DNA Hypermethylation of the Serotonin Receptor Type-2A Gene Is Associated with a Worse Response to a Weight Loss Intervention in Subjects with Metabolic Syndrome

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Study Protocol
2.2. Anthropometry and Blood Pressure
2.3. Biochemical Analysis
2.4. Psychological Assessment
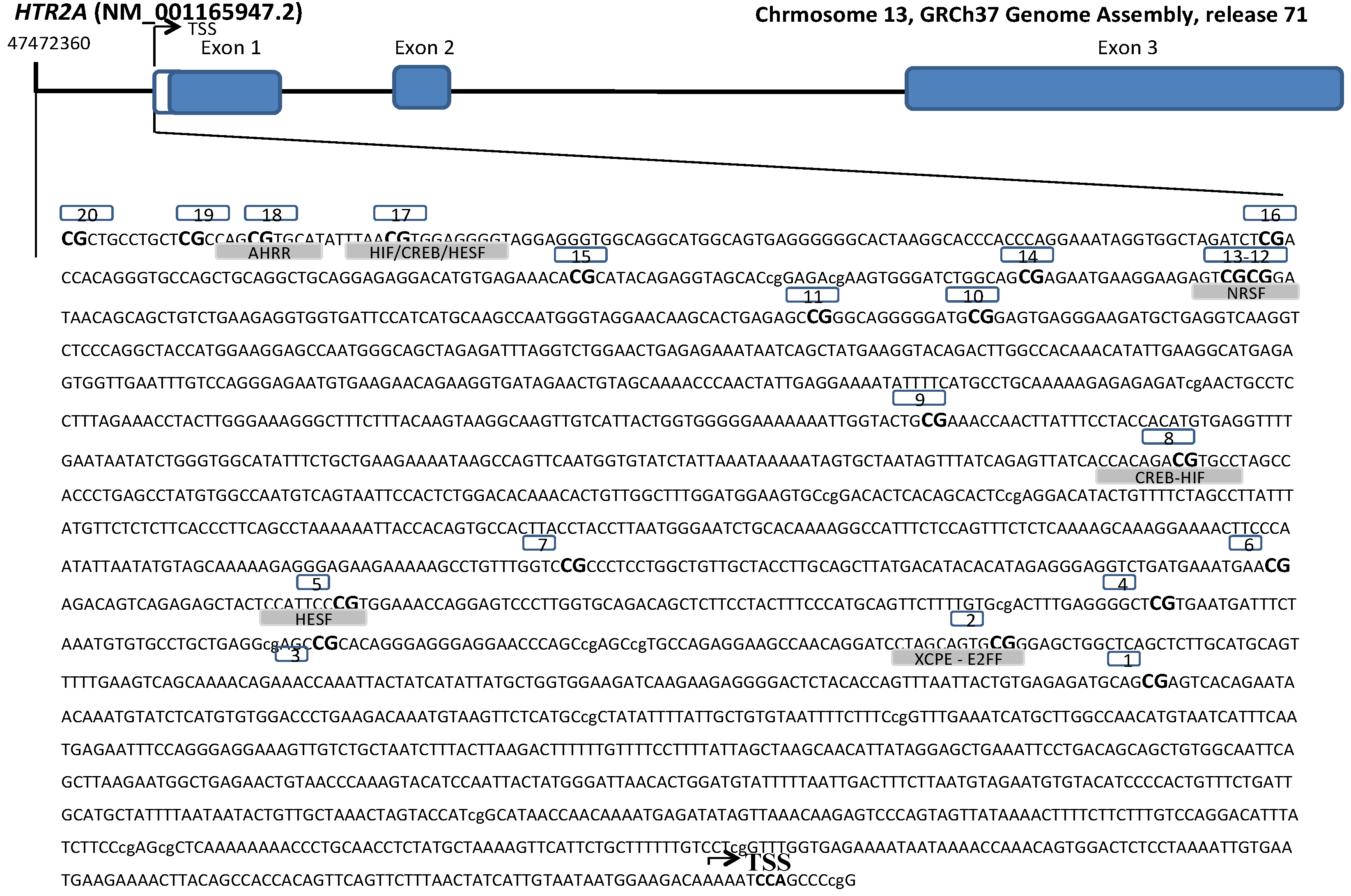
2.5. DNA Isolation and DNA Methylation Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CpG ID 1 | Illumina ID | CHR Position 2 | Reference 3 |
|---|---|---|---|
| 1 | cg15894389 | 13:47470857 | c.-688 |
| 2 | cg02250787 | 13:47470989 | c.-820 |
| 3 | cg06476131 | 13:47471052 | c.-883 |
| 4 | cg16188532 | 13:47471090 | c.-921 |
| 5 | cg09361691 | 13:47471169 | c.-1000 |
| 6 | cg11514288 | 13:47471197 | c.-1028 |
| 7 | cg27068143 | 13:47471264 | c.-1095 |
| 8 | cg10323433 | 13:47471562 | c.-1393 |
| 9 | cg02027079 | 13:47471705 | c.-1536 |
| 10 | cg01192538 | 13:47472050 | c.-1881 |
| 11 | cg01620540 | 13:47472064 | c.-1895 |
| 12 | cg06020661 | 13:47472138 | c.-1969 |
| 13 | cg09798090 | 13:47472140 | c.-1971 |
| 14 | cg24320398 | 13:47472158 | c.-1989 |
| 15 | cg18200810 | 13:47472200 | c.-2031 |
| 16 | cg15692052 | 13:47472250 | c.-2081 |
| 17 | cg24118521 | 13:47472330 | c.-2161 |
| 18 | cg23881368 | 13:47472343 | c.-2174 |
| 19 | cg05506829 | 13:47472349 | c.-2180 |
| 20 | cg07075299 | 13:47472360 | c.-2191 |
2.6. Analysis of Gene Expression by Quantitative Real-Time PCR
2.7. In Silico Sequence Analysis
2.8. Statistical Analyses
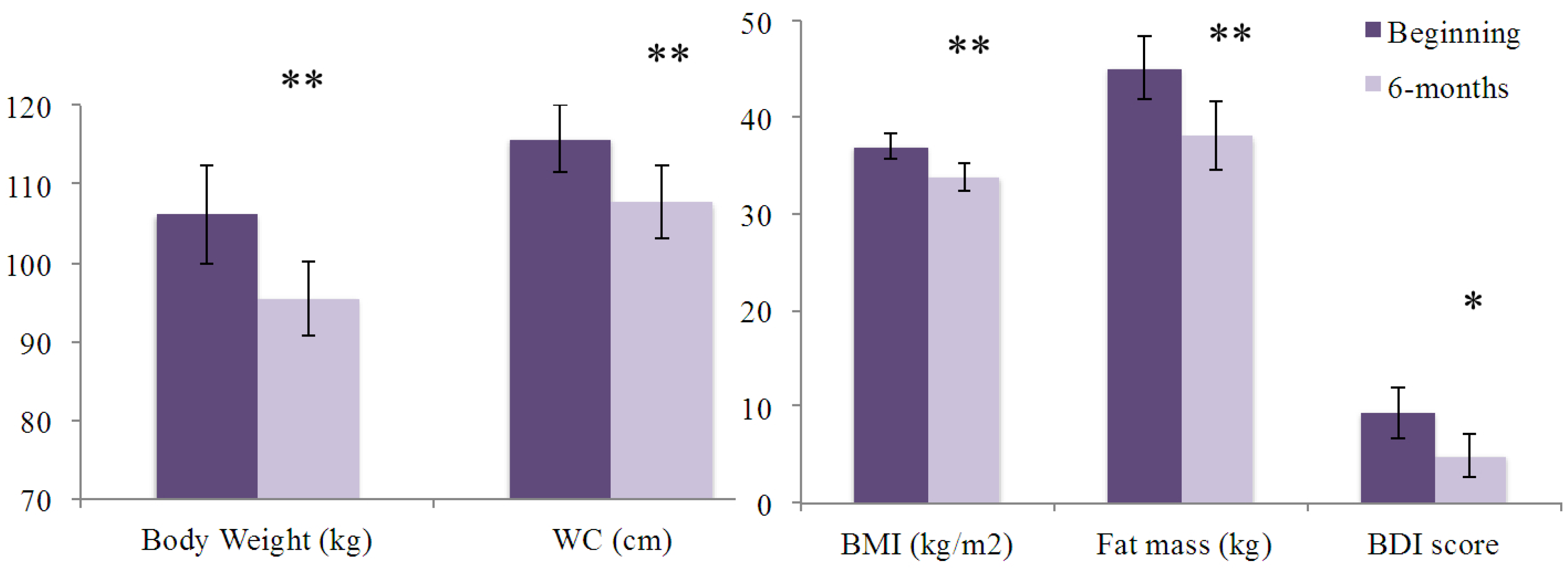
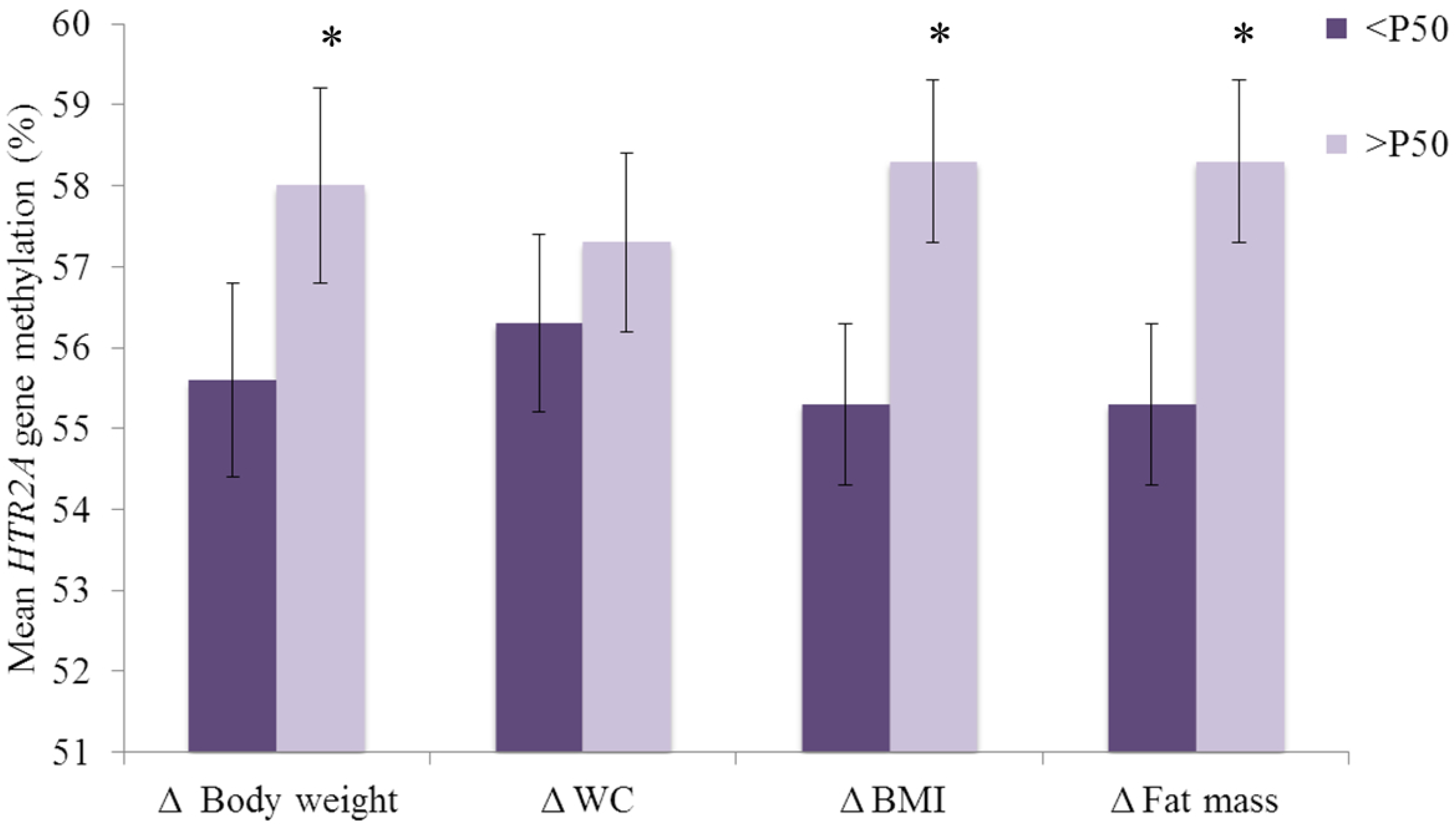
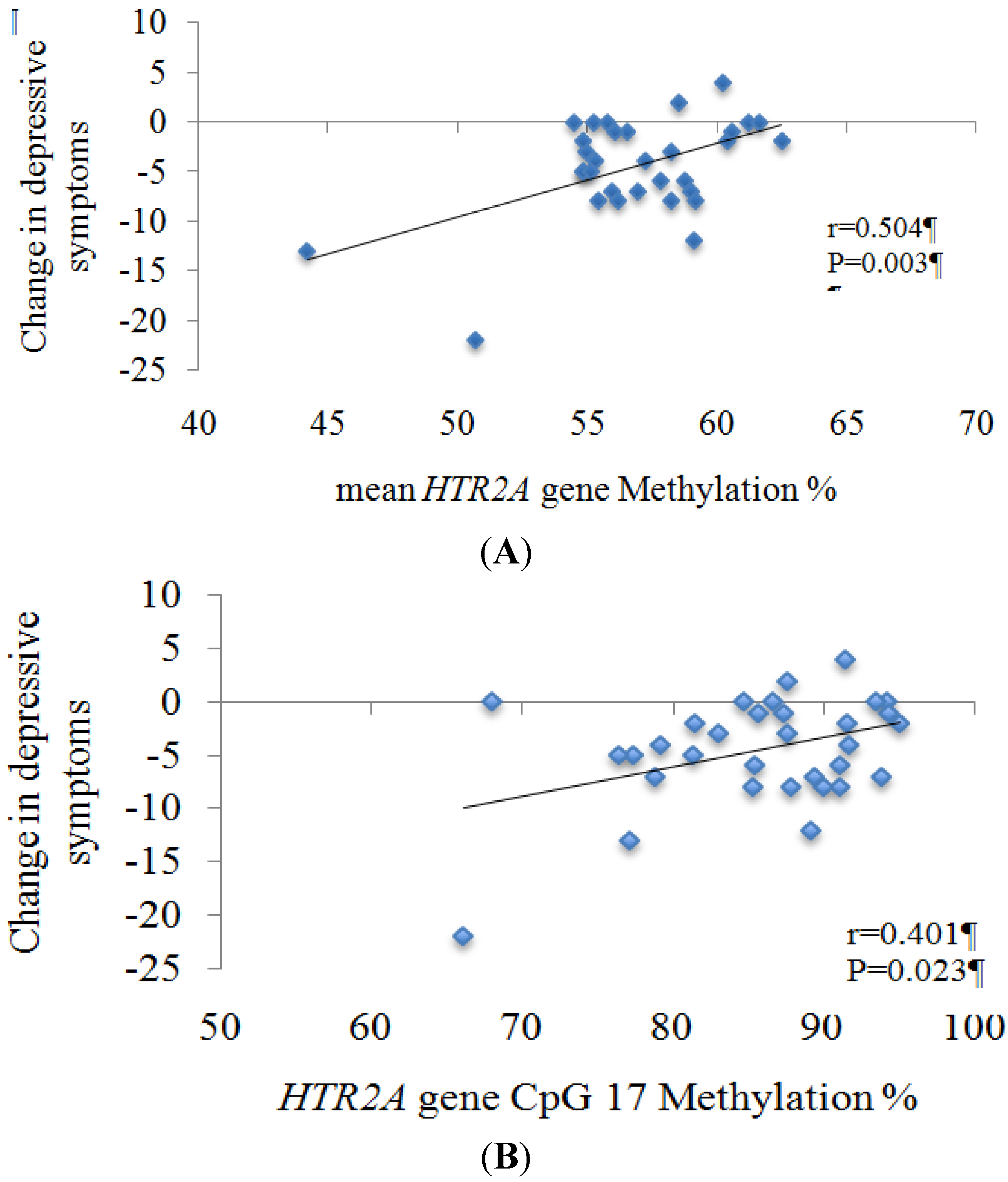
3. Results
| Variables | Mean HTR2A Gene Methylation % | p Trend | ||
|---|---|---|---|---|
| Low (n = 14) | Medium (n = 14) | High (n = 13) | ||
| Body weight (kg) | 97.6 ± 4.6 | 106.4 ± 4.6 | 110.7 ± 4.8 | 0.059 |
| BMI (kg/m2) | 37.0 ± 1.0 | 37.0 ± 1.0 | 37.0 ± 1.0 | 0.995 |
| Waist circumference (cm) | 110.8 ± 3.0 | 114.0 ± 3.0 | 120.5 ± 3.1 | 0.027 |
| Total fat mass (kg) | 43.8 ± 2.4 | 45.9 ± 2.4 | 45.4 ± 2.5 | 0.682 |
| Truncal fat mass (kg) | 26.1 ± 1.8 | 27.6 ± 1.8 | 26.2 ± 1.9 | 0.993 |
| SBP (mmHg) | 150.3 ± 4.8 | 151.2 ± 4.8 | 148.1 ± 4.9 | 0.733 |
| DBP (mmHg) | 85.4 ± 2.2 | 87.3 ± 2.2 | 84.8 ± 2.3 | 0.784 |
| Glucose (mg/dL) | 9.8 ± 1.5 | 7.4 ± 1.5 | 11.1 ± 1.6 | 0.498 |
| Insulin (μU/mL) | 11.0 ± 2.3 | 18.1 ± 2.3 | 21.7 ± 2.4 | 0.004 |
| TC (mg/dL) | 225.1 ± 12.4 | 211.1 ± 12.9 | 205.1 ± 12.9 | 0.280 |
| Triglycerides (mg/dL) | 166.8 ± 29.9 | 224.6 ± 31.1 | 247.3 ± 31.1 | 0.079 |
| NEFA (nmol/L) | 0.58 ± 0.05 | 0.50 ± 0.05 | 0.56 ± 0.05 | 0.705 |
| BDI score | 9.8 ± 1.9 | 8.3 ± 1.9 | 10.9 ± 1.9 | 0.636 |


| Δ Body Weight | Δ WC | Δ BMI | Δ Fat Mass (kg) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Baseline | CpG Sites | r | Uncorrected p-Value | r | Uncorrected p-Value | r | Uncorrected p-Value | r | Uncorrected p-Value |
| cg15894389 | 1 | 0.110 | 0.537 | −0.117 | 0.510 | 0.127 | 0.474 | 0.167 | 0.346 |
| cg02250787 | 2 | −0.063 | 0.725 | −0.083 | 0.639 | −0.063 | 0.723 | 0.008 | 0.963 |
| cg06476131 | 3 | 0.284 | 0.103 | 0.080 | 0.652 | 0.286 | 0.100 | 0.381 | 0.026 * |
| cg16188532 | 4 | 0.379 | 0.027 * | 0.124 | 0.483 | 0.383 | 0.025 * | 0.415 | 0.015 * |
| cg09361691 | 5 | −0.195 | 0.268 | −0.188 | 0.287 | −0.198 | 0.261 | −0.153 | 0.386 |
| cg11514288 | 6 | −0.143 | 0.418 | −0.154 | 0.383 | −0.144 | 0.414 | −0.145 | 0.412 |
| cg27068143 | 7 | 0.369 | 0.032 * | 0.116 | 0.513 | 0.397 | 0.020 * | 0.382 | 0.025 * |
| cg10323433 | 8 | 0.292 | 0.094 | 0.052 | 0.769 | 0.350 | 0.042 | 0.249 | 0.155 |
| cg02027079 | 9 | 0.011 | 0.951 | 0.160 | 0.365 | 0.009 | 0.957 | 0.042 | 0.815 |
| cg01192538 | 10 | 0.225 | 0.201 | 0.193 | 0.274 | 0.230 | 0.190 | 0.214 | 0.225 |
| cg01620540 | 11 | −0.070 | 0.694 | −0.251 | 0.152 | −0.014 | 0.937 | −0.073 | 0.680 |
| cg06020661 | 12 | 0.177 | 0.316 | 0.206 | 0.243 | 0.196 | 0.267 | 0.196 | 0.267 |
| cg09798090 | 13 | −0.042 | 0.814 | −0.054 | 0.762 | −0.037 | 0.837 | −0.059 | 0.740 |
| cg24320398 | 14 | 0.338 | 0.051 | 0.266 | 0.128 | 0.377 | 0.028 * | 0.343 | 0.047 |
| cg18200810 | 15 | 0.310 | 0.075 | 0.222 | 0.206 | 0.360 | 0.037 * | 0.322 | 0.063 |
| cg15692052 | 16 | 0.091 | 0.608 | −0.025 | 0.890 | 0.121 | 0.494 | 0.103 | 0.563 |
| cg24118521 | 17 | 0.342 | 0.047 | 0.280 | 0.108 | 0.381 | 0.026 * | 0.414 | 0.015 * |
| cg23881368 | 18 | 0.297 | 0.087 | 0.269 | 0.124 | 0.319 | 0.066 | 0.291 | 0.095 |
| cg05506829 | 19 | 0.280 | 0.108 | 0.252 | 0.151 | 0.297 | 0.088 | 0.268 | 0.125 |
| cg07075299 | 20 | 0.296 | 0.089 | 0.250 | 0.153 | 0.307 | 0.078 | 0.301 | 0.084 |

4. Discussion
5. Conclusions
Supplementary Files
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Richards, D. Prevalence and clinical course of depression: A review. Clin. Psychol. Rev. 2011, 31, 1117–1125. [Google Scholar] [CrossRef]
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the international diabetes federation task force on epidemiology and prevention; national heart, lung, and blood institute; american heart association; world heart federation; international atherosclerosis society; and international association for the study of obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef]
- Lichtman, J.H.; Froelicher, E.S.; Blumenthal, J.A.; Carney, R.M.; Doering, L.V.; Frasure-Smith, N.; Freedland, K.E.; Jaffe, A.S.; Leifheit-Limson, E.C.; Sheps, D.S.; et al. Depression as a risk factor for poor prognosis among patients with acute coronary syndrome: Systematic review and recommendations: A scientific statement from the american heart association. Circulation 2014, 129, 1350–1369. [Google Scholar] [CrossRef]
- Pan, A.; Keum, N.; Okereke, O.I.; Sun, Q.; Kivimaki, M.; Rubin, R.R.; Hu, F.B. Bidirectional association between depression and metabolic syndrome: A systematic review and meta-analysis of epidemiological studies. Diabetes Care 2012, 35, 1171–1180. [Google Scholar]
- Bondy, B. Common genetic factors for depression and cardiovascular disease. Dialogues Clin. Neurosci. 2007, 9, 19–28. [Google Scholar]
- Campion, J.; Milagro, F.I.; Martinez, J.A. Individuality and epigenetics in obesity. Obes. Rev. 2009, 10, 383–392. [Google Scholar]
- Schroeder, M.; Krebs, M.O.; Bleich, S.; Frieling, H. Epigenetics and depression: Current challenges and new therapeutic options. Curr. Opin. Psychiatry 2010, 23, 588–592. [Google Scholar] [CrossRef]
- Philibert, R.A.; Sandhu, H.; Hollenbeck, N.; Gunter, T.; Adams, W.; Madan, A. The relationship of 5HTT (SLC6A4) methylation and genotype on mrna expression and liability to major depression and alcohol dependence in subjects from the iowa adoption studies. Am. J. Med. Genet. B 2008, 147, 543–549. [Google Scholar]
- De la Iglesia-González, R.; Mansego-Talavera, M.; Sánchez-Muniz, F.J.; Zulet-Alzórriz, M.; Martinez-Hernández, J. Arylesterase activity is associated with antioxidant intake and paraoxonase 1 (PON1) gene methylation in metabolic syndrome patients following an energy restricted diet. EXCLI J. 2014, 13, 416–426. [Google Scholar]
- Ordovas, J.M.; Smith, C.E. Epigenetics and cardiovascular disease. Nat. Rev. Cardiol. 2010, 7, 510–519. [Google Scholar] [CrossRef]
- Terry, M.B.; Delgado-Cruzata, L.; Vin-Raviv, N.; Wu, H.C.; Santella, R.M. DNA methylation in white blood cells: Association with risk factors in epidemiologic studies. Epigenetics 2011, 6, 828–837. [Google Scholar] [CrossRef]
- Moleres, A.; Campion, J.; Milagro, F.I.; Marcos, A.; Campoy, C.; Garagorri, J.M.; Gomez-Martinez, S.; Martinez, J.A.; Azcona-Sanjulian, M.C.; Marti, A. Differential DNA methylation patterns between high and low responders to a weight loss intervention in overweight or obese adolescents: The evasyon study. FASEB J. 2013, 27, 2504–2512. [Google Scholar] [CrossRef]
- Berger, M.; Gray, J.A.; Roth, B.L. The expanded biology of serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef]
- Stunes, A.K.; Reseland, J.E.; Hauso, O.; Kidd, M.; Tommeras, K.; Waldum, H.L.; Syversen, U.; Gustafsson, B.I. Adipocytes express a functional system for serotonin synthesis, reuptake and receptor activation. Diabetes Obes. Metab. 2011, 13, 551–558. [Google Scholar]
- Fidalgo, S.; Ivanov, D.K.; Wood, S.H. Serotonin: From top to bottom. Biogerontology 2013, 14, 21–45. [Google Scholar] [CrossRef]
- Fabbri, C.; Marsano, A.; Serretti, A. Genetics of serotonin receptors and depression: State of the art. Curr. Drug Targets 2013, 14, 531–548. [Google Scholar] [CrossRef]
- Li, P.; Tiwari, H.K.; Lin, W.Y.; Allison, D.B.; Chung, W.K.; Leibel, R.L.; Yi, N.; Liu, N. Genetic association analysis of 30 genes related to obesity in a european american population. Int. J. Obes. 2014, 38, 724–729. [Google Scholar]
- Falkenberg, V.R.; Gurbaxani, B.M.; Unger, E.R.; Rajeevan, M.S. Functional genomics of serotonin receptor 2A (HTR2A): Interaction of polymorphism, methylation, expression and disease association. Neuromolecular Med. 2011, 13, 66–76. [Google Scholar] [CrossRef]
- Polesskaya, O.O.; Aston, C.; Sokolov, B.P. Allele c-specific methylation of the 5-HT2A receptor gene: Evidence for correlation with its expression and expression of DNA methylase DNMT1. J. Neurosci. Res. 2006, 83, 362–373. [Google Scholar] [CrossRef]
- Ghadirivasfi, M.; Nohesara, S.; Ahmadkhaniha, H.R.; Eskandari, M.R.; Mostafavi, S.; Thiagalingam, S.; Abdolmaleky, H.M. Hypomethylation of the serotonin receptor type-2A gene (HTR2A) at T102C polymorphic site in DNA derived from the saliva of patients with schizophrenia and bipolar disorder. Am. J. Med. Genet. B 2011, 156, 536–545. [Google Scholar] [CrossRef]
- Abdolmaleky, H.M.; Yaqubi, S.; Papageorgis, P.; Lambert, A.W.; Ozturk, S.; Sivaraman, V.; Thiagalingam, S. Epigenetic dysregulation of HTR2A in the brain of patients with schizophrenia and bipolar disorder. Schizophr. Res. 2011, 129, 183–190. [Google Scholar] [CrossRef]
- Perez-Cornago, A.; Zulet, M.; Martinez, J. Association between mood and diet quality in subjects with metabolic syndrome participating in a behavioural weight-loss program: A cross-sectional assessment. Nutr. Neurosci. 2014. [Google Scholar] [CrossRef]
- Perez-Cornago, A.; Lopez-Legarrea, P.; de la Iglesia, R.; Lahortiga, F.; Martinez, J.A.; Zulet, M.A. Longitudinal relationship of diet and oxidative stress with depressive symptoms in patients with metabolic syndrome after following a weight loss treatment: The RESMENA project. Clin. Nutr. 2013. [Google Scholar] [CrossRef]
- Perez-Cornago, A.; de la Iglesia, R.; Lopez-Legarrea, P.; Abete, I.; Navas-Carretero, S.; Lacunza, C.I.; Lahortiga, F.; Martinez-Gonzalez, M.A.; Martinez, J.A.; Zulet, M.A. A decline in inflammation is associated with less depressive symptoms after a dietary intervention in metabolic syndrome patients: A longitudinal study. Nutr. J. 2014, 13, 36. [Google Scholar] [CrossRef]
- Perez-Cornago, A.; Ramírez, M.; Zulet, M.; Martinez, J.A. Effect of dietary restriction on peripheral monoamines and anxiety symptoms in obese subjects with metabolic syndrome. Psychoneuroendocrinology 2014, 47, 98–106. [Google Scholar] [CrossRef]
- Zulet, M.A.; Bondia-Pons, I.; Abete, I.; de la Iglesia, R.; Lopez-Legarrea, P.; Forga, L.; Navas-Carretero, S.; Martinez, J.A. The reduction of the metabolyc syndrome in navarra-spain (RESMENA-S) study: A multidisciplinary strategy based on chrononutrition and nutritional education, together with dietetic and psychological control. Nutr. Hosp. 2011, 26, 16–26. [Google Scholar]
- Whitworth, J.A.; Chalmers, J. World health organisation-international society of hypertension (WHO/ISH) hypertension guidelines. Clin. Exp. Hypertens. 2004, 26, 747–752. [Google Scholar] [CrossRef]
- Hermsdorff, H.H.; Mansego, M.L.; Campion, J.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A. TNF-alpha promoter methylation in peripheral white blood cells: Relationship with circulating tnfalpha, truncal fat and n-6 pufa intake in young women. Cytokine 2013, 64, 265–271. [Google Scholar] [CrossRef]
- Lopez-Legarrea, P.; Mansego, M.L.; Zulet, M.A.; Martinez, J.A. Serpine1, PAI-1 protein coding gene, methylation levels and epigenetic relationships with adiposity changes in obese subjects with metabolic syndrome features under dietary restriction. J. Clin. Biochem. Nutr. 2013, 53, 139–144. [Google Scholar] [CrossRef]
- Conde, C.; Useros, E. Adaptación castellana de la escala de evaluación conductual para la depresión de beck. Rev. Psiqiat Psicol. Med. 1975, 12, 217–236. [Google Scholar]
- Touleimat, N.; Tost, J. Complete pipeline for Infinium(®) Human Methylation 450K BeadChip data processing using subset quantile normalization for accurate DNA methylation estimation. Epigenomics 2012, 4, 325–341. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- National Center of Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 12 January 2014).
- Quandt, K.; Frech, K.; Karas, H.; Wingender, E.; Werner, T. Matind and matinspector: New fast and versatile tools for detection of consensus matches in nucleotide sequence data. Nucleic Acids Res. 1995, 23, 4878–4884. [Google Scholar] [CrossRef]
- Jiang, X.; Tian, F.; Du, Y.; Copeland, N.G.; Jenkins, N.A.; Tessarollo, L.; Wu, X.; Pan, H.; Hu, X.Z.; Xu, K.; et al. BHLHB2 controls bdnf promoter 4 activity and neuronal excitability. J. Neurosci. 2008, 28, 1118–1130. [Google Scholar] [CrossRef]
- Rosenberger, P.; Schwab, J.M.; Mirakaj, V.; Masekowsky, E.; Mager, A.; Morote-Garcia, J.C.; Unertl, K.; Eltzschig, H.K. Hypoxia-inducible factor-dependent induction of netrin-1 dampens inflammation caused by hypoxia. Nat. Immunol. 2009, 10, 195–202. [Google Scholar] [CrossRef]
- Urdampilleta, A.; Gonzalez-Muniesa, P.; Portillo, M.P.; Martinez, J.A. Usefulness of combining intermittent hypoxia and physical exercise in the treatment of obesity. J. Physiol. Biochem. 2012, 68, 289–304. [Google Scholar] [CrossRef]
- Milagro, F.I.; Mansego, M.L.; de Miguel, C.; Martinez, J.A. Dietary factors, epigenetic modifications and obesity outcomes: Progresses and perspectives. Mol. Asp. Med. 2013, 34, 782–812. [Google Scholar] [CrossRef]
- Belzeaux, R.; Formisano-Treziny, C.; Loundou, A.; Boyer, L.; Gabert, J.; Samuelian, J.C.; Feron, F.; Naudin, J.; Ibrahim, E.C. Clinical variations modulate patterns of gene expression and define blood biomarkers in major depression. J. Psychiatr. Res. 2010, 44, 1205–1213. [Google Scholar] [CrossRef]
- Abdolmaleky, H.M.; Smith, C.L.; Faraone, S.V.; Shafa, R.; Stone, W.; Glatt, S.J.; Tsuang, M.T. Methylomics in psychiatry: Modulation of gene-environment interactions may be through DNA methylation. Am. J. Med. Genet B 2004, 127B, 51–59. [Google Scholar] [CrossRef]
- Cordero, P.; Gomez-Uriz, A.M.; Campion, J.; Milagro, F.I.; Martinez, J.A. Dietary supplementation with methyl donors reduces fatty liver and modifies the fatty acid synthase DNA methylation profile in rats fed an obesogenic diet. Genes Nutr. 2013, 8, 105–113. [Google Scholar] [CrossRef]
- Kring, S.I.; Werge, T.; Holst, C.; Toubro, S.; Astrup, A.; Hansen, T.; Pedersen, O.; Sorensen, T.I. Polymorphisms of serotonin receptor 2A and 2C genes and comt in relation to obesity and type 2 diabetes. PLoS One 2009, 4, e6696. [Google Scholar]
- Cao, J.; Liu, X.; Han, S.; Zhang, C.K.; Liu, Z.; Li, D. Association of the HTR2A gene with alcohol and heroin abuse. Hum. Genet 2014, 133, 357–365. [Google Scholar] [CrossRef]
- Sorli, J.V.; Frances, F.; Gonzalez, J.I.; Guillen, M.; Portoles, O.; Sabater, A.; Coltell, O.; Corella, D. Impact of the −1438 G > A polymorphism in the serotonin 2A receptor gene on anthropometric profile and obesity risk: A case-control study in a Spanish Mediterranean population. Appetite 2008, 50, 260–265. [Google Scholar]
- Mandelli, L.; Serretti, A. Gene environment interaction studies in depression and suicidal behavior: An update. Neurosci. Biobehav. Rev. 2013, 37, 2375–2397. [Google Scholar] [CrossRef]
- Garbett, K.; Gal-Chis, R.; Gaszner, G.; Lewis, D.A.; Mirnics, K. Transcriptome alterations in the prefrontal cortex of subjects with schizophrenia who committed suicide. Neuropsychopharmacol Hung 2008, 10, 9–14. [Google Scholar]
- Hurlemann, R.; Matusch, A.; Kuhn, K.U.; Berning, J.; Elmenhorst, D.; Winz, O.; Kolsch, H.; Zilles, K.; Wagner, M.; Maier, W.; et al. 5-HT2A receptor density is decreased in the at-risk mental state. Psychopharmacology 2008, 195, 579–590. [Google Scholar]
- Sigurdh, J.; Allard, P.; Spigset, O.; Hagglof, B. Platelet serotonin transporter and 5-HT2A receptor binding in adolescents with eating disorders. Int. J. Neurosci. 2013, 123, 333–338. [Google Scholar] [CrossRef]
- Nonogaki, K.; Nozue, K.; Oka, Y. Increased hypothalamic 5-HT2A receptor gene expression and effects of pharmacologic 5-HT2A receptor inactivation in obese Ay mice. Biochem. Biophys. Res. Commun. 2006, 351, 1078–1082. [Google Scholar] [CrossRef]
- Shelton, R.C.; Sanders-Bush, E.; Manier, D.H.; Lewis, D.A. Elevated 5-HT2A receptors in postmortem prefrontal cortex in major depression is associated with reduced activity of protein kinase A. Neuroscience 2009, 158, 1406–1415. [Google Scholar] [CrossRef]
- Smith, R.A.; Levine, T.R.; Lachlan, K.A.; Fediuk, T.A. The high cost of complexity in experimental design and data analysis: Type I and type II error rates in multiway anova. Hum. Commun. Res. 2002, 28, 515–530. [Google Scholar]
- Keppel, G.; Wickens, T. Design and Analysis: A Researcher’s Handbook, 4th ed.; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Cohen, J.; Cohen, P.; West, S.; Aiken, L. Applied Multiple Regression/Correlation Analysis for the Behavioral Sciences, 3rd ed.; Routledge: Mahwah, NJ, USA, 2002. [Google Scholar]
- Milagro, F.I.; Martinez, J.A. Epigenetics of obesity and weight loss. Endocrinol. Nutr. 2013, 60, 12–14. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Perez-Cornago, A.; Mansego, M.L.; Zulet, M.A.; Martinez, J.A. DNA Hypermethylation of the Serotonin Receptor Type-2A Gene Is Associated with a Worse Response to a Weight Loss Intervention in Subjects with Metabolic Syndrome. Nutrients 2014, 6, 2387-2403. https://doi.org/10.3390/nu6062387
Perez-Cornago A, Mansego ML, Zulet MA, Martinez JA. DNA Hypermethylation of the Serotonin Receptor Type-2A Gene Is Associated with a Worse Response to a Weight Loss Intervention in Subjects with Metabolic Syndrome. Nutrients. 2014; 6(6):2387-2403. https://doi.org/10.3390/nu6062387
Chicago/Turabian StylePerez-Cornago, Aurora, Maria L. Mansego, María Angeles Zulet, and José Alfredo Martinez. 2014. "DNA Hypermethylation of the Serotonin Receptor Type-2A Gene Is Associated with a Worse Response to a Weight Loss Intervention in Subjects with Metabolic Syndrome" Nutrients 6, no. 6: 2387-2403. https://doi.org/10.3390/nu6062387
APA StylePerez-Cornago, A., Mansego, M. L., Zulet, M. A., & Martinez, J. A. (2014). DNA Hypermethylation of the Serotonin Receptor Type-2A Gene Is Associated with a Worse Response to a Weight Loss Intervention in Subjects with Metabolic Syndrome. Nutrients, 6(6), 2387-2403. https://doi.org/10.3390/nu6062387