elegans. J. Radiat. Res. 2008, 49, 211–218. [Google Scholar] [CrossRef]
- Underwood, L.E.; Clemmons, D.R.; Maes, M.; D’Ercole, A.J.; Ketelslegers, J.M. Regulation of somatomedin-C/insulin-like growth factor I by nutrients. Horm. Res. 1986, 24, 166–176. [Google Scholar] [CrossRef]
- Norat, T.; Dossus, L.; Rinaldi, S.; Overvad, K.; Grønbaek, H.; Tjønneland, A.; Olsen, A.; Clavel-Chapelon, F.; Boutron-Ruault, M.C.; Boeing, H.; et al. Diet, serum insulin-like growth factor-I and IGF-binding protein-3 in European women. Eur. J. Clin. Nutr. 2007, 61, 91–98. [Google Scholar] [CrossRef]
- Thissen, J.P.; Ketelslegers, J.M.; Underwood, L.E. Nutritional regulation of the insulin-like growth factors. Endocr. Rev. 1994, 15, 80–101. [Google Scholar]
- Holmes, M.D.; Pollack, M.N.; Willett, W.C.; Hankinson, S.E. Dietary correlates of plasma insulin-like growth factor 1 and insulin-like growth factor binding protein 3 concentrations. Cancer Epidemiol. Biomark. Prev. 2002, 11, 852–861. [Google Scholar]
- Gran, I.T.; Norat, T.; Rinaldi, S.; Dossus, L.; Ulanova, A.; Téhard, B.; Clavel-Chapelon, F.; van Gils, C.H.; van Noord, P.A.; Peeters, P.H.; et al. Body mass index, waist circumference and waist-hip ratio and serum levels of IGF-I and IGFBP-3 in European women. Int. J. Obes. 2006, 30, 1623–1631. [Google Scholar] [CrossRef]
- Salbe, A.D.; Kotler, D.P.; Wang, J.; Pierson, R.N.; Campbell, R.G. Predictive value of IGF-1 concentrations in HIV-infected patients. Clin. Res. 1991, 39, 385A. [Google Scholar]
- Kirschner, B.S.; Sutton, M.M. Somatomedin-C levels in growth-impaired children and adolescents with chronic inflammatory bowel disease. Gastroenterology 1986, 91, 830–836. [Google Scholar]
- Clemmons, D.R.; Klibanski, A.; Underwood, L.E.; McArthur, J.W.; Ridgway, E.C.; Beitins, I.Z.; van Wyk, J.J. Reduction of plasma immunoreactive somatomedin-C during fasting in humans. J. Clin. Endorinol. Metab. 1981, 53, 1247–1250. [Google Scholar] [CrossRef]
- Tirapegui, J.; Fukushima, S.E.; Grimaldi, G. Growth, somatomedin and nutrition. Arch. Latinoam. Nutr. 1993, 43, 94–104. [Google Scholar]
- Vehe, K.L.; Brown, R.O.; Moore, L.W.; Acchiardo, S.R.; Luther, R.W. The efficacy of nutrition support in infected patients with chronic renal failure. Pharmacotherapy 1991, 11, 303–307. [Google Scholar]
- Lecornu, M.; David, L.; Francois, R. Low serum somatomedin activity in celiac disease. Helv. Paediatr. Acta 1978, 33, 509–516. [Google Scholar]
- O’Connor, J.C.; McCusker, R.H.; Strle, K.; Johnson, R.W.; Dantzer, R.; Kelley, K.W. Regulation of IGF-I function by proinflammatory cytokines: At the interface of immunology and endocrinology. Cell. Immunol. 2008, 252, 91–110. [Google Scholar] [CrossRef]
- Ni, F.; Sun, R.; Fu, B.; Wang, F.; Guo, C.; Tian, Z.; Wei, H. IGF-1 promotes the development and cytotoxic activity of human NK cells. Nat. Commun 2013, 4. [Google Scholar] [CrossRef]
- Brill, K.T.; Weltman, A.L.; Gentili, A.; Patrie, J.T.; Fryburg, D.A.; Hanks, J.B.; Urban, R.J.; Veldhuis, J.D. Single and combined effects of growth hormone and testosterone administration on measures of body composition, physical performance, mood, sexual function, bone turnover, and muscle gene expression in healthy older men. Clin. Endocrinol. Metab. 2002, 87, 5649–5657. [Google Scholar] [CrossRef]
- Wolf, F.I.; Cittadini, A. Chemistry and biochemistry of magnesium. Mol. Asp. Med. 2003, 24, 3–9. [Google Scholar] [CrossRef]
- Barbagallo, M.; Dominguez, L.J.; Galioto, A.; Ferlisi, A.; Cani, C.; Malfa, L.; Pineo, A.; Busardo’, A.; Paolisso, G. Role of magnesium in insulin action, diabetes and cardiometabolic syndrome X. Mol. Asp. Med. 2003, 24, 39–52. [Google Scholar] [CrossRef]
- Champagne, C.M. Magnesium in hypertension, cardiovascular disease, metabolic syndrome, and other conditions: A review. Nutr. Clin. Pract. 2008, 23, 142–151. [Google Scholar] [CrossRef]
- Lukaski, H.C.; Nielsen, F.H. Dietary magnesium depletion affects metabolic responses during submaximal exercise in postmenopausal women. J. Nutr. 2003, 132, 930–935. [Google Scholar]
- Brilla, L.R.; Haley, T.F. Effect of magnesium supplementation on strength training in humans. J. Am. Coll. Nutr. 1992, 11, 326–329. [Google Scholar] [CrossRef]
- Vaquero, M.P. Magnesium and trace elements in the elderly: Intake, status and recommendations. J. Nutr. Health Aging 2002, 6, 147–153. [Google Scholar]
- Dominguez, L.J.; Barbagallo, M.; Lauretani, F.; Bandinelli, S.; Bos, A.; Corsi, A.M.; Simonsick, E.M.; Ferrucci, L. Magnesium and muscle performance in older persons: The InCHIANTI study. Am. J. Clin. Nutr. 2006, 84, 419–426. [Google Scholar]
- Maggio, M.; Ceda, G.P.; Lauretani, F.; Cattabiani, C.; Avantaggiato, E.; Morganti, S.; Ablondi, F.; Bandinelli, S.; Dominguez, L.J.; Barbagallo, M.; et al. Magnesiumand anabolic hormones in older men. Int. J. Androl. 2011, 34, e594–e600. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Barbagallo, M.; Sowers, J.R.; Resnick, L.M. Magnesium responsiveness to insulin and insulin-like growth factor I in erythrocytes from normotensive and hypertensive subjects. J. Clin. Endocrinol. Metab. 1998, 83, 4402–4407. [Google Scholar] [CrossRef]
- Rayssiguier, Y.; Durlach, J.; Gueux, E.; Rock, E.; Mazur, A. Magnesium and aging. I. Experimental data: Importance of oxidative damage. Magnes. Res. 1993, 6, 369–378. [Google Scholar]
- Yang, Y.; Wu, Z.; Chen, Y.; Qiao, J.; Gao, M.; Yuan, J.; Nie, W.; Guo, Y. Magnesium deficiency enhances hydrogen peroxide production and oxidative damage in chick embryo hepatocyte in vitro. Biometals 2006, 19, 71–81. [Google Scholar] [CrossRef]
- Barbagallo, M.; Dominguez, L.J. Magnesium and aging. Curr. Pharm. Des. 2010, 16, 832–839. [Google Scholar] [CrossRef]
- Hunt, C.D.; Johnsom, L.K. Magnesium requirements: New estimations for men and women by cross-sectional statistical analyses of metabolic magnesium balance data. Am. J. Clin. Nutr. 2006, 84, 843–852. [Google Scholar]
- Ford, E.S.; Mokdad, A.H. Dietary magnesium intake in a national sample of US adults. J. Nutr. 2003, 133, 2879–2882. [Google Scholar]
- Barbagallo, M.; Belvedere, M.; Dominguez, L.J. Magnesium homeostasis and aging. Magnes. Res. 2009, 22, 235–246. [Google Scholar]
- Gullestad, L.; Nes, M.; Rønneberg, R.; Midtvedt, K.; Falch, D.; Kjekshus, J. Magnesium status in healthy free-living elderly Norwegians. J. Am. Coll. Nutr. 1994, 13, 45–50. [Google Scholar] [CrossRef]
- Lameris, A.L.; Monnens, L.A.; Bindels, R.J.; Hoenderop, J.G. Drug-induced alterations in Mg2+ homoeostasis. Clin. Sci. 2012, 123, 1–14. [Google Scholar] [CrossRef]
- Cundy, T.; Dissanayake, A. Severe hypomagnesaemia in long-term users of proton-pump inhibitors. Clin. Endocrinol. 2008, 69, 338–341. [Google Scholar] [CrossRef]
- Hoorn, E.J.; van der Hoek, J.; de Man, R.A.; Kuipers, E.J.; Bolwerk, C.; Zietse, R. A case series of proton pump inhibitor-induced hypomagnesemia. Am. J. Kidney Dis. 2010, 56, 112–116. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell. Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef]
- Markropoulos, V.; Bruning, T.; Schulze-Osthoff, K. Selenium-mediated inhibition of transcription factor NK-kappa B and HIV-1 LTR promoter activity. Arch. Toxicol. 1996, 70, 277–283. [Google Scholar] [CrossRef]
- Kaeck, M.; Lu, J.; Strange, R.; Ganther, H.E.; Thompson, H.J. Differential induction of growth arrest inducible genes by selenium compounds. Biochem. Pharmacol. 1997, 53, 921–926. [Google Scholar] [CrossRef]
- Schomburg, L.; Köhrle, J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E. Selenoprotein P-expression, functions, and roles in mammals. Biochim. Biophys. Acta 2009, 1790, 1441–1447. [Google Scholar] [CrossRef]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar]
- World Health Organization (WHO), International Programme on Chemical Safety; WHO: Geneva, Switzerland, 1987.
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Bleys, J.; Navas-Acien, A.; Guallar, E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch. Intern. Med. 2008, 168, 404–410. [Google Scholar] [CrossRef]
- Akbaraly, N.T.; Arnaud, J.; Hininger-Favier, I.; Gourlet, V.; Roussel, A.M.; Berr, C. Selenium and mortality in the elderly: Results from the EVA study. Clin. Chem. 2005, 51, 2117–2123. [Google Scholar] [CrossRef]
- Lauretani, F.; Semba, R.D.; Bandinelli, S.; Ray, A.L.; Ruggiero, C.; Cherubini, A.; Guralnik, J.M.; Ferrucci, L. Low plasma selenium concentrations and mortality among older community-dwelling adults: The InCHIANTI study. Aging Clin. Exp. Res. 2008, 20, 153–158. [Google Scholar] [CrossRef]
- Hurwitz, B.E.; Klaus, J.R.; Llabre, M.M.; Gonzalez, A.; Lawrence, P.J.; Maher, K.J.; Greeson, J.M.; Baum, M.K.; Shor-Posner, G.; Skyler, J.S.; et al. Suppression of human immunodeficiency virus type 1 viral load with selenium supplementation: A randomized controlled trial. Arch. Intern. Med. 2007, 167, 148–154. [Google Scholar] [CrossRef]
- Toulis, K.A.; Anastasilakis, A.D.; Tzellos, T.G.; Goulis, D.G.; Kouvelas, D. Selenium supplementation in the treatment of Hashimoto’s thyroiditis: A systematic review and a meta-analysis. Thyroid 2010, 20, 1163–1173. [Google Scholar] [CrossRef]
- Amaral, A.F.; Cantor, K.P.; Silverman, D.T.; Malats, N. Selenium and bladder cancer risk: A meta-analysis. Cancer Epidemiol. Biomark. Prev. 2010, 19, 2407–2415. [Google Scholar] [CrossRef]
- Etminan, M.; FitzGerald, J.M.; Gleave, M.; Chambers, K. Intake of selenium in the prevention of prostate cancer: A systematic review and meta-analysis. Cancer Causes Control 2005, 16, 1125–1131. [Google Scholar] [CrossRef]
- Rederstorff, M.; Krol, A.; Lescure, A. Understanding the importance of selenium and selenoproteins in muscle function. Cell. Mol. Life Sci. 2006, 63, 52–59. [Google Scholar] [CrossRef]
- Moosmann, B.; Behl, C. Selenoprotein synthesis and side-effects of statins. Lancet 2004, 363, 892–894. [Google Scholar] [CrossRef]
- Beck, J.; Ferrucci, L.; Sun, K.; Walston, J.; Fried, L.P.; Varadhan, R.; Guralnik, J.; Semba, R.D. Low serum selenium concentrations are associated with poor grip strength among older women living in the community. Biofactors 2007, 29, 37–44. [Google Scholar] [CrossRef]
- Lauretani, F.; Semba, R.D.; Bandinelli, S.; Ray, A.L.; Guralnik, J.M.; Ferrucci, L. Association of low plasma selenium concentrations with poor muscle strength in older community-dwelling adults: The InCHIANTI study. Am. J. Clin. Nutr. 2007, 86, 347–352. [Google Scholar]
- Helmersson, J.; Arnlov, J.; Vessby, B.; Larsson, A.; Alfthan, G.; Basu, S. Serum selenium predicts levels of F2-isoprostanes and prostaglandin F2alpha in a 27 year follow-up study of Swedish men. Free Radic. Res. 2005, 39, 763–770. [Google Scholar] [CrossRef]
- Karl, J.P.; Alemany, J.A.; Koenig, C.; Kraemer, W.J.; Frystyk, J.; Flyvbjerg, A.; Young, A.J.; Nindl, B.C. Diet, body composition, and physical fitness influences on IGF-I bioactivity in women. Growth Horm. IGF Res. 2009, 19, 491–496. [Google Scholar] [CrossRef]
- Maggio, M.; Ceda, G.P.; Lauretani, F.; Bandinelli, S.; Dall’Aglio, E.; Guralnik, J.M.; Paolisso, G.; Semba, R.D.; Nouvenne, A.; Borghi, L.; et al. Association of plasma selenium concentrations with total IGF-1 among older community-dwelling adults: The InCHIANTI study. Clin. Nutr. 2010, 29, 674–677. [Google Scholar] [CrossRef]
- Moreno-Reyes, R.; Egrise, D.; Nève, J.; Pasteels, J.L.; Schoutens, A. Selenium deficiency-induced growth retardation is associated with an impaired bone metabolism and osteopenia. J. Bone Miner. Res. 2001, 16, 1556–1563. [Google Scholar] [CrossRef]
- Thompson, K.M.; Haibach, H.; Sunde, R.A. Growth and plasma triiodothyronine concentrations are modified by selenium deficiency and repletion in second-generation selenium-deficient rats. J. Nutr. 1995, 125, 864–873. [Google Scholar]
- Semba, R.D.; Ricks, M.O.; Ferrucci, L.; Xue, Q.L.; Guralnik, J.M.; Fried, L.P. Low serum selenium is associated with anemia among older adults in the United State. Eur.J. Clin. Nutr. 2009, 63, 93–99. [Google Scholar] [CrossRef]
- Vihervuori, E.; Virtanem, M.; Koistinen, H.; Koistinen, R.; Seppala, M.; Siimes, M.A. Haemoglobin level is linked to growth hormone-dependent proteins in short children. Blood 1996, 87, 2075–2081. [Google Scholar]
- Nilsson-Ehle, H.; Bengtsson, B.A.; Lindstedt, G.; Mellstrom, D. Insulin-like growth factor-1 is a predictor of blood haemoglobin concentration in 70-yr-old subjects. Eur. J. Haematol. 2005, 74, 111–116. [Google Scholar] [CrossRef]
- Walston, J.; Xue, Q.; Semba, R.D.; Ferrucci, L.; Cappola, A.R.; Ricks, M.; Guralnik, J.; Fried, L.P. Serum antioxidants, inflammation, and mortality in older women. Am. J. Epidemiol. 2006, 163, 18–26. [Google Scholar]
- Meunier, N.; O’Connor, J.M.; Maiani, G.; Cashman, K.D.; Secker, D.L.; Ferry, M.; Roussel, A.M.; Coudray, C. Importance of zinc in the elderly: The ZENITH study. Eur. J. Clin. Nutr. 2005, 59, S1–S4. [Google Scholar]
- Hambidge, M. Human zinc deficiency. J. Nutr. 2000, 130, 1344S–1349S. [Google Scholar]
- Matsui, T.; Yamaguchi, M. Zinc modulation of insulin-like growth factor’s effect in osteoblastic MC3T3–E1 cells. Peptides 1995, 16, 1063–1068. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Hashizume, M. Effect of beta-alanyl-l-histidinato zinc on protein components in osteoblastic MC3T3-El cells: Increase in osteocalcin, insulin-like growth factor-I and transforming growth factor-beta. Mol. Cell. Biochem. 1994, 136, 163–169. [Google Scholar] [CrossRef]
- McNall, A.D.; Etherton, T.D.; Fosmire, G.J. The impaired growth induced by zinc deficiency in rats is associated with decreased expression of the hepatic insulin-like growth factor I and growth hormone receptor genes. J. Nutr. 1995, 125, 874–879. [Google Scholar]
- Ninh, N.X.; Thissen, J.P.; Maiter, D.; Adam, E.; Mulumba, N.; Ketelslegers, J.M. Reduced liver insulin-like growth factor-I gene expression in young zinc deprived rats is associated with a decrease in liver growth hormone (GH) receptors and serum GH-binding protein. J. Endocrinol. 1995, 144, 449–456. [Google Scholar] [CrossRef]
- Droke, E.A.; Spears, J.W.; Armstrong, J.D.; Kegley, E.B.; Simpson, R.B. Dietary zinc affects serum concentrations of insulin and insulin-like growth factor I in growing lambs. J. Nutr. 1993, 123, 13–19. [Google Scholar]
- Roth, H.P.; Kirchgessner, M. Influence of alimentary zinc deficiency on the concentration of growth hormone (GH), insulin-like growth factor I (IGF-I) and insulin in the serum of force-fed rats. Horm. Metab. Res. 1994, 26, 404–408. [Google Scholar] [CrossRef]
- Dorup, I.; Flyvbjerg, A.; Everts, M.E.; Clausen, T. Role of insulin-like growth factor-1 and growth hormone in growth inhibition induced by magnesium and zinc deficiencies. Br. J. Nutr. 1991, 66, 505–552. [Google Scholar] [CrossRef]
- Browning, J.D.; MacDonald, R.S.; Thornton, W.H.; O’Dell, B.L. Reduced food intake in zinc deficient rats is normalized by megestrol acetate but not by insulin-like growth factor-I. J. Nutr. 1998, 128, 136–142. [Google Scholar]
- Cossack, Z.T. Decline in somatomedin-C, insulin-like growth factor-1, with experimentally induced zinc deficiency in human subjects. Clin. Nutr. 1991, 10, 284–291. [Google Scholar] [CrossRef]
- Nakamura, T.; Nishiyama, S.; Futagoishi-Suginohara, Y.; Matsuda, I.; Higashi, A. Mild to moderate zinc deficiency in short children: Effect of zinc supplementation on linear growth velocity. J. Pediatr. 1993, 123, 65–69. [Google Scholar] [CrossRef]
- Ninh, N.X.; Thissen, J.P.; Collette, L.; Gerard, G.; Khoi, H.H.; Ketelslegers, J.M. Zinc supplementation increases growth and circulating insulin-like growth factor I (IGF-I) in growth-retarded Vietnamese children. Am. J. Clin. Nutr. 1996, 63, 514–519. [Google Scholar]
- Hamza, R.T.; Hamed, A.I.; Sallam, M.T. Effect of zinc supplementation on growth hormone-insulin growth factor axis in short Egyptian children with zinc deficiency. Ital. J. Pediatr. 2012, 24, 38. [Google Scholar]
- Imamoğlu, S.; Bereket, A.; Turan, S.; Taga, Y.; Haklar, G. Effect of zinc supplementation on growth hormone secretion, IGF-I, IGFBP-3, somatomedin generation, alkaline phosphatase, osteocalcin and growth in prepubertal children with idiopathic short stature. J. Pediatr. Endocrinol. Metab. 2005, 18, 69–74. [Google Scholar]
- Blumberg, J. Nutritional needs of seniors. J. Am. Coll. Nutr. 1997, 16, 517–523. [Google Scholar]
- Herzberg, M.; Foldes, J.; Steinberg, R.; Menczel, J. Zinc excretion in osteoporotic women. J. Bone Miner. Res. 1990, 3, 251–257. [Google Scholar]
- Walhvork, J.C.; Sandstead, H.H. Zinc. In Nutrition and Bone Development; Simmons, D.J., Ed.; Oxford University Press: New York, NY, USA, 1990; p. 316. [Google Scholar]
- Gur, A.; Colpan, L.; Nas, K.; Cevik, R.; Sarac, J.; Erdogan, F.; Duz, M.Z. The role of trace minerals in the pathogenesis of postmenopausal osteoporosis and a new effect of calcitonin. J. Bone Miner. Metab. 2002, 20, 39–43. [Google Scholar] [CrossRef]
- Lowe, N.M.; Fraser, W.D.; Jackson, M.J. Is there a potential therapeutic value of copper and zinc for osteoporosis? Proc. Nutr.Soc. 2002, 61, 181–185. [Google Scholar] [CrossRef]
- Devine, A.; Rosen, C.; Mohan, S.; Baylink, D.; Prince, R.L. Effects of zinc and other nutritional factors on insulin-like growth factor I and insulin-like growth factor binding proteins in postmenopausal women. Am. J. Clin. Nutr. 1998, 68, 200–206. [Google Scholar]
- Rodondi, A.; Ammann, P.; Ghilardi-Beuret, S.; Rizzoli, R. Zinc increases the effects of essential amino acids-whey protein supplements in frail elderly. J. Nutr. Health Aging 2009, 6, 491–497. [Google Scholar]
- Powell, S.R. The antioxidant properties of zinc. J. Nutr. 2000, 130, 1447S–1454S. [Google Scholar]
- Klotz, L.O.; Kröncke, K.D.; Buchczyk, D.P.; Sies, H. Role of copper, zinc, selenium and tellurium in the cellular defense against oxidative and nitrosative stress. J. Nutr. 2003, 133, 1448S–1451S. [Google Scholar]
- Bettger, W.J. Zinc and selenium, site specific vs. general antioxidant. Can. J. Physiol. Pharmacol. 1993, 71, 721–724. [Google Scholar] [CrossRef]
- Abdallah, S.M.; Samman, S. The effect of increasing dietary zinc on the activity of superoxide dismutase and zinc concentration in erythrocytes of healthy female subjects. Eur. J. Clin. Nutr. 1993, 47, 327–332. [Google Scholar]
- Department of Community Services and Health, National Dietary Survey of Adults: 1983. No. 2. Nutrient intakes, 1st ed.; Australian Government Publishing Service: Canberra, Australia, 1987.
- Larsson, S.C.; Wolk, K.; Brismar, K.; Wolk, A. Association of diet with serum insulin-like growth factor I in middle-aged and elderly men. Am. J. Clin. Nutr. 2005, 81, 1163–1167. [Google Scholar]
- King, J.C. Zinc: An essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679S–684S. [Google Scholar] [CrossRef]
- Lowe, N.M.; Medina, M.W.; Stammers, A.L.; Patel, S.; Souverein, O.W.; Dullemeijer, C.; Serra-Majem, L.; Nissensohn, M.; Hall Moran, V. The relationship between zinc intake and serum/plasma zinc concentration in adults: A systematic review and dose-response meta-analysis by the EURRECA Network. Br. J. Nutr. 2012, 108, 1962–1971. [Google Scholar] [CrossRef]
- Seeman, T.E.; Robbins, R.J. Aging and hypothalamic-pituitary-adrenal response to challenge in humans. Endocr. Rev. 1994, 15, 233–260. [Google Scholar]
- Maggio, M.; Ceda, G.P.; de Cicco, G.; Cattadori, E.; Visioli, S.; Ablondi, F.; Beghi, C.; Gherli, T.; Basaria, S.; Ceresini, G.; et al. Acute changes in circulating hormones in older patients with impaired ventricular function undergoing on-pump coronary artery bypass grafting. J. Parenter. Enter. Nutr. 2006, 30, 157–163. [Google Scholar] [CrossRef]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- Hollander, J.M.; Mechanick, J.I. Nutrition support and the chronic critical illness syndrome. Nutr. Clin. Pract. 2006, 21, 587–604. [Google Scholar] [CrossRef]
- Wolfe, R.R. Optimal nutrition, exercise, and hormonal therapy promote muscle anabolism in the elderly. J. Am. Coll. Surg. 2006, 202, 176–180. [Google Scholar] [CrossRef]
- Van den Berhe, G. The neuroendocrine stress response and modern intensive care: The concept revisited. Burns 1999, 25, 7–16. [Google Scholar] [CrossRef]
- Ferrando, A.A.; Paddon-Jones, D.; Wolfe, R.R. Bed rest and myopathies. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 410–415. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maggio, M.; De Vita, F.; Lauretani, F.; Buttò, V.; Bondi, G.; Cattabiani, C.; Nouvenne, A.; Meschi, T.; Dall'Aglio, E.; Ceda, G.P. IGF-1, the Cross Road of the Nutritional, Inflammatory and Hormonal Pathways to Frailty. Nutrients 2013, 5, 4184-4205. https://doi.org/10.3390/nu5104184
Maggio M, De Vita F, Lauretani F, Buttò V, Bondi G, Cattabiani C, Nouvenne A, Meschi T, Dall'Aglio E, Ceda GP. IGF-1, the Cross Road of the Nutritional, Inflammatory and Hormonal Pathways to Frailty. Nutrients. 2013; 5(10):4184-4205. https://doi.org/10.3390/nu5104184
Chicago/Turabian StyleMaggio, Marcello, Francesca De Vita, Fulvio Lauretani, Valeria Buttò, Giuliana Bondi, Chiara Cattabiani, Antonio Nouvenne, Tiziana Meschi, Elisabetta Dall'Aglio, and Gian Paolo Ceda. 2013. "IGF-1, the Cross Road of the Nutritional, Inflammatory and Hormonal Pathways to Frailty" Nutrients 5, no. 10: 4184-4205. https://doi.org/10.3390/nu5104184
APA StyleMaggio, M., De Vita, F., Lauretani, F., Buttò, V., Bondi, G., Cattabiani, C., Nouvenne, A., Meschi, T., Dall'Aglio, E., & Ceda, G. P. (2013). IGF-1, the Cross Road of the Nutritional, Inflammatory and Hormonal Pathways to Frailty. Nutrients, 5(10), 4184-4205. https://doi.org/10.3390/nu5104184