Urinary Metabolite Profiles in Premature Infants Show Early Postnatal Metabolic Adaptation and Maturation

Abstract
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
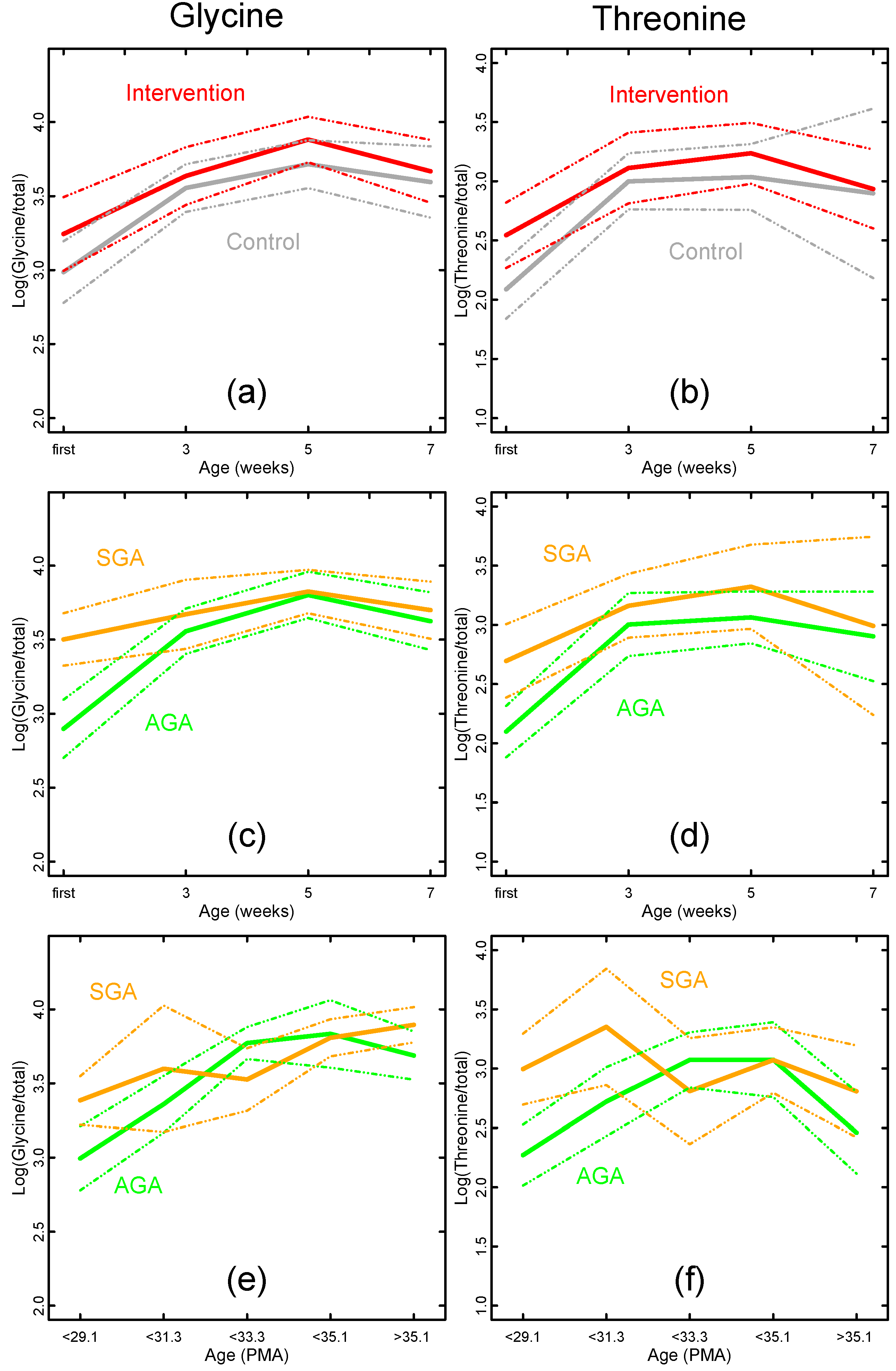{kind=link}
| Intervention (n = 23) | Control (n = 21) | p | |
|---|---|---|---|
| Human milk, mL/kg/day | 133 (110–139) | 134 (124–141) | 0.37 |
| Energy, kcal/kg/day | 139 (128–145) | 126 (121–128) | <0.001 |
| Protein, g/kg/day | 4.0 (3.9–4.2) | 3.2 (3.1–3.3) | <0.001 |
| Lipids, g/kg/day | 7.3 (6.4–7.6) | 5.9 (5.6–6.1) | <0.001 |
| Carbohydrates, g/kg/day | 14.4 (13.4–14.8) | 14.7 (14.3–15.1) | 0.12 |
| Arachidonic acid, mg/kg/day | 68 (57–73) | 24 (23–25) | <0.001 |
| Docosahexaenoic acid, mg/kg/day | 87 (81–91) | 36 (34–38) | <0.001 |
| Vitamin A, μg/kg/day | 1300 (1105–1442) | 252 (238–257) | <0.001 |
2. Materials and Methods
2.1. Study Design and Population
| Intervention (n = 24) | Control (n = 24) | p | |
|---|---|---|---|
| Gestational age (weeks), mean (range) | 28.1 (25.0–33.6) | 28.5 (24.0–32.6) | 0.60 |
| Birth weight (g), mean (range) | 940 (460–1311) | 1083 (571–1414) | 0.03 |
| Small for gestational age, n (%) | 11/24 (46%) | 5/24 (21%) | 0.12 |
| Sex, boys, n (%) | 15/24 (63%) | 15/24 (63%) | 1.00 |
| Cesarean section, n (%) | 16/24 (67%) | 19/24 (79%) | 0.52 |
| APGAR-score, 5-min, mean (± SD) | 7.33 (± 1.7) | 7.54 (± 1.7) | 0.68 |
| Prenatal steroid treatment, n (%) | 22/24 (92%) | 24/24 (100%) | 0.49 |
| Late onset septicemia, n (%) | 15/24 (63%) | 7/24 (29%) | 0.04 |
| NEC, n (%) | 1/24 (4%) | 2/24 (8%) | 1.00 |
| IVH, grade ≥3, n (%) | 2/24 (9%) | 2/24 (9%) | 1.00 |
| PVL, grade ≥3, n (%) | 0/24 (0%) | 1/24 (4%) | 1.00 |
| ROP (severe grade III/+disease), n (%) a | 3/23 (13%) | 2/23 (9%) | 1.00 |
| O2 at 36 weeks PMA, n (%) a | 5/23 (22%) | 6/23 (26%) | 1.00 |
| PDA surgical treatment, n (%) | 4/24 (17%) | 2/24 (8%) | 0.67 |
| Deaths before 36 weeks PMA, n (%) | 1/24 (4%) | 1/24 (4%) | 1.00 |
| Hypophosphatemia 1st week, n (%) b | 17/22 (77%) | 6/23 (26%) | 0.001 |
| Hypokalemia 1st week, n (%) | 21/24 (88%) | 11/24 (46%) | 0.005 |
2.2. Nutritional Intervention
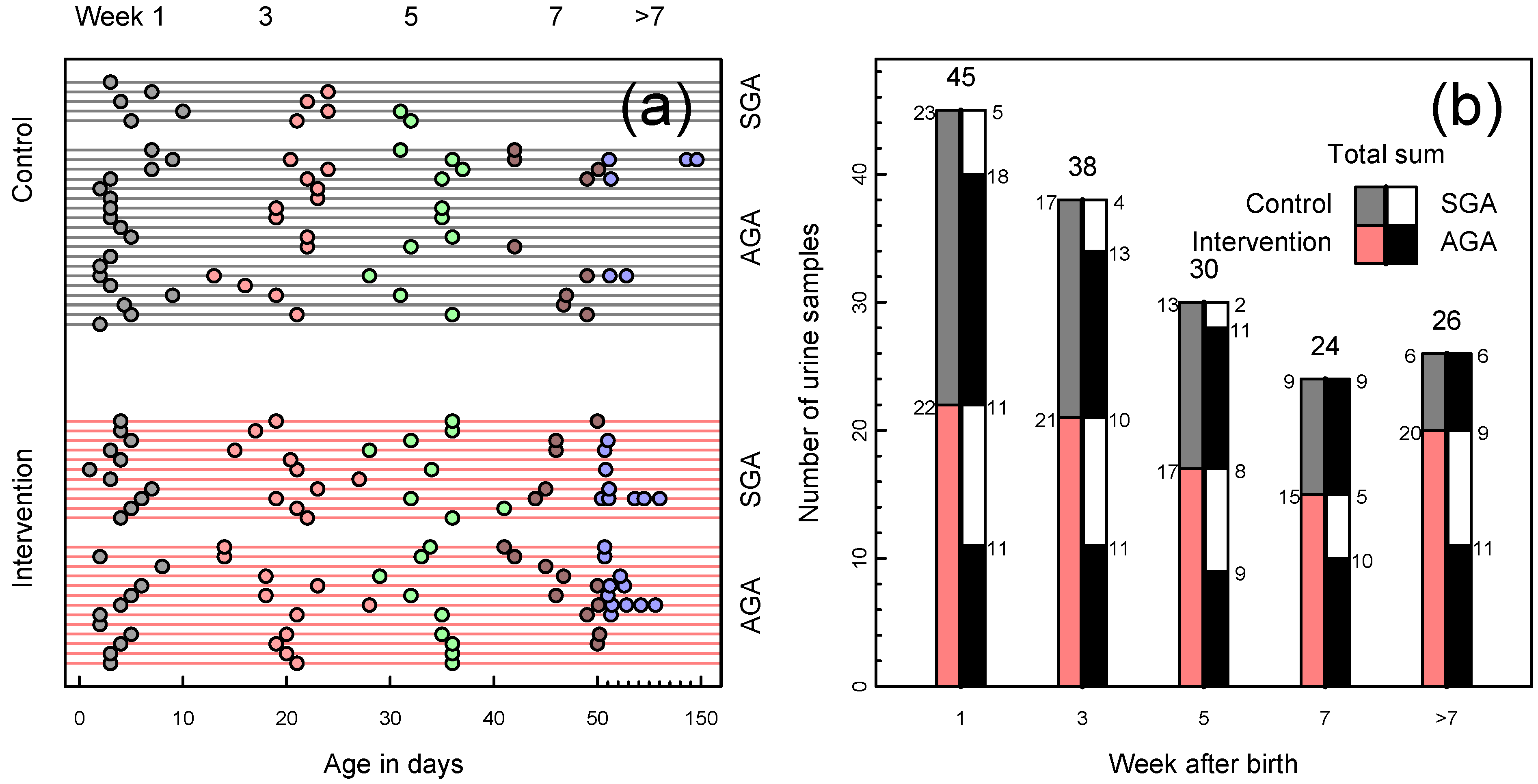
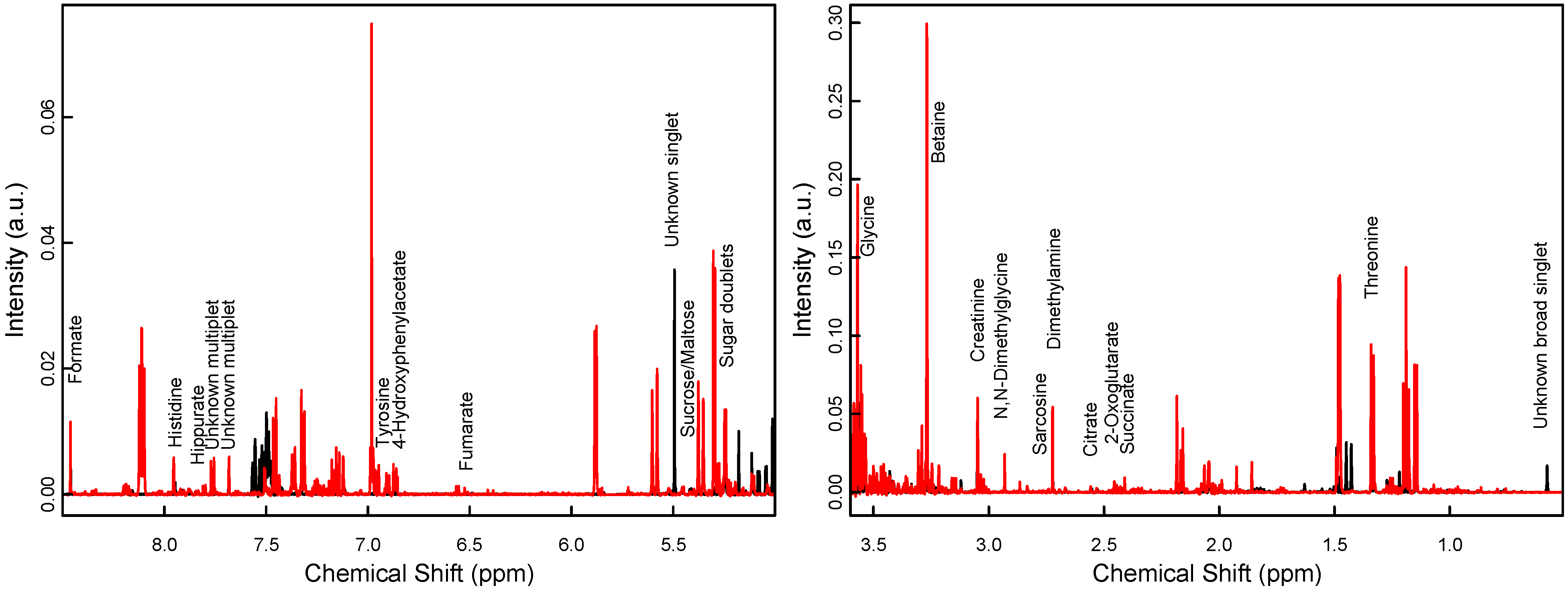
2.3. Sample Collection and Preparation


2.4. Statistical Analysis
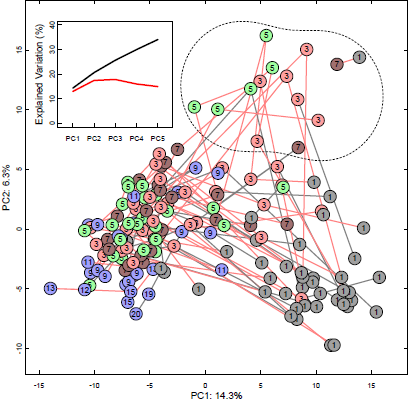
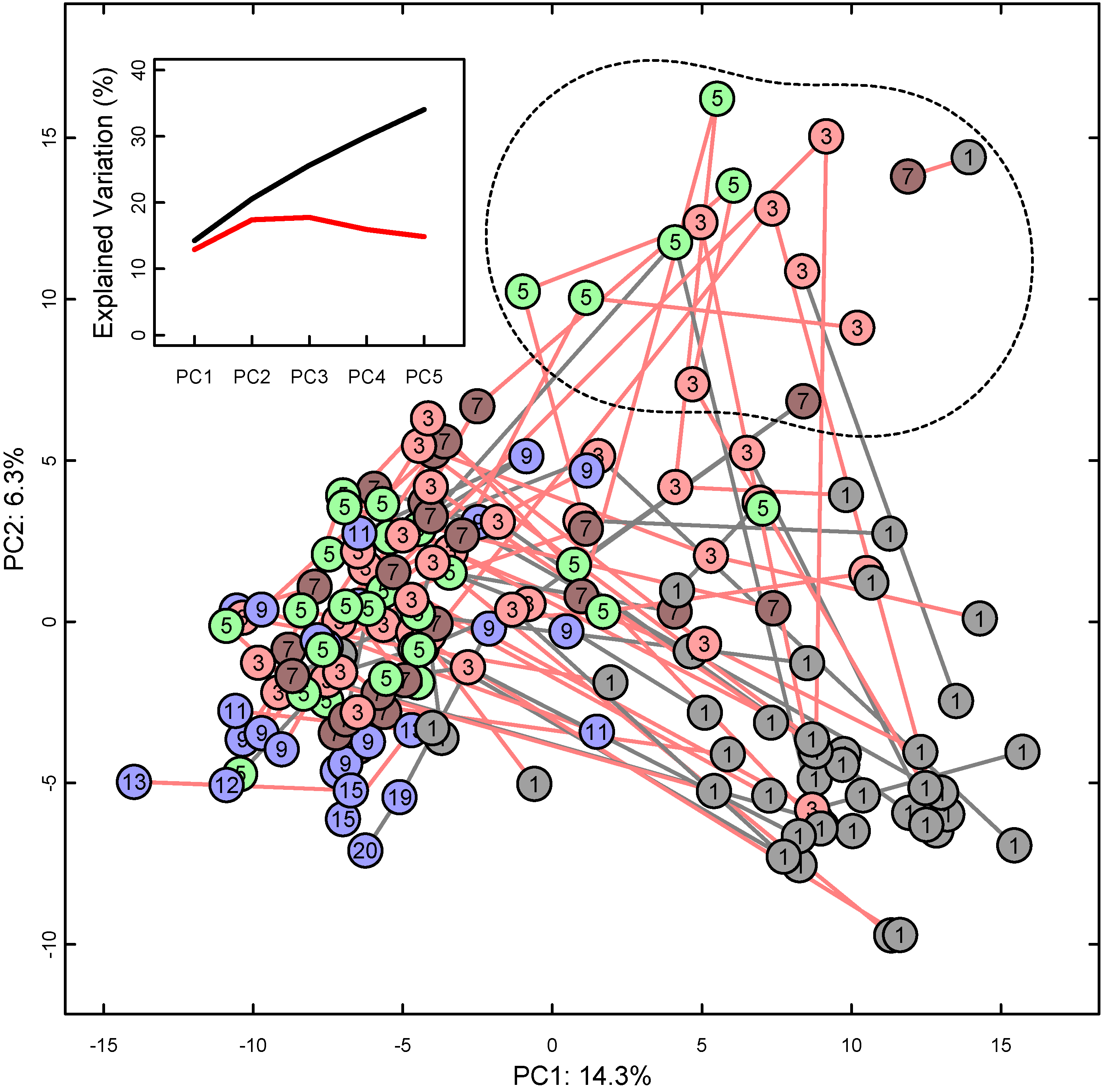
3. Results

| Metabolite | Weeks 1→7 Mixed Model | Weeks 1→3 | Weeks 3→5 | Weeks 5→7 | ||
|---|---|---|---|---|---|---|
| (n = 35 Pairs) | (n = 28 Pairs) | (n = 19 Pairs) | ||||
| p Interaction | p Diet | p Time | FC | FC | FC | |
| Total Integral | −1.2 | −1.2 | 1.2 | |||
| 1-Methylnicotinamide | 0.21 | 0.24 | 0.02 | 1.2 | 1.0 | −1.1 |
| 2-Oxoglutarate | 0.05 | 0.51 | <0.001 | 4.1 | 1.4 | 1.2 |
| 4-Hydroxyphenylacetate | 0.43 | 0.11 | <0.001 | 5.1 | −1.1 | −1.3 |
| Betaine | 0.96 | 0.16 | <0.001 | 1.5 | 1.2 | 1.3 |
| Citrate | 0.80 | 0.26 | <0.001 | 5.5 | 1.9 | 1.5 |
| Creatinine | 0.79 | 0.52 | 0.01 | 1.0 | 1.1 | 1.2 |
| Dimethylamine | 0.87 | 0.39 | 0.003 | 1.1 | 1.1 | 1.0 |
| Fumarate | 0.06 | 0.29 | <0.001 | 3.2 | 1.7 | 1.0 |
| Gluconate | - | - | - | x | - | - |
| Glycine | 0.62 | 0.05 | <0.001 | 1.6 | 1.2 | −1.1 |
| Hipurate | 0.56 | 0.10 | 0.31 | −1.2 | 1.6 | 1.0 |
| Histidine | 0.70 | 0.31 | <0.001 | 1.2 | 1.6 | −1.2 |
| myo-Inositol | 0.60 | 0.28 | 0.08 | 1.3 | 1.0 | −1.2 |
| N,N-Dimethylglycine | 0.23 | 0.62 | <0.001 | 1.3 | 1.2 | 1.3 |
| Succinate | 0.42 | 0.67 | <0.001 | 5.0 | 1.4 | 1.0 |
| Sucrose/Maltose | 0.12 | 0.51 | <0.001 | −8.4 | 1.1 | −1.1 |
| Sugar doublets, 5.23 ppm | 0.30 | 0.08 | 0.03 | 1.0 | −1.2 | −1.2 |
| Threonine | 0.58 | 0.34 | <0.001 | 2.0 | 1.0 | −1.7 |
| trans-4-Hydroxy-l-proline | 0.26 | 0.16 | <0.001 | 3.1 | 1.3 | 1.0 |
| Tyrosine | 0.33 | 0.37 | <0.001 | 3.7 | 1.0 | −1.4 |
| Unknown, 0.57 ppm | - | - | - | x | - | - |
| Unknown, 5.50 ppm | - | - | - | x | - | - |
| Unknown, 7.68 ppm | 0.60 | 0.48 | <0.001 | 1.2 | 1.1 | 1.1 |
| Unknown, 7.76 ppm | 0.43 | 0.9 | <0.001 | 1.6 | 1.4 | 1.3 |
| Variable | Samples a | A b | Q2 | R2 | Q2/R2 |
|---|---|---|---|---|---|
| Intervention | all | - | - | - | |
| first | - | - | - | ||
| 36 weeks PMA | - | - | - | ||
| SGA status | all | 3 | 34% | 84% | 0.40 |
| first | 1 | 27% | 50% | 0.53 | |
| 36 weeks PMA | - | - | - | ||
| Infections | all | - | - | - | |
| first | - | - | - | ||
| 36 weeks PMA | - | - | - | ||
| Age (since birth) | all | 2 | 41% | 81% | 0.51 |
| 36 weeks PMA | - | - | - | ||
| Age (PMA) | all | 3 | 67% | 89% | 0.75 |
| first | 1 | 54% | 70% | 0.76 | |
| Sex | all | - | - | - | |
| first | - | - | - | ||
| 36 weeks PMA | - | - | - |
| Metabolite a | SGA Alone b | PMA Alone c | Mutually Adjusted d | |||||
|---|---|---|---|---|---|---|---|---|
| (Week 1) | FC | p | FC | p | FC (SGA) | p (SGA) | FC (PMA) | p (PMA) |
| 2-Oxoglutarate | 1.6 | 0.15 | 1.3 | 4 × 10−4 | −1.1 | 0.81 | 1.3 | 0.001 |
| Betaine | 1.4 | 0.057 | 1.1 | 9 × 10−5 | 1.0 | 0.95 | 1.1 | 5 × 10−4 |
| Citrate | 1.9 | 0.12 | 1.3 | 0.001 | −1.1 | 0.87 | 1.3 | 0.004 |
| Creatinine | 1.8 | 0.018 | 1.2 | 3 × 10−7 | 1.0 | 0.86 | 1.2 | 6 × 10−6 |
| Dimethylamine | 1.5 | 0.036 | 1.2 | 2 × 10−4 | 1.1 | 0.69 | 1.1 | 0.001 |
| Formate | 1.6 | 0.083 | 1.2 | 0.005 | 1.1 | 0.66 | 1.1 | 0.022 |
| Glycine | 1.8 | 2 × 10−4 | 1.1 | 7 × 10−5 | 1.4 | 0.027 | 1.1 | 0.005 |
| Histidine | 2.0 | 8 × 10−4 | 1.2 | 9 × 10−6 | 1.4 | 0.091 | 1.2 | 6 × 10−4 |
| myo-Inositol | 1.5 | 0.062 | 1.1 | 0.001 | 1.4 | 0.73 | 1.1 | 0.006 |
| N,N-Dimethylglycine | 1.0 | 0.93 | 1.2 | 0.020 | −1.6 | 0.18 | 1.2 | 0.008 |
| Succinate | 2.1 | 0.039 | 1.3 | 3 × 10−4 | 1.2 | 0.63 | 1.3 | 0.003 |
| Sugar doublets, 5.23 ppm | −1.3 | 0.11 | −1.1 | 0.002 | −1.1 | 0.66 | −1.1 | 0.012 |
| Threonine | 1.8 | 0.003 | 1.1 | 0.040 | 1.6 | 0.033 | 1.0 | 0.37 |
| trans-4-Hydroxy-l-proline | 1.9 | 0.016 | 1.3 | 5 × 10−7 | 1.1 | 0.81 | 1.3 | 1 × 10−5 |

4. Discussion
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Lapillonne, A.; O’Connor, D.L.; Wang, D.; Rigo, J. Nutritional recommendations for the late-preterm infant and the preterm infant after hospital discharge. J. Pediatr. 2013, 162, S90–S100. [Google Scholar] [CrossRef]
- Henriksen, C.; Westerberg, A.C.; Ronnestad, A.; Nakstad, B.; Veierod, M.B.; Drevon, C.A.; Iversen, P.O. Growth and nutrient intake among very-low-birth-weight infants fed fortified human milk during hospitalisation. Br. J. Nutr. 2009, 102, 1179–1186. [Google Scholar] [CrossRef]
- Lapillonne, A.; Griffin, I.J. Feeding preterm infants today for later metabolic and cardiovascular outcomes. J. Pediatr. 2013, 162, S7–S16. [Google Scholar] [CrossRef]
- Lucas, A. Long-term programming effects of early nutrition—Implications for the preterm infant. J. Perinatol. 2005, 25 (Suppl. 2), S2–S6. [Google Scholar] [CrossRef]
- Koletzko, B.; Brands, B.; Poston, L.; Godfrey, K.; Demmelmair, H. Early nutrition programming of long-term health. Proc. Nutr. Soc. 2012, 71, 371–378. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C. Infant mortality, childhood nutrition, and ischaemic heart disease in England and Wales. Lancet 1986, 1, 1077–1081. [Google Scholar] [CrossRef]
- Patel, M.S.; Srinivasan, M. Metabolic programming as a consequence of the nutritional environment during fetal and the immediate postnatal periods. In Neonatal Nutrition and Metabolism, 2nd ed.; Thureen, P., Hay, W., Eds.; Cambridge University Press: Cambridge, UK, 2006; pp. 76–90. [Google Scholar]
- Singhal, A.; Lucas, A. Early origins of cardiovascular disease: Is there a unifying hypothesis? Lancet 2004, 363, 1642–1645. [Google Scholar] [CrossRef]
- Moltu, S.J.; Blakstad, E.W.; Strommen, K.; Almaas, A.N.; Nakstad, B.; Ronnestad, A.; Braekke, K.; Veierod, M.B.; Drevon, C.A.; Iversen, P.O.; et al. Enhanced feeding and diminished postnatal growth failure in very-low-birth-weight infants. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 344–351. [Google Scholar] [CrossRef]
- Moltu, S.J.; Strommen, K.; Blakstad, E.W.; Almaas, A.N.; Westerberg, A.C.; Braekke, K.; Ronnestad, A.; Nakstad, B.; Berg, J.P.; Veierod, M.B.; et al. Enhanced feeding in very-low-birth-weight infants may cause electrolyte disturbances and septicemia—A randomized, controlled trial. Clin. Nutr. 2013, 32, 207–212. [Google Scholar] [CrossRef]
- Koletzko, B.; Goulet, O.; Hunt, J.; Krohn, K.; Shamir, R. 1. Guidelines on paediatric parenteral nutrition of the european society of paediatric gastroenterology, hepatology and nutrition (ESPGHAN) and the european society for clinical nutrition and metabolism (ESPEN), supported by the european society of paediatric research (ESPR). J. Pediatr. Gastroenterol. Nutr. 2005, 41 (Suppl. 2), S1–S87. [Google Scholar]
- Agostoni, C.; Buonocore, G.; Carnielli, V.P.; de, C.M.; Darmaun, D.; Decsi, T.; Domellof, M.; Embleton, N.D.; Fusch, C.; Genzel-Boroviczeny, O.; et al. Enteral nutrient supply for preterm infants: Commentary from the European Society of Paediatric Gastroenterology, Hepatology and Nutrition Committee on Nutrition. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 85–91. [Google Scholar] [CrossRef]
- Moco, S.; Collino, S.; Rezzi, S.; Martin, F.P. Metabolomics perspectives in pediatric research. Pediatr. Res. 2013, 73, 570–576. [Google Scholar] [CrossRef]
- Emwas, A.H.; Salek, R.M.; Griffin, J.L.; Merzaban, J. NMR-based metabolomics in human disease diagnosis: Applications, limitations, and recommendations. Metabolomics 2013, 9, 1048–1072. [Google Scholar] [CrossRef]
- Holmes, E.; Wilson, I.D.; Nicholson, J.K. Metabolic phenotyping in health and disease. Cell 2008, 134, 714–717. [Google Scholar] [CrossRef]
- Ellis, D.I.; Dunn, W.B.; Griffin, J.L.; Allwood, J.W.; Goodacre, R. Metabolic fingerprinting as a diagnostic tool. Pharmacogenomics 2007, 8, 1243–1266. [Google Scholar] [CrossRef]
- Atzori, L.; Antonucci, R.; Barberini, L.; Griffin, J.L.; Fanos, V. Metabolomics: A new tool for the neonatologist. J. Matern. Fetal Neonatal Med. 2009, 22 (Suppl. 3), 50–53. [Google Scholar] [CrossRef]
- Fanos, V.; Antonucci, R.; Barberini, L.; Noto, A.; Atzori, L. Clinical application of metabolomics in neonatology. J. Matern. Fetal Neonatal Med. 2012, 25, 104–109. [Google Scholar]
- Dessi, A.; Atzori, L.; Noto, A.; Visser, G.H.A.; Gazzolo, D.; Zanardo, V.; Barberini, L.; Puddu, M.; Ottonello, G.; Atzei, A.; et al. Metabolomics in newborns with intrauterine growth retardation (IUGR): Urine reveals markers of metabolic syndrome. J. Matern. Fetal Neonatal Med. 2011, 24, 36–40. [Google Scholar]
- Bell, M.J.; Ternberg, J.L.; Feigin, R.D.; Keating, J.P.; Marshall, R.; Barton, L.; Brotherton, T. Neonatal necrotizing enterocolitis. Therapeutic decisions based upon clinical staging. Ann. Surg. 1978, 187, 1–7. [Google Scholar] [CrossRef]
- International Committee for the Classification of Retinopathy of Prematurity. The international classification of retinopathy of prematurity revisited. Arch. Ophthalmol. 2005, 123, 991–999. [Google Scholar] [CrossRef]
- De Vries, L.S.; Eken, P.; Dubowitz, L.M.S. The spectrum of leukomalacia using cranial ultrasound. Behav. Brain Res. 1992, 49, 1–6. [Google Scholar] [CrossRef]
- Papile, L.A.; Burstein, J.; Burstein, R.; Koffler, H. Incidence and evolution of subependymal and intraventricular hemorrhage: A study of infants with birth weights less than 1500 gm. J. Pediatr. 1978, 92, 529–534. [Google Scholar] [CrossRef]
- Skjaerven, R.; Gjessing, H.K.; Bakketeig, L.S. Birthweight by gestational age in Norway. Acta Obstet. Gynecol. Scand. 2000, 79, 440–449. [Google Scholar] [CrossRef]
- Patel, A.L.; Engstrom, J.L.; Meier, P.P.; Kimura, R.E. Accuracy of methods for calculating postnatal growth velocity for extremely low birth weight infants. Pediatrics 2005, 116, 1466–1473. [Google Scholar] [CrossRef]
- Sachse, D.; Sletner, L.; Mørkrid, K.; Jenum, A.K.; Birkeland, K.I.; Rise, F.; Piehler, A.P.; Berg, J.P. Metabolic changes in urine during and after pregnancy in a large, multiethnic population-based cohort study of gestational diabetes. PLoS One 2012, 7, e52399. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S.; et al. HMDB: A knowledgebase for the human metabolome. Nucleic Acids Res. 2009, 37, D603–D610. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C. Systems biology: Metabonomics. Nature 2008, 455, 1054–1056. [Google Scholar] [CrossRef]
- Schlotterbeck, G.; Ross, A.; Dieterle, F.; Senn, H. Metabolic profiling technologies for biomarker discovery in biomedicine and drug development. Pharmacogenomics 2006, 7, 1055–1075. [Google Scholar] [CrossRef]
- Westerhuis, J.A.; Hoefsloot, H.C.J.; Smit, S.; Vis, D.J.; Smilde, A.K.; van Velzen, E.J.J.; van Duijnhoven, J.P.M.; van Dorsten, F.A. Assessment of PLSDA cross validation. Metabolomics 2008, 4, 81–89. [Google Scholar] [CrossRef]
- De Curtis, M.; Rigo, J. Nutrition and kidney in preterm infant. J. Matern. Fetal Neonatal Med. 2012, 25 (Suppl. 1), 55–59. [Google Scholar] [CrossRef]
- Foxall, P.J.; Bewley, S.; Neild, G.H.; Rodeck, C.H.; Nicholson, J.K. Analysis of fetal and neonatal urine using proton nuclear magnetic resonance spectroscopy. Arch. Dis. Child. Fetal Neonatal Ed. 1995, 73, F153–F157. [Google Scholar] [CrossRef]
- Greengard, O. Enzymic differentiation in mammalian tissues. Essays Biochem. 1971, 7, 159–205. [Google Scholar]
- Harvey, R.A.; Ferrier, D.R. Tricarboxylic Acid Cycle. In Lippincott’s Illustrated Reviews: Biochemistry, 5th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2010; pp. 109–116. [Google Scholar]
- Younoszai, M.K.; Andersen, D.W.; Filer, L.J., Jr.; Fomon, S.J. Urinary excretion of endogenous hydroxyproline by normal male infants. Pediatr. Res. 1967, 1, 266–270. [Google Scholar] [CrossRef]
- O’Sullivan, A.; He, X.; McNiven, E.M.; Hinde, K.; Haggarty, N.W.; Lonnerdal, B.; Slupsky, C.M. Metabolomic phenotyping validates the infant rhesus monkey as a model of human infant metabolism. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 355–363. [Google Scholar] [CrossRef]
- Diaz, S.O.; Pinto, J.; Graca, G.; Duarte, I.F.; Barros, A.S.; Galhano, E.; Pita, C.; Almeida, M.C.; Goodfellow, B.J.; Carreira, I.M.; et al. Metabolic biomarkers of prenatal disorders: An exploratory NMR metabonomics study of second trimester maternal urine and blood plasma. J. Proteome Res. 2011, 10, 3732–3742. [Google Scholar]
- McDonald, T.J.; Wu, G.; Nijland, M.J.; Jenkins, S.L.; Nathanielsz, P.W.; Jansson, T. Effect of 30% nutrient restriction in the first half of gestation on maternal and fetal baboon serum amino acid concentrations. Br. J. Nutr. 2013, 109, 1382–1388. [Google Scholar] [CrossRef]
- Cetin, I.; Corbetta, C.; Sereni, L.P.; Marconi, A.M.; Bozzetti, P.; Pardi, G.; Battaglia, F.C. Umbilical amino acid concentrations in normal and growth-retarded fetuses sampled in utero by cordocentesis. Am. J. Obstet. Gynecol. 1990, 162, 253–261. [Google Scholar] [CrossRef]
- Economides, D.L.; Nicolaides, K.H.; Gahl, W.A.; Bernardini, I.; Evans, M.I. Plasma amino acids in appropriate- and small-for-gestational-age fetuses. Am. J. Obstet. Gynecol. 1989, 161, 1219–1227. [Google Scholar] [CrossRef]
- Bonsante, F.; Iacobelli, S.; Chantegret, C.; Martin, D.; Gouyon, J.B. The effect of parenteral nitrogen and energy intake on electrolyte balance in the preterm infant. Eur. J. Clin. Nutr. 2011, 65, 1088–1093. [Google Scholar] [CrossRef]
- Bonsante, F.; Iacobelli, S.; Latorre, G.; Rigo, J.; de Felice, C.; Robillard, P.Y.; Gouyon, J.B. Initial amino acid intake influences phosphorus and calcium homeostasis in preterm infants—It is time to change the composition of the early parenteral nutrition. PLoS One 2013, 8, e72880. [Google Scholar]
- Elstgeest, L.E.; Martens, S.E.; Lopriore, E.; Walther, F.J.; te Pas, A.B. Does parenteral nutrition influence electrolyte and fluid balance in preterm infants in the first days after birth? PLoS One 2010, 5, e9033. [Google Scholar]
- Ichikawa, G.; Watabe, Y.; Suzumura, H.; Sairenchi, T.; Muto, T.; Arisaka, O. Hypophosphatemia in small for gestational age extremely low birth weight infants receiving parenteral nutrition in the first week after birth. J. Pediatr. Endocrinol. Metab. 2012, 25, 317–321. [Google Scholar]
- Mizumoto, H.; Mikami, M.; Oda, H.; Hata, D. Refeeding syndrome in a small-for-dates micro-preemie receiving early parenteral nutrition. Pediatr. Int. 2012, 54, 715–717. [Google Scholar] [CrossRef]
- Rigo, J.; Marlowe, M.L.; Bonnot, D.; Senterre, T.; Lapillonne, A.; Kermorvant-Duchemin, E.; Hascoet, J.M.; Desandes, R.; Malfilatre, G.; Pladys, P.; et al. Benefits of a new pediatric triple-chamber bag for parenteral nutrition in preterm infants. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 210–217. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moltu, S.J.; Sachse, D.; Blakstad, E.W.; Strømmen, K.; Nakstad, B.; Almaas, A.N.; Westerberg, A.C.; Rønnestad, A.; Brække, K.; Veierød, M.B.; et al. Urinary Metabolite Profiles in Premature Infants Show Early Postnatal Metabolic Adaptation and Maturation. Nutrients 2014, 6, 1913-1930. https://doi.org/10.3390/nu6051913
Moltu SJ, Sachse D, Blakstad EW, Strømmen K, Nakstad B, Almaas AN, Westerberg AC, Rønnestad A, Brække K, Veierød MB, et al. Urinary Metabolite Profiles in Premature Infants Show Early Postnatal Metabolic Adaptation and Maturation. Nutrients. 2014; 6(5):1913-1930. https://doi.org/10.3390/nu6051913
Chicago/Turabian StyleMoltu, Sissel J., Daniel Sachse, Elin W. Blakstad, Kenneth Strømmen, Britt Nakstad, Astrid N. Almaas, Ane C. Westerberg, Arild Rønnestad, Kristin Brække, Marit B. Veierød, and et al. 2014. "Urinary Metabolite Profiles in Premature Infants Show Early Postnatal Metabolic Adaptation and Maturation" Nutrients 6, no. 5: 1913-1930. https://doi.org/10.3390/nu6051913