Leucine Promotes Proliferation and Differentiation of Primary Preterm Rat Satellite Cells in Part through mTORC1 Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Animals
2.2. Reagents
2.3. Primary Culture of Preterm Rat Satellite Cells
2.4. Differentiation Culture
2.5. Immunocytochemical
2.6. Cell Proliferation Rate Assay
2.7. Western Blot
2.8. Statistical Analysis
3. Results
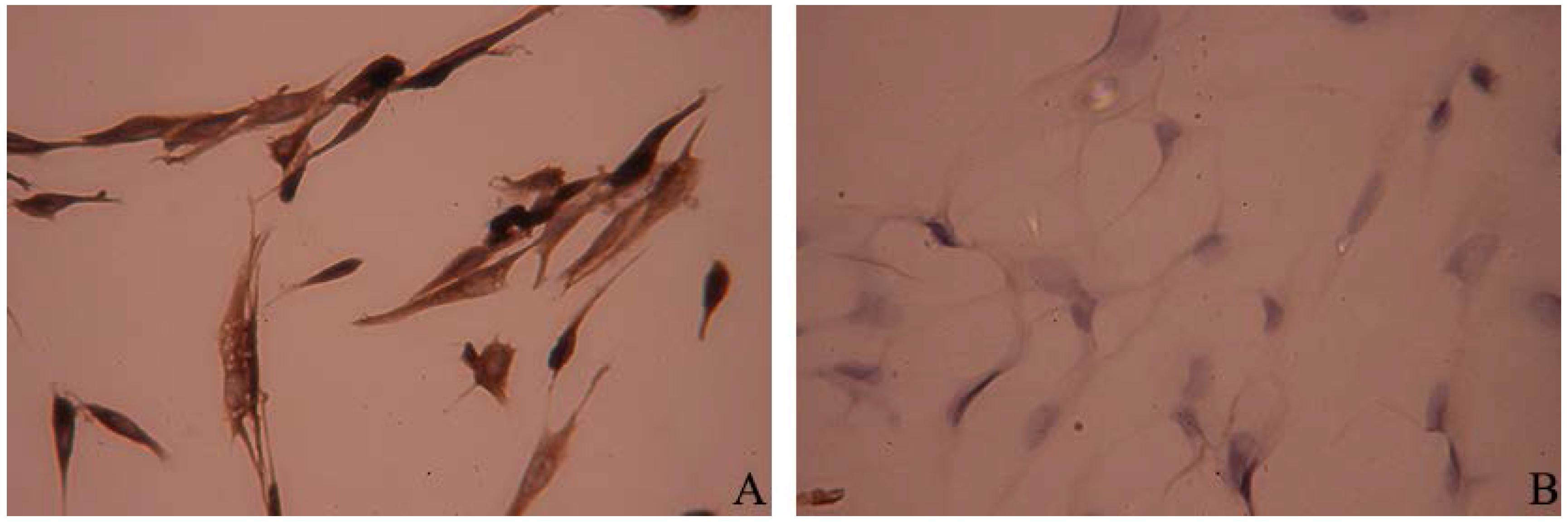
3.1. Culture and Identification of Primary Preterm Rat Satellite Cells
3.2. Leucine Promotes Primary Satellite Cells Proliferation Through mTORC1 Pathway



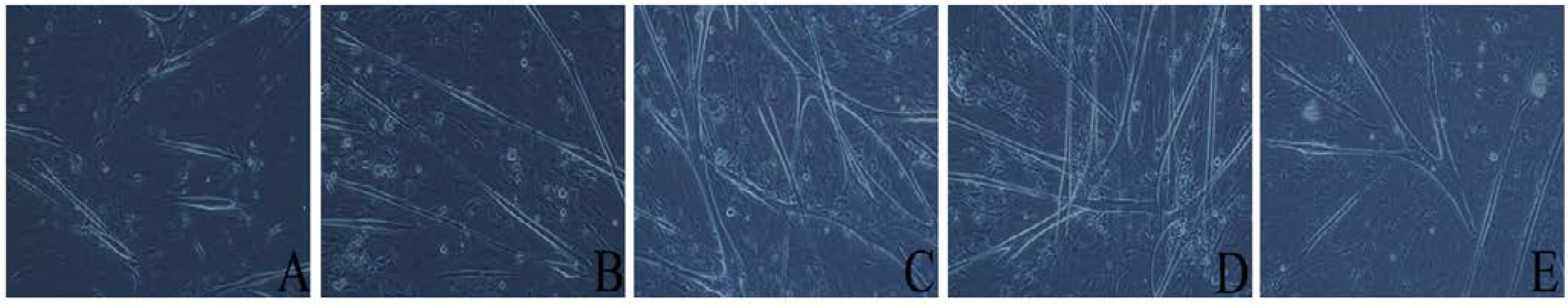
3.3. Leucine Promotes Differentiation of Primary Preterm Rat Satellite Cells

3.4. Involvement of mTORC1 in Leucine-Stimulated Differentiation of Primary Satellite Cells

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Simmer, K.; Rakshasbhuvankar, A.; Deshpande, G. Standardised parenteral nutrition. Nutrients 2013, 5, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.H.; Thomas, P.; Peabody, J. Extrauterine growth restriction remains a serious problem in prematurely born neonates. Pediatrics 2003, 111, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Ehrenkranz, R.A.; Dusick, A.M.; Vohr, B.R.; Wright, L.L.; Wrage, L.A.; Poole, W.K. Growth in the neonatal intensive care unit influences neurodevelopmental and growth outcomes of extremely low birth weight infants. Pediatrics 2006, 117, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Fenton, T.R.; Premji, S.S.; Al-Wassia, H.; Sauve, R.S. Higher versus lower protein intake in formula-fed low birth weight infants. Cochrane Database Syst. Rev. 2014, 4, D3959. [Google Scholar]
- Yu, M.X.; Shen, Z.Y.; Qiu, X.S.; Mo, Q.P. High-protein diets alters body composition and improves insulin resistance in a rat model of low birth weight. J. Investig. Med. 2012, 60, 1174–1179. [Google Scholar] [PubMed]
- Klein, C.J. Nutrient requirements for preterm infant formulas. J. Nutr. 2002, 132, 1395S–1577S. [Google Scholar] [PubMed]
- Suryawan, A.; Davis, T.A. Regulation of protein synthesis by amino acids in muscle of neonates. Front Biosci. 2011, 16, 1445–1460. [Google Scholar] [CrossRef]
- Drummond, M.J.; Dreyer, H.C.; Fry, C.S.; Glynn, E.L.; Rasmussen, B.B. Nutritional and contractile regulation of human skeletal muscle protein synthesis and mTORC1 signaling. J. Appl. Physiol. 2009, 106, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Kido, T.; Yoshizawa, F.; Ito, Y.; Nishizawa, N. Rapid suppression of protein degradation in skeletal muscle after oral feeding of leucine in rats. J. Nutr. Biochem. 2002, 13, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Sarbassov, D.D.; Ali, S.M.; Sabatini, D.M. Growing roles for the mTOR pathway. Curr. Opin. Cell Biol. 2005, 17, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.M.; Blenis, J. Molecular mechanisms of mTOR-mediated translational control. Nat. Rev. Mol. Cell Biol. 2009, 10, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Sancak, Y.; Sabatini, D.M. Rag proteins regulate amino-acid-induced mTORC1 signalling. Biochem. Soc. Trans. 2009, 37, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimball, S.R.; Jefferson, L.S. Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. J. Nutr. 2006, 136, 227S–231S. [Google Scholar] [PubMed]
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal muscle mass and distribution in 468 men and women aged 18–88 yr. J. Appl. Physiol. 2000, 89, 81–88. [Google Scholar] [PubMed]
- Brown, L.D. Endocrine regulation of fetal skeletal muscle growth: Impact on future metabolic health. J. Endocrinol. 2014, 221, R13–R29. [Google Scholar] [CrossRef] [PubMed]
- Relaix, F.; Zammit, P.S. Satellite cells are essential for skeletal muscle regeneration: The cell on the edge returns centre stage. Development 2012, 139, 2845–2856. [Google Scholar] [CrossRef] [PubMed]
- Le Grand, F.; Rudnicki, M.A. Skeletal muscle satellite cells and adult myogenesis. Curr. Opin. Cell Biol. 2007, 19, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012. [Google Scholar] [CrossRef]
- Ishibashi, J.; Perry, R.L.; Asakura, A.; Rudnicki, M.A. MyoD induces myogenic differentiation through cooperation of its NH2- and COOH-terminal regions. J. Cell Biol. 2005, 171, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Yablonka-Reuveni, Z.; Rudnicki, M.A.; Rivera, A.J.; Primig, M.; Anderson, J.E.; Natanson, P. The transition from proliferation to differentiation is delayed in satellite cells from mice lacking MyoD. Dev. Biol. 1999, 210, 440–455. [Google Scholar] [CrossRef] [PubMed]
- Cornelison, D.D.; Olwin, B.B.; Rudnicki, M.A.; Wold, B.J. MyoD(−/−) satellite cells in single-fiber culture are differentiation defective and MRF4 deficient. Dev. Biol. 2000, 224, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Kumar, R.M.; Penn, B.H.; Berkes, C.A.; Kooperberg, C.; Boyer, L.A.; Young, R.A.; Tapscott, S.J. Global and gene-specific analyses show distinct roles for Myod and Myog at a common set of promoters. Embo J. 2006, 25, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Myer, A.; Olson, E.N.; Klein, W.H. MyoD cannot compensate for the absence of myogenin during skeletal muscle differentiation in murine embryonic stem cells. Dev. Biol. 2001, 229, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Erbay, E.; Chen, J. The mammalian target of rapamycin regulates C2C12 myogenesis via a kinase-independent mechanism. J. Biol. Chem. 2001, 276, 36079–36082. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ge, Y.; Drnevich, J.; Zhao, Y.; Band, M.; Chen, J. Mammalian target of rapamycin regulates miRNA-1 and follistatin in skeletal myogenesis. J. Cell Biol. 2010, 189, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.M.; Wang, L.F.; Cheng, K.T.; Hsu, H.H.; Gau, B.; Su, B. Effects of Anoectochilus formosanus Hayata extract and glucocorticoid on lung maturation in preterm rats. Phytomedicine 2004, 11, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Chang, L.; Li, W.; Liu, H. Effect of amygdalin on the proliferation of hyperoxia-exposed type II alveolar epithelial cells isolated from premature rat. J. Huazhong Univ. Sci. Technol. Med. Sci. 2004, 24, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, S.; Li, W.; Du, H.; Zhu, W. Isolation and characterization of primary skeletal muscle satellite cells from rats. Toxicol. Mech. Methods 2012, 22, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Anthony, T.G.; Kimball, S.R.; Jefferson, L.S. Signaling pathways involved in translational control of protein synthesis in skeletal muscle by leucine. J. Nutr. 2001, 131, 856S–860S. [Google Scholar] [PubMed]
- Averous, J.; Gabillard, J.C.; Seiliez, I.; Dardevet, D. Leucine limitation regulates myf5 and myoD expression and inhibits myoblast differentiation. Exp. Cell Res. 2012, 318, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Huang, Z.; Chen, D.; Yang, T.; Liu, G. MicroRNA-27a is induced by leucine and contributes to leucine-induced proliferation promotion in C2C12 cells. Int. J. Mol Sci. 2013, 14, 14076–14084. [Google Scholar] [CrossRef] [PubMed]
- Jastrzebski, K.; Hannan, K.M.; Tchoubrieva, E.B.; Hannan, R.D.; Pearson, R.B. Coordinate regulation of ribosome biogenesis and function by the ribosomal protein S6 kinase, a key mediator of mTOR function. Growth Fact. 2007, 25, 209–226. [Google Scholar] [CrossRef]
- Gibbons, J.J.; Abraham, R.T.; Yu, K. Mammalian target of rapamycin: discovery of rapamycin reveals a signaling pathway important for normal and cancer cell growth. Semin. Oncol. 2009, 36, S3–S17. [Google Scholar] [CrossRef] [PubMed]
- Varma, S.; Khandelwal, R.L. Effects of rapamycin on cell proliferation and phosphorylation of mTOR and p70(S6K) in HepG2 and HepG2 cells overexpressing constitutively active Akt/PKB. Biochim. Biophys. Acta 2007, 1770, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Chen, J. Mammalian target of rapamycin (mTOR) signaling network in skeletal myogenesis. J. Biol. Chem. 2012, 287, 43928–43935. [Google Scholar] [CrossRef] [PubMed]
- Coolican, S.A.; Samuel, D.S.; Ewton, D.Z.; McWade, F.J.; Florini, J.R. The mitogenic and myogenic actions of insulin-like growth factors utilize distinct signaling pathways. J. Biol. Chem. 1997, 272, 6653–6662. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, J.-M.; Yu, M.-X.; Shen, Z.-Y.; Guo, C.-Y.; Zhuang, S.-Q.; Qiu, X.-S. Leucine Promotes Proliferation and Differentiation of Primary Preterm Rat Satellite Cells in Part through mTORC1 Signaling Pathway. Nutrients 2015, 7, 3387-3400. https://doi.org/10.3390/nu7053387
Dai J-M, Yu M-X, Shen Z-Y, Guo C-Y, Zhuang S-Q, Qiu X-S. Leucine Promotes Proliferation and Differentiation of Primary Preterm Rat Satellite Cells in Part through mTORC1 Signaling Pathway. Nutrients. 2015; 7(5):3387-3400. https://doi.org/10.3390/nu7053387
Chicago/Turabian StyleDai, Jie-Min, Mu-Xue Yu, Zhen-Yu Shen, Chu-Yi Guo, Si-Qi Zhuang, and Xiao-Shan Qiu. 2015. "Leucine Promotes Proliferation and Differentiation of Primary Preterm Rat Satellite Cells in Part through mTORC1 Signaling Pathway" Nutrients 7, no. 5: 3387-3400. https://doi.org/10.3390/nu7053387