Therapeutic Effect of Polysaccharide of Large Yellow Croaker Swim Bladder on Lupus Nephritis of Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Polysaccharide of Large Yellow Croaker Swim Bladder (PLYCSB) Preparation
2.2. Lupus Nephritis Experiment
2.3. Analysis of Inflammation-Related Cytokines in Serum by Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Serum Levels of SCr (Serum Creatinine), BUN (Blood Urea Nitrogen), TC (Total Cholesterol), TG (Triglyceride), TP (Total protein) and ALB (Albumin) Determination
2.5. Auto-Antibody ds-DNA Assay
2.6. Urinary Protein Excretion Test
2.7. Histology Assay
2.8. Reverse Transcription-Polymerase Chain reaction (RT-PCR) of Inflammation—Related Gene Expression in the Kidney Tissue
2.9. Protein Extraction and Western Blot Analysis in the Kidney Tissue
2.10. Statistical Analysis
3. Results
3.1. Cytokine IL-6, IL-12, TNF-α and IFN-γ Levels

{kind=link}
{kind=link}
{kind=link}
| Group | IL-6 (pg/mL) | IL-12 (pg/mL) | TNF-α (pg/mL) | IFN-γ (pg/mL) |
|---|---|---|---|---|
| Normal | 51.34 ± 3.63 a | 432.65 ± 41.26 A | 110.36 ± 6.68 a | 41.27 ± 2.98 A |
| Control | 308.95 ± 31.23 b | 1711.56 ± 89.91 B | 731.83 ± 13.60 b | 127.28 ± 8.13 B |
| PLYCSB | ||||
| 25 mg/kg | 201.46 ± 22.37 c | 1227.10 ± 77.21 C | 537.83 ± 19.55 c | 81.43 ± 5.44 C |
| 50 mg/kg | 131.61 ± 20.43 d | 871.74 ± 32.36 D | 301.18 ± 14.76 d | 63.65 ± 6.35 D |
3.2. Serum SCr, BUN, TC, TG, TP and ALB Levels
| Group | SCr (μmol/L) | BUN (mmol/L) | TC (mmol/L) | TG (mmol/L) | TP (g/L) | ALB (g/L) |
|---|---|---|---|---|---|---|
| Normal | 61.21 ± 4.21 a | 1.44 ± 0.36 A | 1.71 ± 0.23 d | 0.97 ± 0.12 A | 52.27 ± 3.28 b | 35.18 ± 1.38 B |
| Control | 134.72 ± 17.88 b | 14.87 ± 2.12 B | 16.42 ± 2.17 a | 14.76 ± 1.28 B | 22.82 ± 1.65 a | 12.27 ± 0.22 A |
| PLYCSB | ||||||
| 25 mg/kg | 107.82 ± 2.86 c | 7.22 ± 1.18 C | 12.67 ± 2.97 b | 8.39 ± 1.26 C | 35.91 ± 1.88 d | 20.09 ± 0.98 D |
| 50 mg/kg | 80.37 ± 2.77 d | 4.62 ± 0.87 D | 6.26 ± 0.67 c | 4.51 ± 1.19 D | 42.67 ± 1.20 c | 28.17 ± 0.68 C |
3.3. ds-DNA Positive Rate during the Experiment
| Group | 2 Weeks | 4 Weeks | 6 Weeks | 8 Weeks | 10 Weeks | 12 Weeks |
|---|---|---|---|---|---|---|
| Normal | 0/10 a (0%) | 0/10 A (0%) | 0/10 d (0%) | 0/10 D (0%) | 0/10 c (0%) | 0/10 C (0%) |
| Control | 5/10 b (50%) | 8/10 B (80%) | 10/10 b (100%) | 10/10 B (100%) | 10/10 b (100%) | 10/10 B (100%) |
| PLYCSB | ||||||
| 25 mg/kg | 4/10 c (40%) | 6/10 C (60%) | 8/10 c (80%) | 10/10 B (100%) | 10/10 b (100%) | 10/10 B (100%) |
| 50 mg/kg | 4/10 c (40%) | 5/10 D (50%) | 6/10 a (60%) | 7/10 C (70%) | 10/10 b (100%) | 10/10 B (100%) |
3.4. Change of Urine Protein during the Experiment
| Group | 2 Weeks | 4 Weeks | 6 Weeks | 8 Weeks | 10 Weeks | 12 Weeks |
|---|---|---|---|---|---|---|
| Normal | 0.73 ± 0.09 a | 0.79 ± 0.11 A | 0.82 ± 0.06 a | 0.88 ± 0.10 A | 0.87 ± 0.11 a | 0.90 ± 0.04 A |
| Control | 5.75 ± 0.63 b | 6.22 ± 0.54 B | 7.87 ± 0.81 b | 8.24 ± 0.72 B | 8.88 ± 0.50 b | 9.12 ± 1.01 B |
| PLYCSB | ||||||
| 25 mg/kg | 3.26 ± 0.41 c | 4.03 ± 0.61 C | 4.50 ± 0.36 c | 5.11 ± 0.53 C | 6.05 ± 0.39 c | 6.39 ± 0.41 C |
| 50 mg/kg | 2.02 ± 0.27 d | 2.35 ± 0.19 D | 2.72 ± 0.11 d | 3.12 ± 0.23 D | 3.41 ± 0.07 d | 4.89 ± 0.38 D |
3.5. Glomerular Number and Glomerular Sclerosis Index by Histological Analysis
| Group | Number of Glomerular | Glomerular Sclerosis Index |
|---|---|---|
| Normal | 35 ± 3 a | 0.00 ± 0.00 A |
| Control | 13 ± 3 b | 3.12 ± 0.96 B |
| PLYCSB | ||
| 25 mg/kg | 22 ± 2 c | 2.03 ± 0.45 C |
| 50 mg/kg | 27 ± 3 d | 1.57 ± 0.32 D |
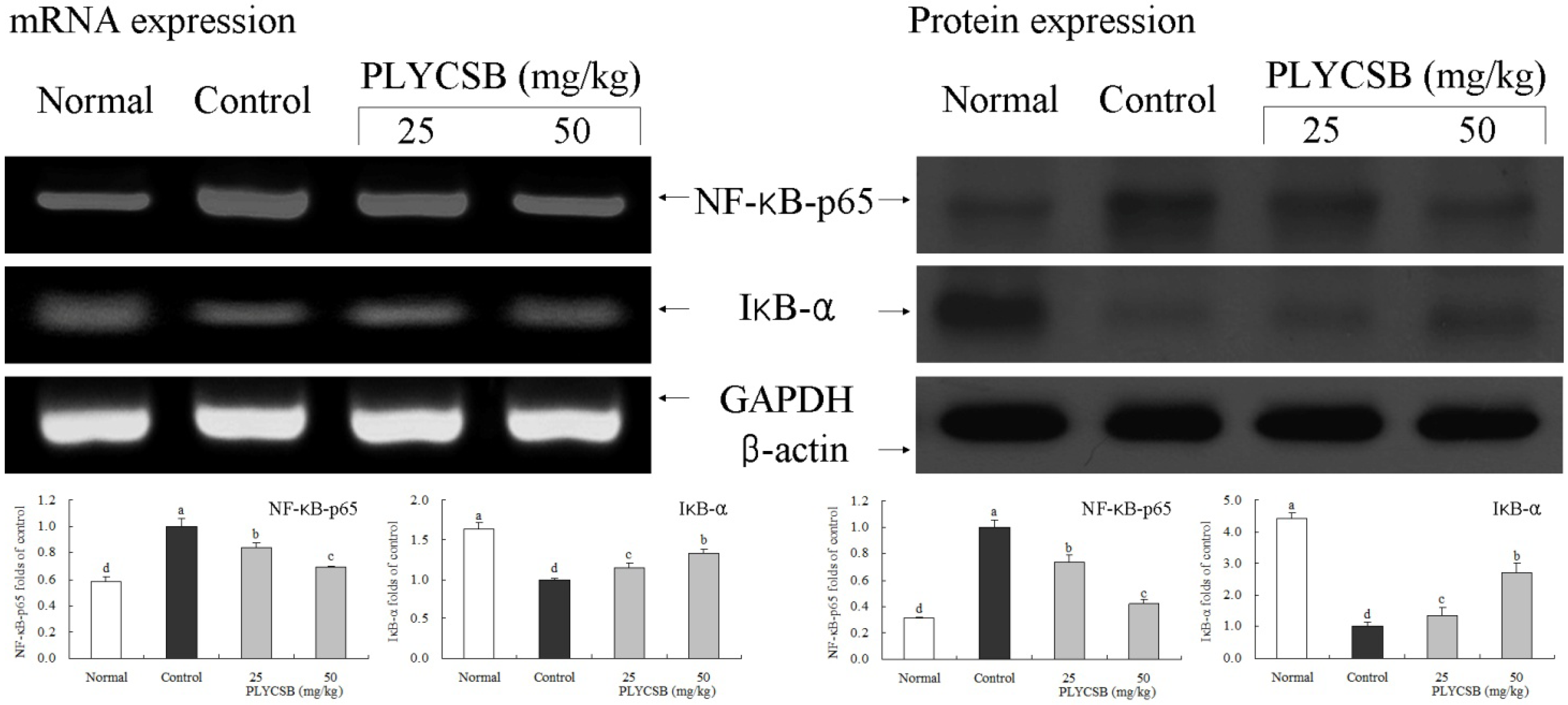
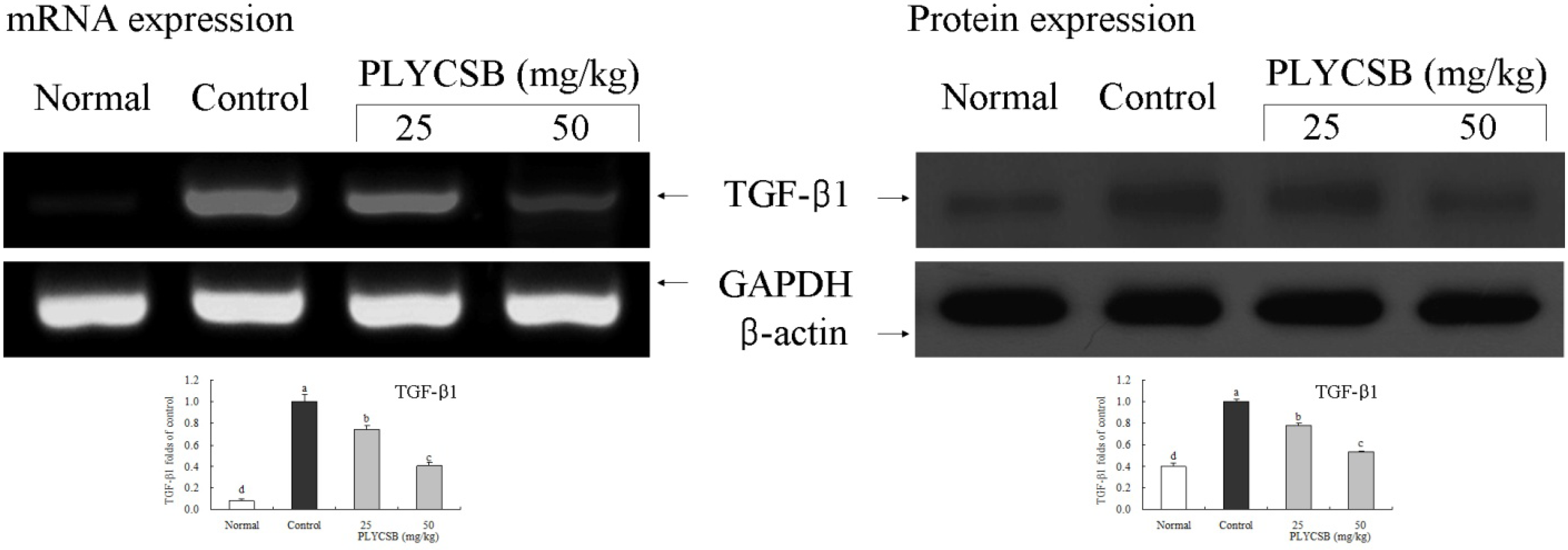
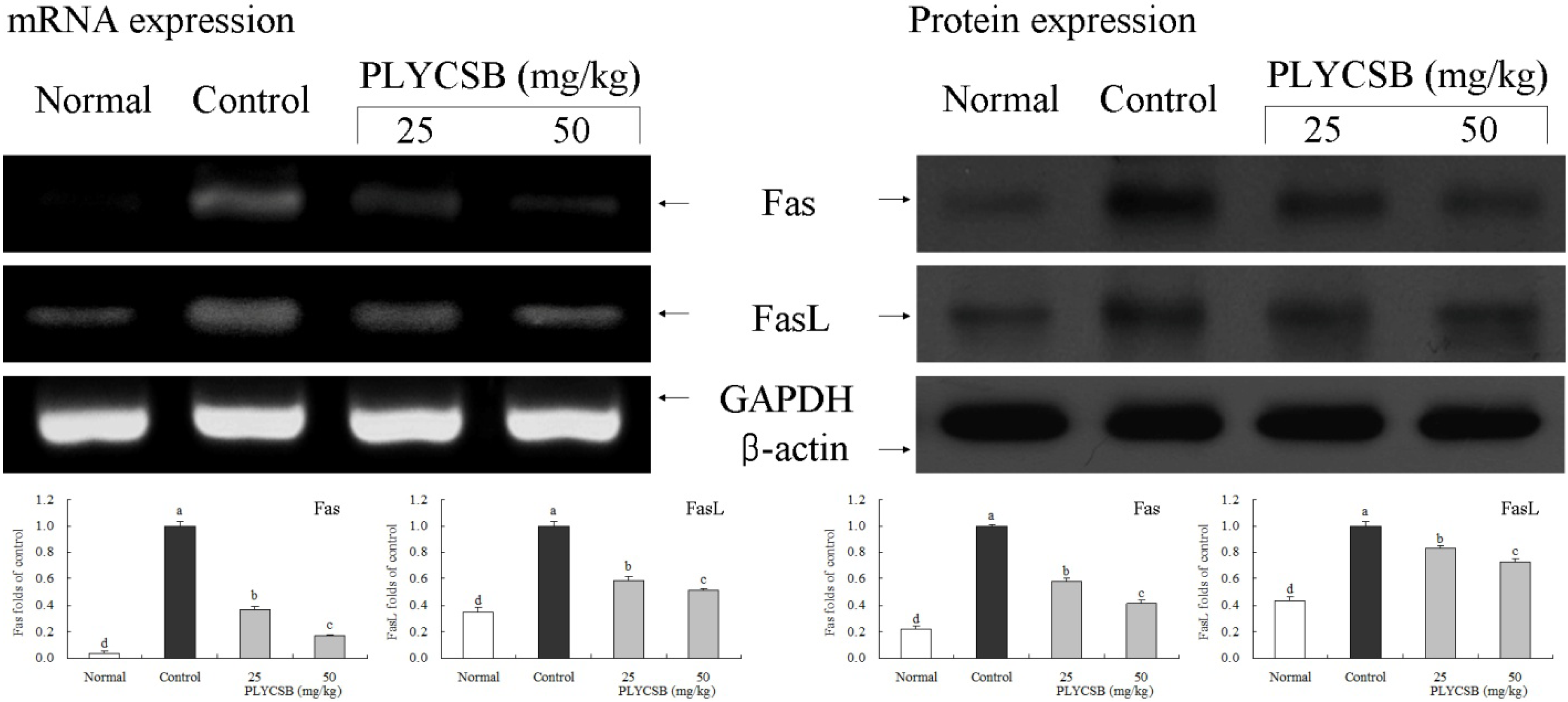
3.6. Gene Expression of NF-κB-p65, IκB-α, TGF-β1, Fas and FasL



4. Discussion
5. Conclusions
Acknowledgments
References
- Jian, J.C.; Wu, Z.H. Effects of traditional Chinese medicine on nonspecific immunity and disease resistance of large yellow croaker, Pseudosciaena crocea (Richardson). Aquaculture 2003, 218, 1–9. [Google Scholar] [CrossRef]
- Li, C.; Yao, C.L. Molecular and expression characterizations of interleukin-8 gene in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2013, 34, 799–809. [Google Scholar] [CrossRef]
- Liu, S.; Yu, B. Peptides from variegated carp (Aristichthys nobilis) swim bladder: Fermentation production and assessment of antioxidant properties. Food Sci. 2009, 30, 332–334. [Google Scholar]
- Yu, Z.H.; Yin, L.H.; Yang, Q.; Liu, Y. Effect of Lentinus edodes polysaccharide on oxidative stress, immunity activity and oral ulceration of rats stimulated by phenol. Carbohydr. Polym. 2009, 75, 115–118. [Google Scholar] [CrossRef]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef]
- Zhang, J.J. Progress in diagnosis and treatment of lupus nephritis. Acad. J. Kunming Med. Coll. 2008, 29. [Google Scholar] [CrossRef]
- Ma, H.L.; Zhang, X.G.; Zhang, X.Z.; Xu, Y. Effect of esculentoside A on therapeutic and cytokines secretion of lupus nephritis in BXSB mice. Chongqing Med. J. 2012, 41, 2954–2955. [Google Scholar]
- Bruijn, J.A.; van Elven, E.H.; Hogendoorn, P.C.; Corver, W.E.; Hoedemaeker, P.J.; Fleuren, G.J. Murine chronic graft-versus-host disease as a model for lupus nephritis. Am. J. Pathol. 1988, 130, 639–641. [Google Scholar]
- Yan, C.; Yan, X. Study on extraction of Lycium barbarum polysaccharides by different methods and their antioxidant effects in vitro. Food Sci. 2008, 29, 183–187. [Google Scholar]
- Bo, Y.H.; Jiang, F.; Su, W.; Ling, Y.L.; Bi, Z.Q. The protective effect of idiotypic peptide on kidney in the GVHD lupus nephritis mice. J. Cap. Med. Univ. 2007, 28, 770–773. [Google Scholar]
- Zhao, X.; Kim, S.Y.; Park, K.Y. Bamboo salt has in vitro anticancer activity in HCT-116 cells and exerts anti-metastatic effects in vivo. J. Med. Food 2013, 16, 9–19. [Google Scholar] [CrossRef]
- Chen, L.H.; Song, J.L.; Qian, Y.; Zhao, X.; Suo, H.Y.; Li, J. Increased preventive effect on colon carcinogenesis by use of resistant starch (RS3) as the carrier for polysaccharide of Larimichthys crocea swimming bladder. Int. J. Mol. Sci. 2014, 15, 817–829. [Google Scholar] [CrossRef]
- Cao, H.; Tian, X.L.; Liu, X. Study on molecular identification and pharmacology of hemostasis action for isinglass. J. Chin. Inst. Food Sci. Technol. 2009, 9, 170–176. [Google Scholar]
- Li, X.Y.; Kong, F.Y.; Zhang, H.Q.; Tang, R.X.; Zheng, K.Y. The duplication and identification of anti-glomerular basement membrane (GBM) nephritis model in mice. Acta Acad. Med. Xuzhou 2011, 31, 527–530. [Google Scholar]
- Chou, A.; Zhou, J.Y.; Zhou, Y.; Hua, J.; Wu, J.B. Therapeutic effect of zhenwu decoction on chronic glomerulonephritis rat model induced by cationization bovine serum albumin osmotic pump. Tradit. Chin. Drug Res. Chin. Pharm. 2012, 23, 626–630. [Google Scholar]
- Chen, W. Changes of urine protein in patients with chronic glomerulonephritis after different doses of irbesartan treatment. Chin. J. Arterioscler. 2010, 18, 909–910. [Google Scholar]
- Du, J.; Wang, Q.S.; Jia, R.H. Glomerular cell proliferation and apoptosis experimental glomerulosclerosis. J. Pract. Med. 2005, 21, 1623–1625. [Google Scholar]
- Wang, M.F. Significance of anti-double stranded DNA in system lupus erythematosus. Chin. J. Health Lab. Technol. 2010, 20, 606–607. [Google Scholar]
- Philip, S.; Bulbule, A.; Kundu, G.C. Matrix metalloproteinase-2: Mechanism and regulation of NF-κB-mediated activation and its role in cell motility and ECM-invasion. Glycocorj. J. 2004, 21, 429–441. [Google Scholar] [CrossRef]
- Yao, C.W.; Tang, D.S.; Liang, D. Expression and significance of NF-κB in mouse renal tissue with lupus nephritis-prone BXSB. J. Guangdong Med. Coll. 2005, 23, 493–495. [Google Scholar]
- Guijarro, C.; Kim, Y.; Kasiske, B.L. Central role of the transcription factor NF-κB in mesangial cell production of chemokines. Contrib. Nephrol. 1997, 120, 210–218. [Google Scholar] [CrossRef]
- Rangan, G.K.; Wang, Y.; Tay, Y.C.; Harris, D.C. Cytokine gene expression in Adriamycin nephropathy: Effects of antioxidant nuclear factor κB inhibitors in established disease. Nephron 2000, 86, 482–490. [Google Scholar] [CrossRef]
- Zhou, P.; Chen, X.R.; Li, S.M.; Tu, S.Q. Expression of transforming growth Factor-β genes in kidneys of BXSB lupus mice. Chin. J. Dermatol. Venereol. 1999, 13, 77–78. [Google Scholar] [CrossRef]
- Li, Z.J.; Li, Y.J.; Yang, Q.Q.; Yang, X.; Xu, Y.W.; Yu, X.Q. Significance of levels IL-12 and IgG in patients with lupus nephritis. J. New Med. 2002, 33, 19–20. [Google Scholar]
- Xiang, L.; Gao, X.X.; Pan, J.R. Serum levels of interferon-gamma and interleukin-10 in patients with chronic glomerulonephritis and their clinical significance. Clin. Med. J. China 2006, 13, 269–271. [Google Scholar]
- Wang, Q.; Sun, P.; Li, G.J.; Zhu, K.; Wang, C.; Zhao, X. Inhibitory effects of Dendrobium candidum Wall ex Lindl. on azoxymethane- and dextran sulfate sodium-induced colon carcinogenesis in C57BL/6 mice. Oncol. Lett. 2014, 7, 493–498. [Google Scholar]
- Takahashi, T.; Tanaka, M.; Inazawa, J.; Abe, T.; Suda, T.; Nagata, S. Human Fas ligand: Gene structure, chromosomal location and species specificity. Int. Immunol. 1994, 6, 1567–1574. [Google Scholar] [CrossRef]
- Itoh, N.; Yonehara, S.; Ishii, A.; Yonehara, M.; Mizushima, S.; Sameshima, M.; Hase, A.; Seto, Y.; Nagata, S. The polypeptide encoded by the cDNA for human cell surface antigen Fas can mediate apoptosis. Cell 1991, 66, 233–243. [Google Scholar] [CrossRef]
- Jiang, J.P.; Yi, Z.H.; Liu, Z.R.; Hu, L.P.; Hou, F.F. Expression Fas on monocytes and plasma soluble FasL in patients with lupus nephritis. Chin. J. Rheumatol. 2001, 5, 168–171. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jiang, X.; Zhao, X.; Luo, H.; Zhu, K. Therapeutic Effect of Polysaccharide of Large Yellow Croaker Swim Bladder on Lupus Nephritis of Mice. Nutrients 2014, 6, 1223-1235. https://doi.org/10.3390/nu6031223
Jiang X, Zhao X, Luo H, Zhu K. Therapeutic Effect of Polysaccharide of Large Yellow Croaker Swim Bladder on Lupus Nephritis of Mice. Nutrients. 2014; 6(3):1223-1235. https://doi.org/10.3390/nu6031223
Chicago/Turabian StyleJiang, Xianhong, Xin Zhao, Huali Luo, and Kai Zhu. 2014. "Therapeutic Effect of Polysaccharide of Large Yellow Croaker Swim Bladder on Lupus Nephritis of Mice" Nutrients 6, no. 3: 1223-1235. https://doi.org/10.3390/nu6031223