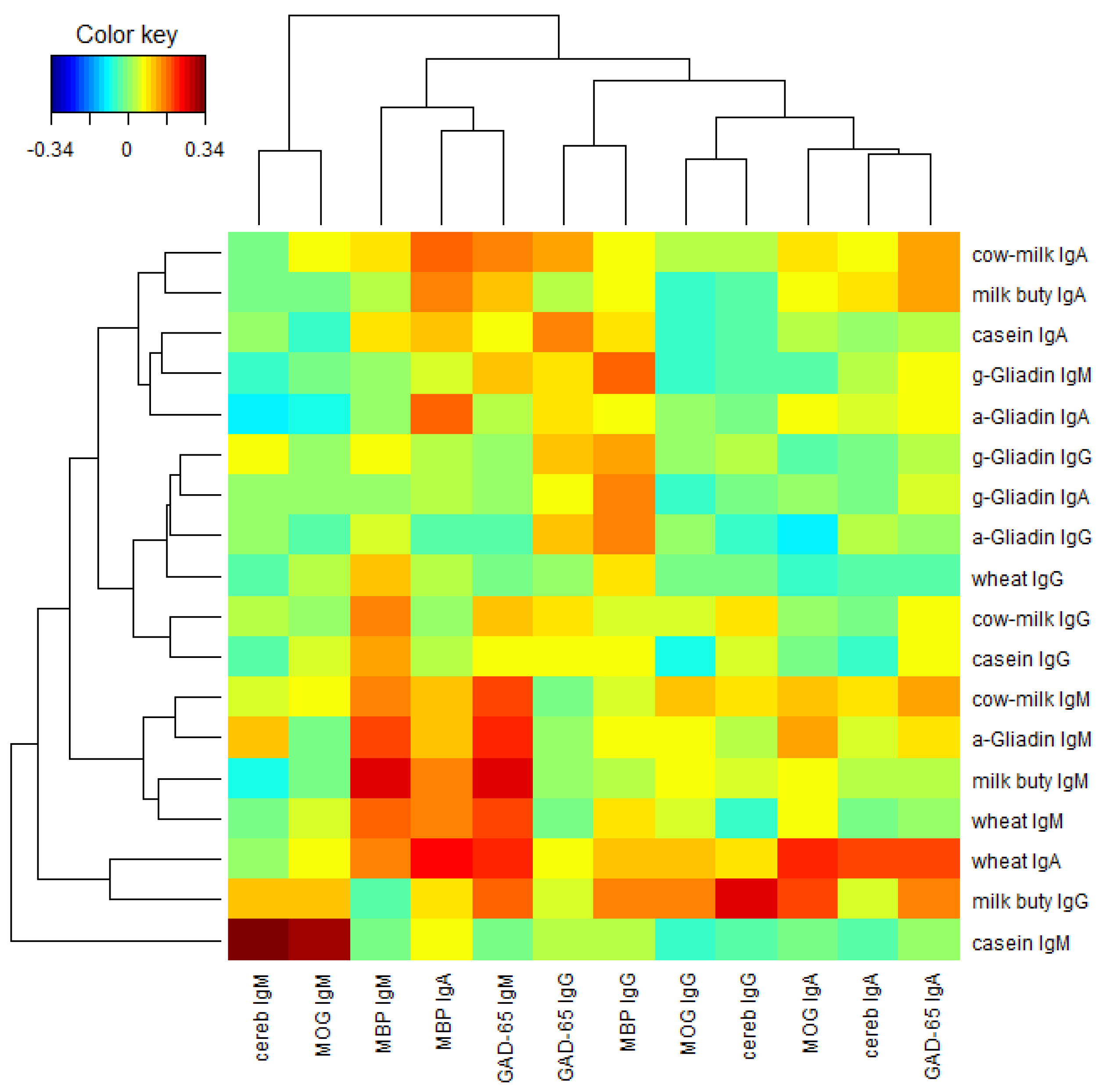
The Prevalence of Antibodies against Wheat and Milk Proteins in Blood Donors and Their Contribution to Neuroimmune Reactivities




Abstract
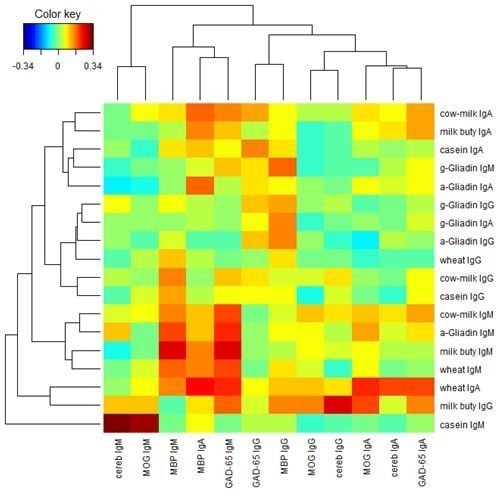
{kind=link}
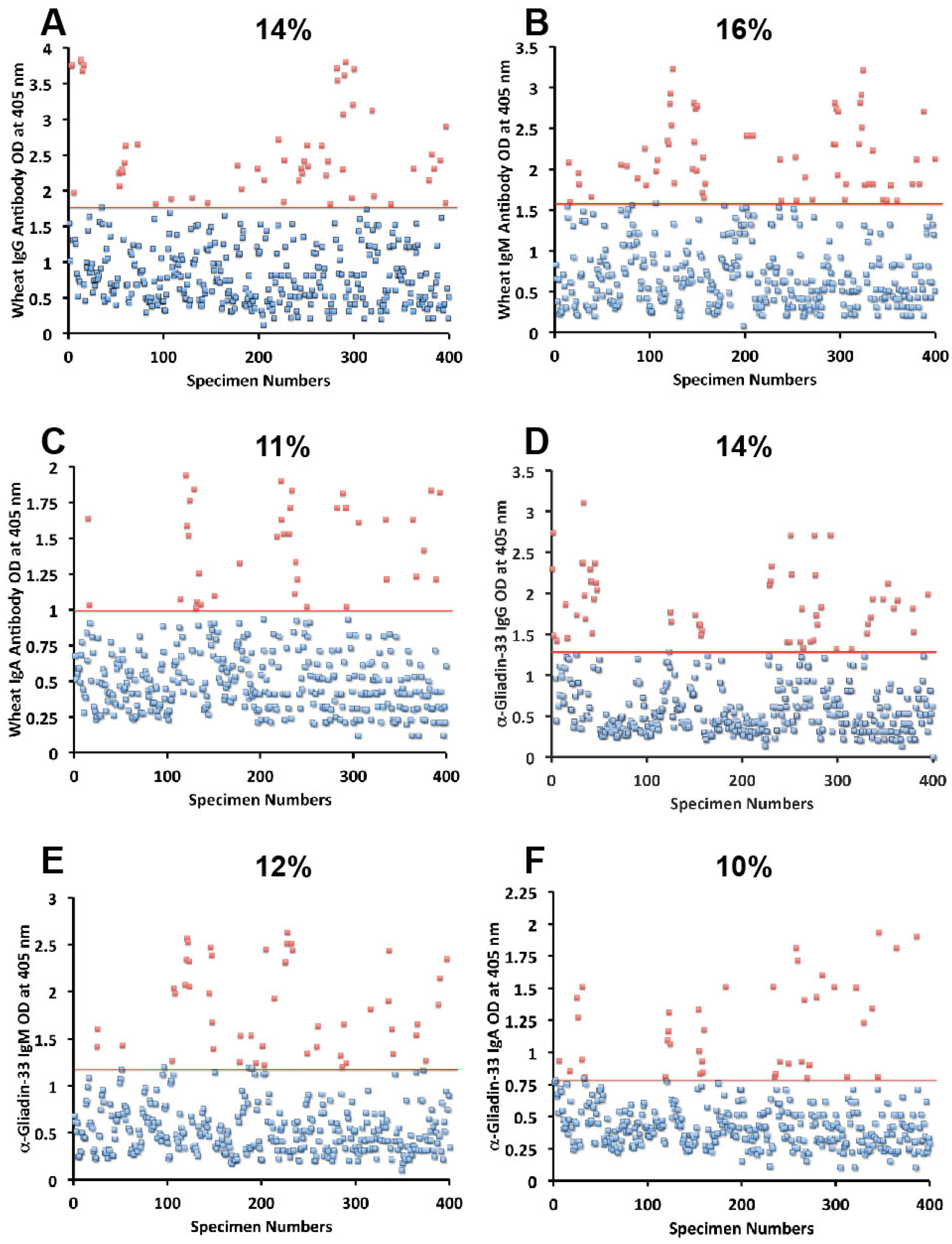{kind=link}
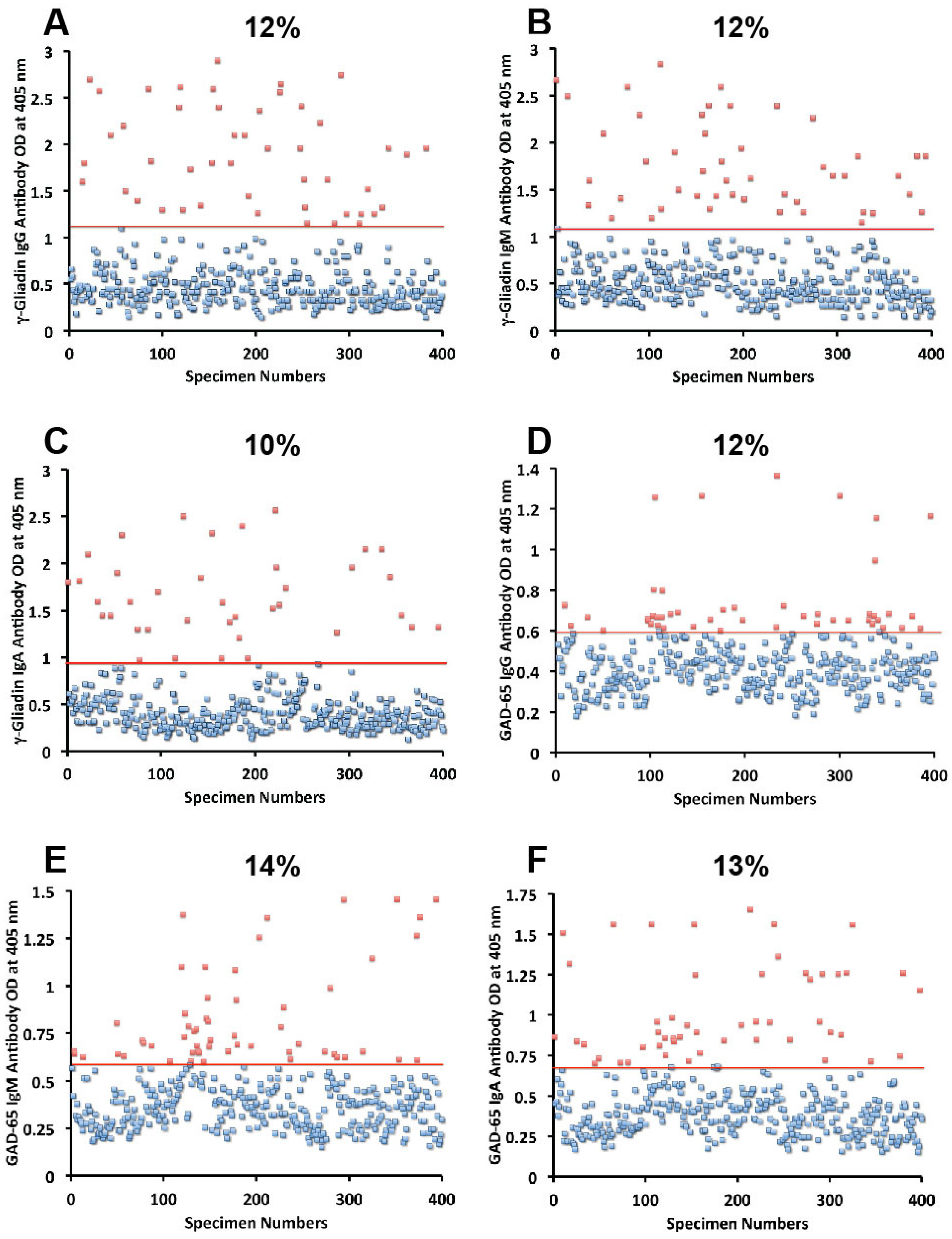{kind=link}
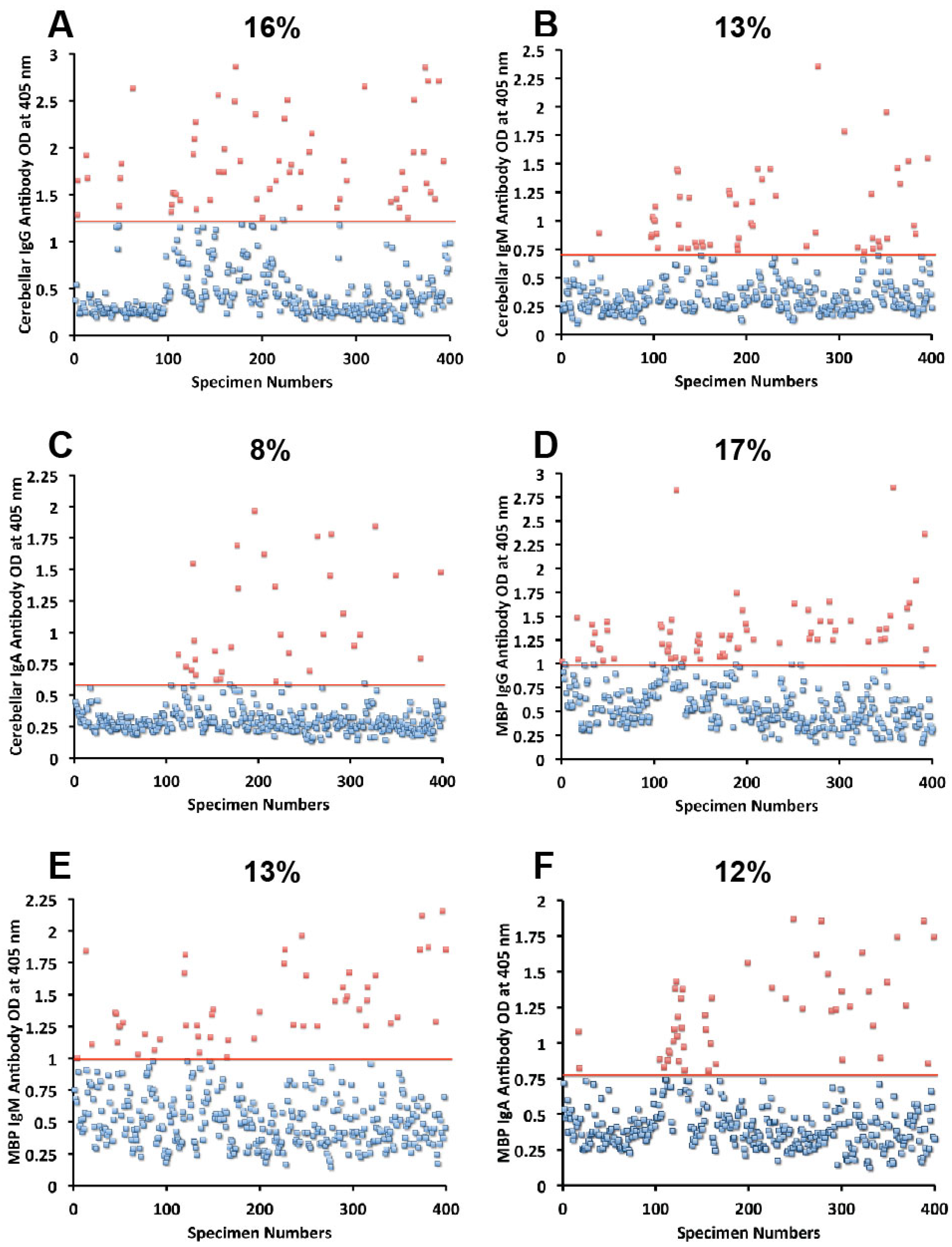{kind=link}
{kind=link}
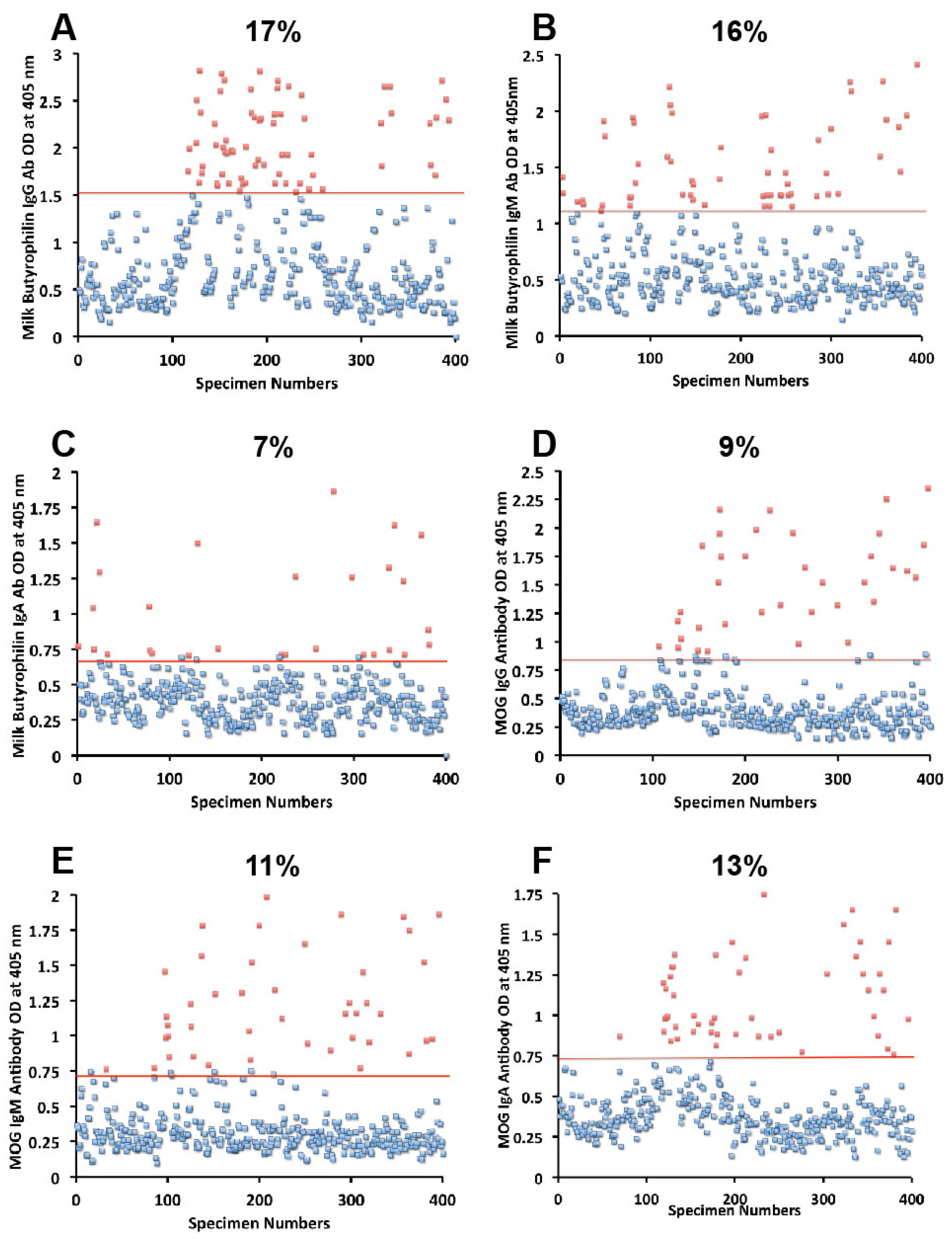{kind=link}
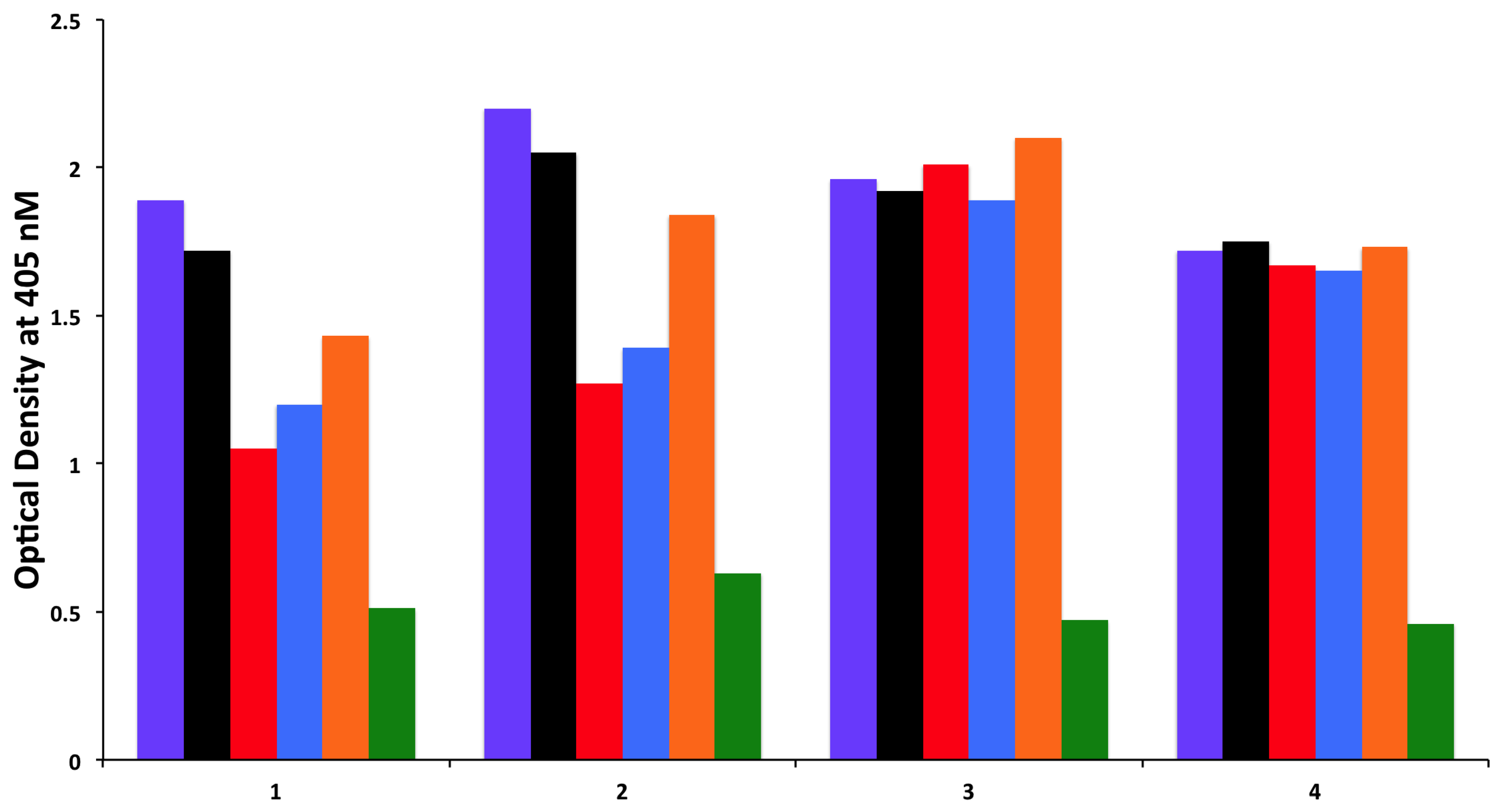{kind=link}
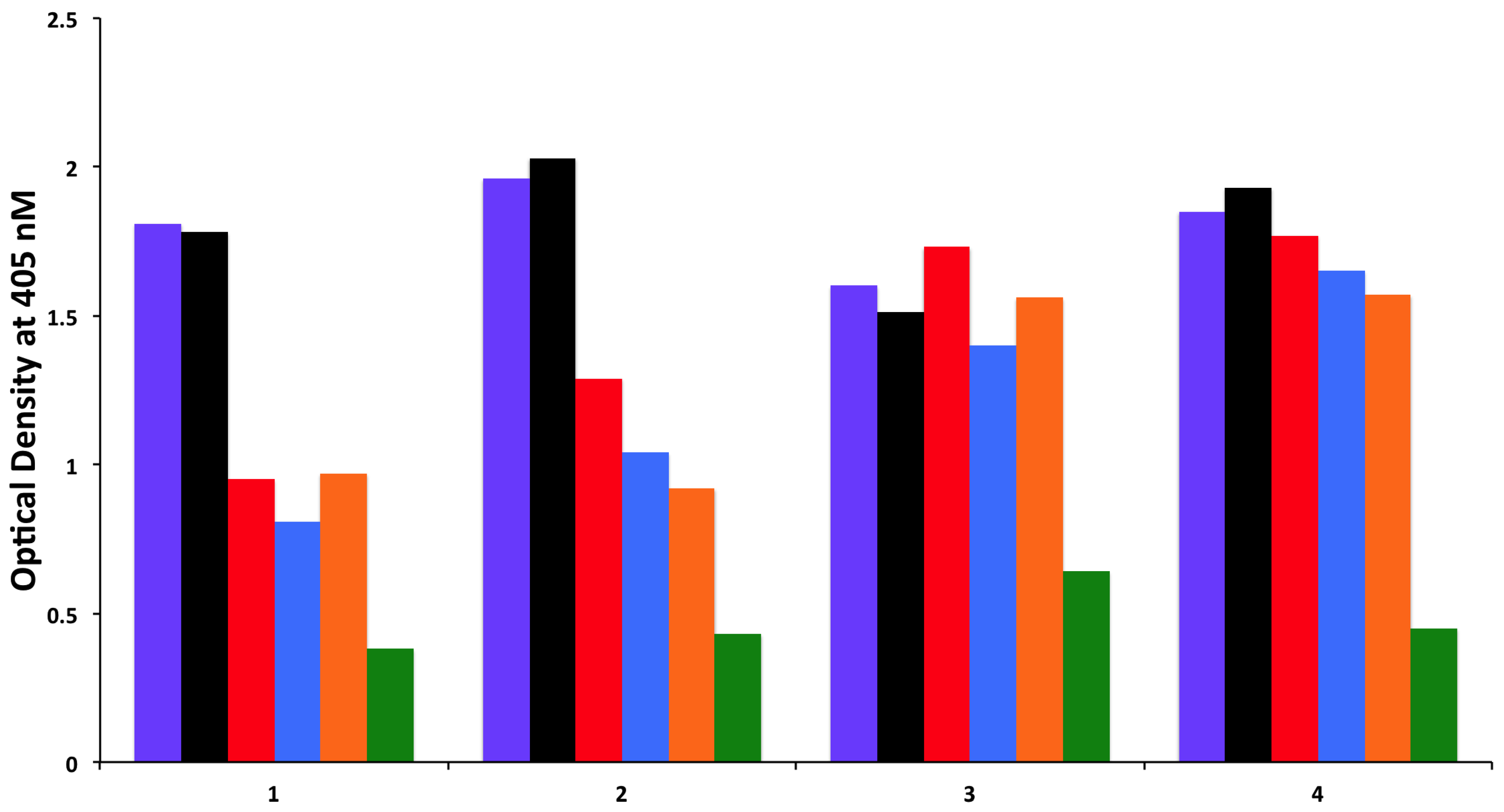{kind=link}
{kind=link}
Share and Cite
Vojdani, A.; Kharrazian, D.; Mukherjee, P.S. The Prevalence of Antibodies against Wheat and Milk Proteins in Blood Donors and Their Contribution to Neuroimmune Reactivities. Nutrients 2014, 6, 15-36. https://doi.org/10.3390/nu6010015
Vojdani A, Kharrazian D, Mukherjee PS. The Prevalence of Antibodies against Wheat and Milk Proteins in Blood Donors and Their Contribution to Neuroimmune Reactivities. Nutrients. 2014; 6(1):15-36. https://doi.org/10.3390/nu6010015
Chicago/Turabian StyleVojdani, Aristo, Datis Kharrazian, and Partha Sarathi Mukherjee. 2014. "The Prevalence of Antibodies against Wheat and Milk Proteins in Blood Donors and Their Contribution to Neuroimmune Reactivities" Nutrients 6, no. 1: 15-36. https://doi.org/10.3390/nu6010015
APA StyleVojdani, A., Kharrazian, D., & Mukherjee, P. S. (2014). The Prevalence of Antibodies against Wheat and Milk Proteins in Blood Donors and Their Contribution to Neuroimmune Reactivities. Nutrients, 6(1), 15-36. https://doi.org/10.3390/nu6010015