Apple Polyphenols Decrease Atherosclerosis and Hepatic Steatosis in ApoE−/− Mice through the ROS/MAPK/NF-κB Pathway

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Assessment of Aortic Atherosclerosis and Hepatic Histology
2.3. Assessment of Hepatic Redox Status
2.4. Measurement of Body Weight, Hepatic Lipids and Metabolic Parameters
2.5. Cell Isolation and Adhesion Assay
2.6. Cell Viability Assay and ROS Detection
2.7. Quantitation of Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mRNA | Forward Primer | Reverse Primer |
|---|---|---|
| rat ICAM-1 | AGATCATACGGGTTTGGGCTTC | TATGACTCGTGAAAGAAATCAGCTC |
| rat VCAM-1 | TGTGGAAGTGT GCCCGAAAT | TGCCTTGCGGATGGTGTAC |
| rat CCL2 | ATGCAGGTCTCTGTCACGCT | GGTGCTGAAGTCCTTAGGGT |
| rat GAPDH | CATGTTCGTCATGGGTGTGAACCA | ATGGCATGGACTGTGGTCATGAGT |
| mouse PPARα | TCAGGGTACCACTACGGAGTTCA | CCGAATAGTTCGCCGAAAGA |
| mouse SCAP | ACTGGACTGAAGGCAGGTCAA | GCCTCTAGTCTAGGTCCAAAGAGTTG |
| mouse SCD-1 | CAGTGCCGCGCATCTCT | CCCGGGATTGAATGTTCTTG |
| mouse HMG-CoA R | CCCAGTTGTGCGTCTTCCA | TTCGAGCCAGGCTTTCACTT |
| mouse HMG-CoA S | GCCGTGAACTGGGTCGAA | GCATATAGCAATGTCCTGCAA |
| mouse Nrf2 | GGCCCAGCATATCCAGACA | TGTGGGCAACCTGGGAGTAG |
| mouse GCLm | ACATTGAAGCCCAGGATTGG | CTCTTCACGATGACCGAGTACCT |
| mouse SOD-1 | A GCCCGGCGGATGAAG | CCTTTCCAGCAGTCACATTGC |
| mouse GAPDH | ATGTTTGTGATGGGTGTGAACCAG | TAGCCATATTCATTGTCATACCAGG |
2.8. Enzyme-Linked Immunosorbent Assay
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
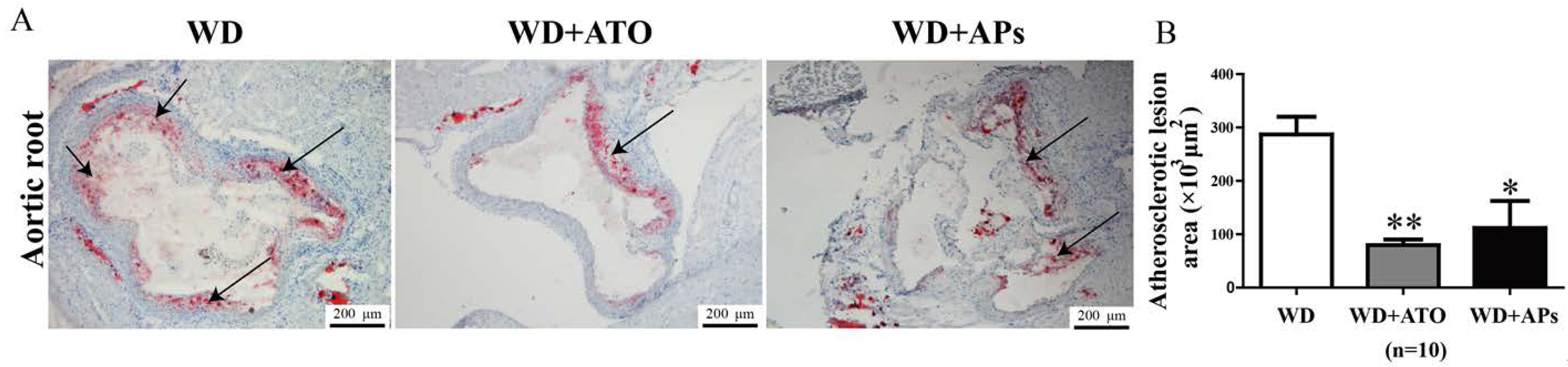
3.1. APs Prevent Atherosclerotic Plaque Development

3.2. APs Normalize the Body Weight and Metabolic Parameters in the Apo −/− Mice Fed with the Western-Type Diet

3.3. APs Ameliorate Liver Lipogenesis and Hepatic Steatosis in WD-fed ApoE−/− Mice

3.4. APs Prevent the Liver from Hepatic Oxidative Stress

3.5. APs Attenuate CD68 Expression in the Aortic Root, and CCL2 and VCAM-1 Levels in Plasma

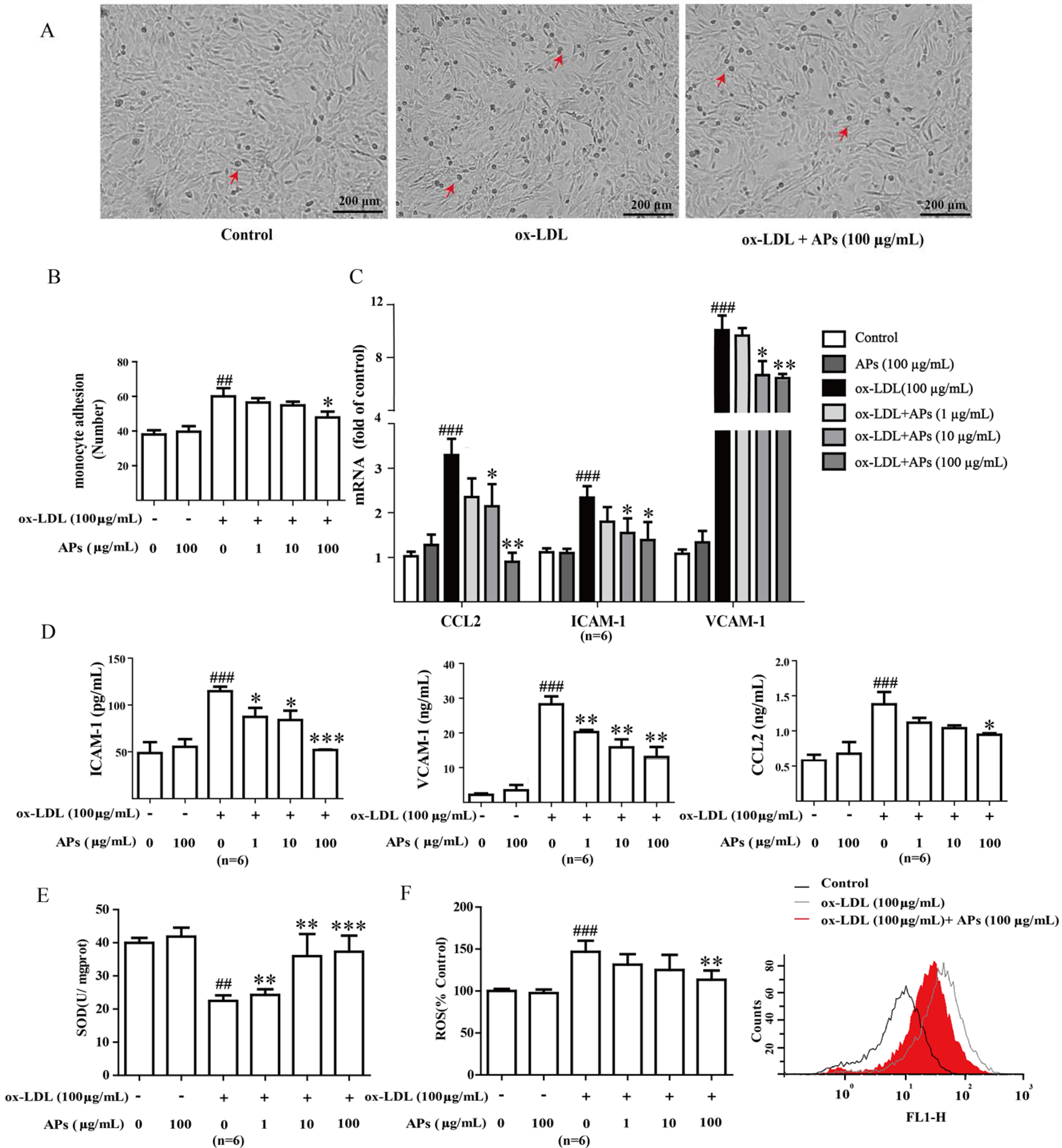
3.6. APs Suppress ox-LDL-Induced Pro-Inflammatory Factors and Alleviates Oxidant Stress in ox-LDL-Treated Endothelial Cells
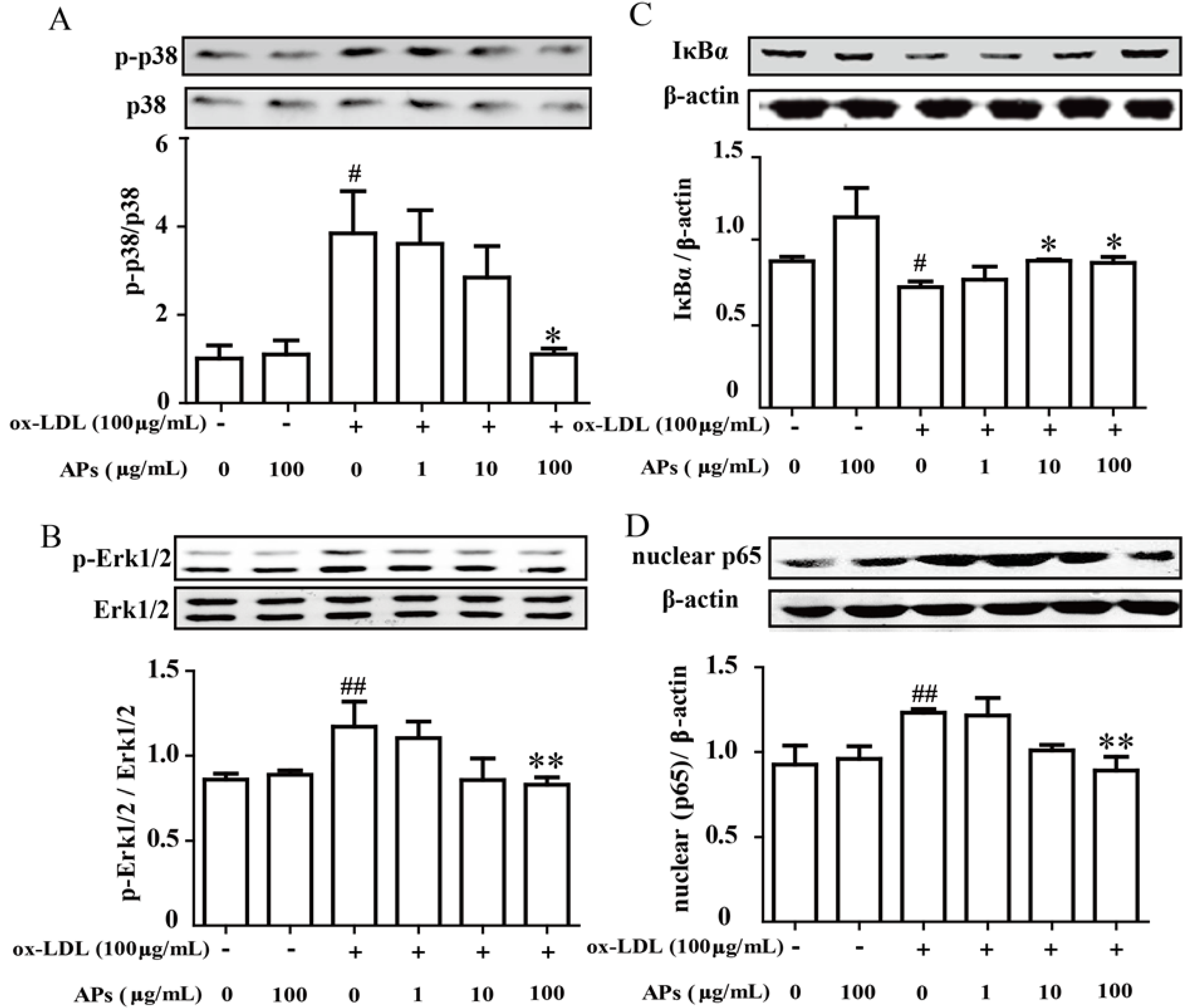
3.7. APs inhibit the ox-LDL-Induced MAPK and NF-κB Pathway Activation in Endothelial Cells


4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ApoE−/− mice | Apolipoprotein E knockout mice |
| ATO | atorvastatin |
| CAT | catalase |
| CCL-2 | The chemokine (C-C motif) ligand 2 |
| CVD | Cardio vascular disorder |
| DAPI | 4′,6-diamidino-2-phenylindole |
| Erk | extracellular regulated protein kinases |
| Fe/Asc | iron-ascorbate |
| GCLm | glutamate-cysteine ligase modifier subunit |
| GPx | glutathione peroxidase |
| GPX-1 | GSH peroxidase |
| GSH-Px | glutathione peroxidase |
| HDL | high-density lipoprotein |
| HDL-c | high-density lipoprotein cholesterol |
| H&E | hematoxylin and eosin |
| HMG-CoA R | HMG-CoA reductase |
| HMG-CoA S | HMG-CoA Synthase |
| ICAM-1 | intracellular adhesion molecule-1 |
| IκB-α | nuclear factor of kappa-light-polypeptide gene enhancer in B-cells inhibitor, alpha |
| IL-1β | Interleukine-1 beta |
| LDL-c | high-density lipoprotein cholesterol |
| LPS | lipopolysaccharide |
| MAPKs | mitogen-activated protein kinases |
| MDA | malondialdehyde |
| MTT | 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide |
| NASH | nonalcoholic steatohepatitis |
| NF-κB | nuclear factor kappa B |
| Nrf2 | nuclear factor erythroid-2 |
| ox-LDL | oxidized low-density lipoprotein |
| PPARα | peroxisome proliferator-activated receptor-α |
| RAECs | rat aortic endothelial cells |
| ROS | reactive oxygen species |
| RPBMNCs | rat peripheral blood mononuclear cells |
| SCAP | SREBP cleavage-activating protein |
| SCD-1 | stearoyl-CoA desaturase-1 |
| SOD | superoxide dismutase |
| SREBP-2 | sterol regulatory element-binding protein-1c |
| TNF-α | tumor necrosis factor-a |
| VCAM-1 | vascular adhesion molecule-1 |
| WD | Western-type diet |
References
- Koutsos, A.; Lovegrove, J.A. An apple a day keeps the doctor away: Inter-relationship between apple consumption, the gut microbiota and cardiometabolic disease risk reduction. In Diet-Microbe Interactions in the Gut: Effects on Human Health and Disease; Tuohy, K., del Rio, D., Eds.; Elsevier: London, UK, 2015; pp. 173–194. [Google Scholar]
- Lotito, S.B.; Frei, B. Relevance of apple polyphenols as antioxidants in human plasma: Contrasting in vitro and in vivo effects. Free Radic. Biol. Med. 2004, 36, 201–211. [Google Scholar] [PubMed]
- Dauchet, L.; Amouyel, P.; Dallongeville, J. Fruits, vegetables and coronary heart disease. Nat. Rev. Cardiol. 2009, 6, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, M.P.; McKay, D.L.; Blumberg, J.B. Flavonoid basics: Chemistry, sources, mechanisms of action, and safety. J. Nutr. Gerontol. Geriatr. 2012, 31, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knekt, P.; Kumpulainen, J.; Jarvinen, R.; Rissanen, H.; Heliovaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [PubMed]
- Shoji, T.; Akazome, Y.; Kanda, T.; Ikeda, M. The toxicology and safety of apple polyphenol extract. Food Chem. Toxicol. 2004, 42, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Ammirati, E.; Moroni, F.; Norata, G.D.; Magnoni, M.; Camici, P.G. Markers of inflammation associated with plaque progression and instability in patients with carotid atherosclerosis. Mediators. Inflamm. 2015, 2015, 718329. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Spagnoli, V.; Tardif, J.C.; L’Allier, P.L. Novel anti-inflammatory therapies for the treatment of atherosclerosis. Atherosclerosis 2015, 240, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Chew, G.T.; Watts, G.F. New peroxisome proliferator-activated receptor agonists: Potential treatments for atherogenic dyslipidemia and non-alcoholic fatty liver disease. Expert Opin. Pharmacother. 2014, 15, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Tiong, A.Y.; Brieger, D. Inflammation and coronary artery disease. Am. Heart J. 2005, 150, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Kim, B.H.; Seo, H.S.; Lee, Y.J.; Kim, H.H.; Son, H.H.; Choi, M.H. Cholesterol-induced non-alcoholic fatty liver disease and atherosclerosis aggravated by systemic inflammation. PLoS ONE 2014, 9, e97841. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Williams, K.J.; Borén, J. Subendothelial lipoprotein retention as the initiating process in atherosclerosis: Update and therapeutic implications. Circulation 2007, 116, 1832–1844. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Z.; Kar, N.S.; Podrez, E.A. Oxidized phospholipids: Biomarker for cardiovascular diseases. Int. J. Biochem. Cell Biol. 2009, 41, 1241–1244. [Google Scholar] [CrossRef] [PubMed]
- Wouters, K.; van Gorp, P.J.; Bieghs, V.; Gijbels, M.J.; Duimel, H.; Lütjohann, D.; Kerksiek, A.; van Kruchten, R.; Maeda, N.; Staels, B.; et al. Dietary cholesterol, rather than liver steatosis, leads to hepatic inflammation in hyperlipidemic mouse models of nonalcoholic steatohepatitis. Hepatology 2008, 48, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Hermida, N.; Balligand, J.L. Low-density lipoprotein-cholesterol-induced endothelial dysfunction and oxidative stress: The role of statins. Antioxid. Redox Signal. 2014, 20, 1216–1237. [Google Scholar] [CrossRef] [PubMed]
- Pober, J.S.; Sessa, W.C. Evolving functions of endothelial cells in inflammation. Nat. Rev. Immunol. 2007, 7, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Stoll, G.; Bendszus, M. Inflammation and atherosclerosis: Novel insights into plaque formation and destabilization. Stroke 2006, 37, 1923–1932. [Google Scholar] [CrossRef] [PubMed]
- Bobryshev, Y.V. Monocyte recruitment and foam cell formation in atherosclerosis. Micron 2006, 37, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.A. Evaluation of anti-atherosclerotic activities of ppar-alpha, ppar-gamma, and lxr agonists in hyperlipidemic atherosclerosis-susceptible f(1)b hamsters. Atherosclerosis 2011, 214, 86–93. [Google Scholar] [CrossRef] [PubMed]
- O’Byrne, D.; Devaraj, S.; Islam, K.N.; Collazo, R.; McDonald, L.; Grundy, S.; Jialal, I. Low-density lipoprotein (LDL)-induced monocyte-endothelial cell adhesion, soluble cell adhesion molecules, and autoantibodies to oxidized-LDL in chronic renal failure patients on dialysis therapy. Metabolism 2001, 50, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Auclair, S.; Milenkovic, D.; Besson, C.; Chauvet, S.; Gueux, E.; Morand, C.; Mazur, A.; Scalbert, A. Catechin reduces atherosclerotic lesion development in apo E-deficient mice: A transcriptomic study. Atherosclerosis 2009, 204, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.B.; Sun, F.; Nie, X.L.; Wang, J. Leptin and coronary heart disease: A systematic review and meta-analysis. Atherosclerosis 2014, 233, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Hampton, R.; Dimster-Denk, D.; Rine, J. The biology of HMG-CoA reductase: The pros of contra-regulation. Trends. Biochem. Sci. 1996, 21, 140–145. [Google Scholar] [CrossRef]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2015, 62, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Steinbrenner, H. Interference of selenium and selenoproteins with the insulin-regulated carbohydrate and lipid metabolism. Free Radic. Biol. Med. 2013, 65, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Polimeni, L.; del Ben, M.; Baratta, F.; Perri, L.; Albanese, F.; Pastori, D.; Violi, F.; Angelico, F. Oxidative stress: New insights on the association of non-alcoholic fatty liver disease and atherosclerosis. World J. Hepatol. 2015, 7, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Jiang, Y.F.; Ponnusamy, M.; Diallo, M. Role of Nrf2 in chronic liver disease. World J. Gastroenterol. 2014, 20, 13079–13087. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.R.; Ren, Y.L.; Zhu, J.J.; Hu, Y.J.; Zheng, J.S.; Fan, H.; Xu, Y.; Wang, G.; Liu, W.X. Resveratrol increases nephrin and podocin expression and alleviates renal damage in rats fed a high-fat diet. Nutrients 2014, 6, 2619–2631. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.A. Nonpharmacological approaches for reducing serum low-density lipoprotein cholesterol. Curr. Opin. Cardiol. 2014, 29, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Stöger, J.L.; Gijbels, M.J.; van der Velden, S.; Manca, M.; van der Loos, C.M.; Biessen, E.A.; Daemen, M.J.; Lutgens, E.; de Winther, M.P. Distribution of macrophage polarization markers in human atherosclerosis. Atherosclerosis 2012, 225, 461–468. [Google Scholar] [CrossRef] [PubMed]
- McLaren, J.E.; Michael, D.R.; Ashlin, T.G.; Ramji, D.P. Cytokines, macrophage lipid metabolism and foam cells: Implications for cardiovascular disease therapy. Prog. Lipid Res. 2011, 50, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, A.L.; Subramanian, S.S.; Cury, R.C.; Truong, Q.A.; Gardecki, J.A.; Tearney, G.J.; Hoffmann, U.; Brady, T.J.; Tawakol, A. Distribution of inflammation within carotid atherosclerotic plaques with high-risk morphological features: A comparison between positron emission tomography activity, plaque morphology, and histopathology. Circ. Cardiovasc. Imaging 2012, 5, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Xu, Y.; Chen, L.; Fang, Q.; Yan, X. Atorvastatin inhibits CD68 expression in aortic root through a GRP78-involved pathway. Cardiovasc. Drugs Ther. 2014, 28, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Kakkar, V.; Lu, X. Impact of MCP-1 in atherosclerosis. Curr. Pharm. Des. 2014, 20, 4580–4588. [Google Scholar] [CrossRef] [PubMed]
- Rolin, J.; Maghazachi, A.A. Implications of chemokines, chemokine receptors, and inflammatory lipids in atherosclerosis. J. Leukoc. Biol. 2014, 95, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, P.N. Molecular biology of atherosclerosis. Physiol. Rev. 2013, 93, 1317–1542. [Google Scholar] [CrossRef] [PubMed]
- Asare, Y.; Shagdarsuren, E.; Schmid, J.A.; Tilstam, P.V.; Grommes, J.; El Bounkari, O.; Schütz, A.K.; Weber, C.; de Winther, M.P.; Noels, H.; et al. Endothelial CSN5 impairs NF-κB activation and monocyte adhesion to endothelial cells and is highly expressed in human atherosclerotic lesions. Thromb. Haemost. 2013, 110, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.X.; Zhang, S.J.; Xiong, Y.K.; Jia, Y.L.; Sun, Y.H.; Lin, X.X.; Shen, H.J.; Xie, Q.M.; Yan, X.F. EETs Attenuate Ox-LDL-Induced LTB4 Production and Activity by Inhibiting p38 MAPK Phosphorylation and 5-LO/BLT1 Receptor Expression in Rat Pulmonary Arterial Endothelial Cells. PLoS ONE 2015, 10, e0128278. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.X.; Zhang, S.J.; Liu, Y.N.; Lin, X.X.; Sun, Y.H.; Shen, H.J.; Yan, X.F.; Xie, Q.M. EETs alleviate ox-LDL-induced inflammation by inhibiting LOX-1 receptor expression in rat pulmonary arterial endothelial cells. Eur. J. Pharmacol. 2014, 727, 43–51. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.-R.; Li, J.-Y.; Dong, X.-W.; Tan, Z.-J.; Wu, W.-Z.; Xie, Q.-M.; Yang, Y.-M. Apple Polyphenols Decrease Atherosclerosis and Hepatic Steatosis in ApoE−/− Mice through the ROS/MAPK/NF-κB Pathway. Nutrients 2015, 7, 7085-7105. https://doi.org/10.3390/nu7085324
Xu Z-R, Li J-Y, Dong X-W, Tan Z-J, Wu W-Z, Xie Q-M, Yang Y-M. Apple Polyphenols Decrease Atherosclerosis and Hepatic Steatosis in ApoE−/− Mice through the ROS/MAPK/NF-κB Pathway. Nutrients. 2015; 7(8):7085-7105. https://doi.org/10.3390/nu7085324
Chicago/Turabian StyleXu, Zhe-Rong, Jin-You Li, Xin-Wei Dong, Zhong-Ju Tan, Wei-Zhen Wu, Qiang-Min Xie, and Yun-Mei Yang. 2015. "Apple Polyphenols Decrease Atherosclerosis and Hepatic Steatosis in ApoE−/− Mice through the ROS/MAPK/NF-κB Pathway" Nutrients 7, no. 8: 7085-7105. https://doi.org/10.3390/nu7085324
APA StyleXu, Z.-R., Li, J.-Y., Dong, X.-W., Tan, Z.-J., Wu, W.-Z., Xie, Q.-M., & Yang, Y.-M. (2015). Apple Polyphenols Decrease Atherosclerosis and Hepatic Steatosis in ApoE−/− Mice through the ROS/MAPK/NF-κB Pathway. Nutrients, 7(8), 7085-7105. https://doi.org/10.3390/nu7085324