Excess Folic Acid Increases Lipid Storage, Weight Gain, and Adipose Tissue Inflammation in High Fat Diet-Fed Rats

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Histological Analysis of Adipose and Liver Samples
2.3. Glucose Tolerance Tests
2.4. Plasma Measurements
2.5. Culture of 3T3L1 Adipocytes
2.6. Analytical Procedures
2.7. mRNA Quantification
2.8. Statistical Analysis
3. Results
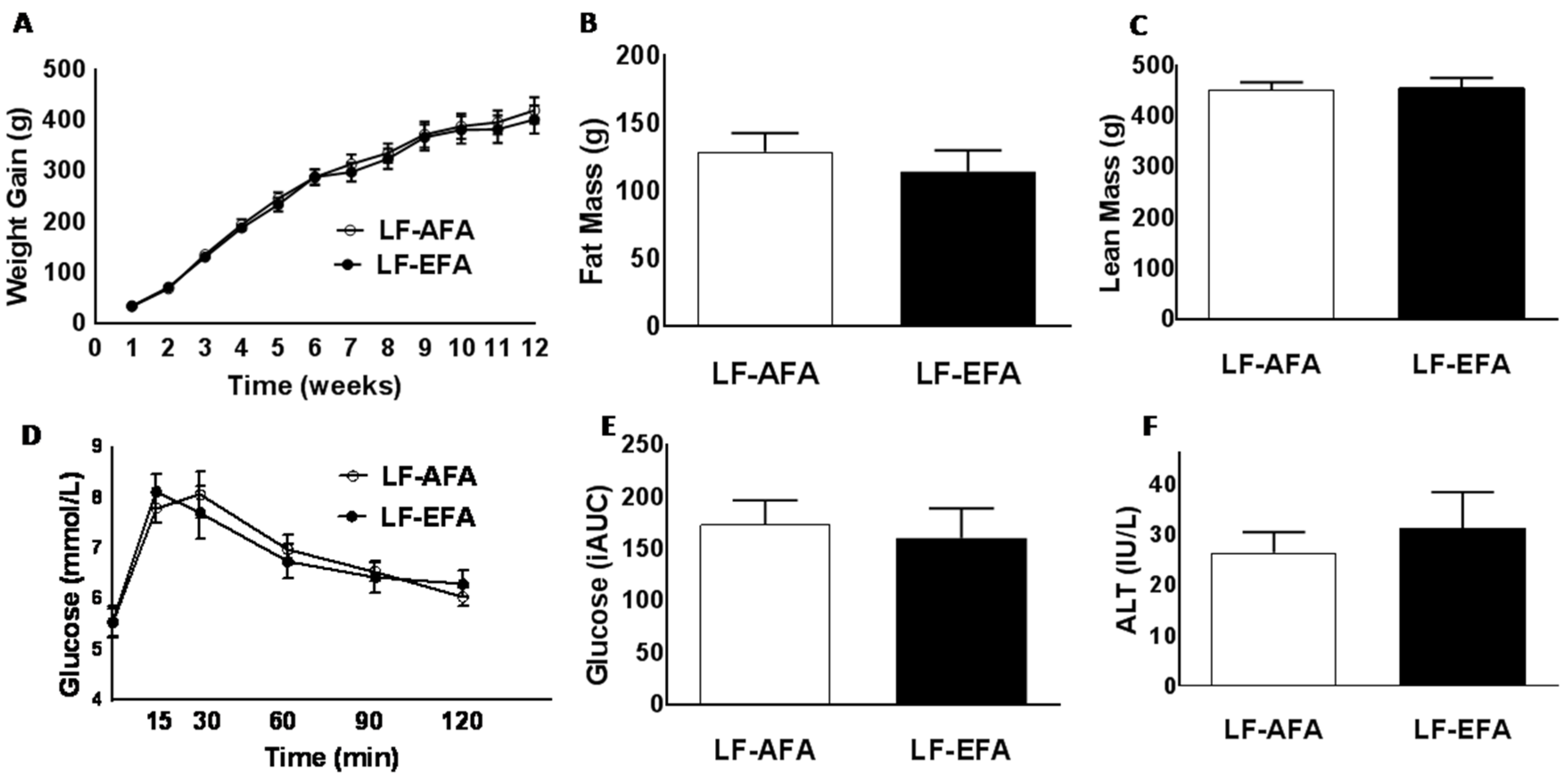
3.1. Excess Folic Acid Intake Does Not Influence Body Weight, Body Composition, or Glucose Tolerance on a Low Fat Diet
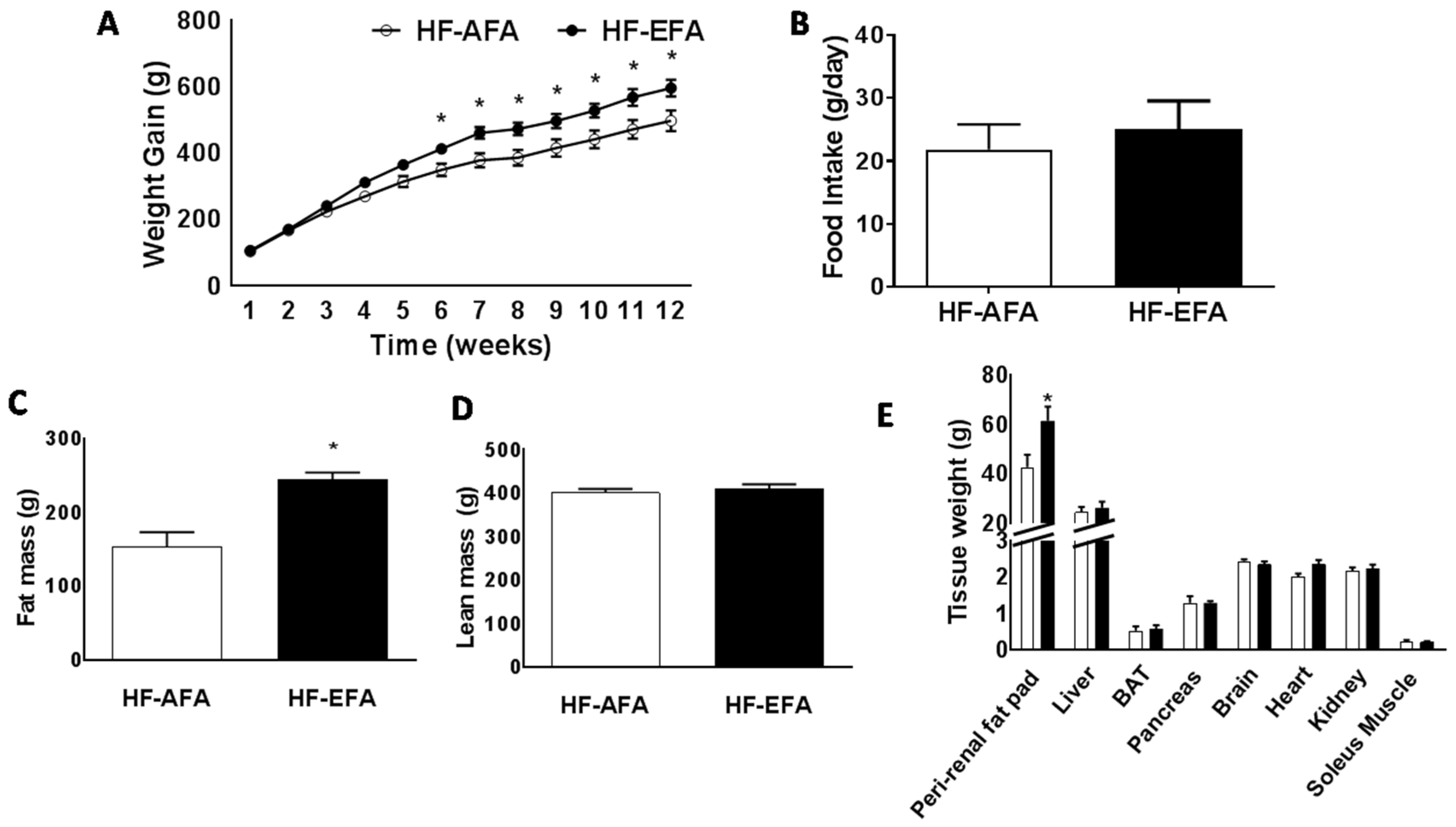
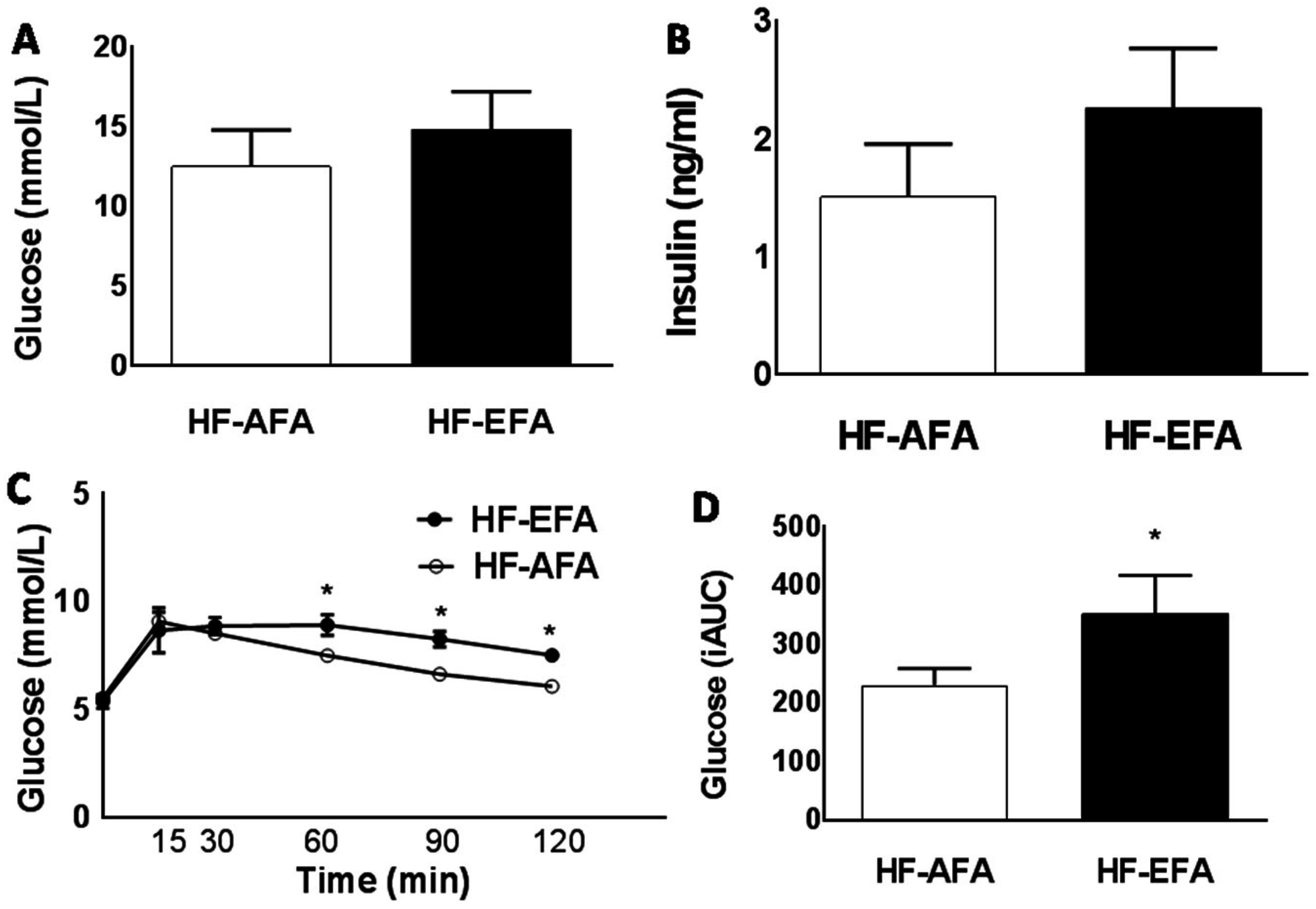
3.2. Excess Folic Acid Intake Increases Weight Gain, Fat Mass and Glucose Intolerance on a High Fat Diet
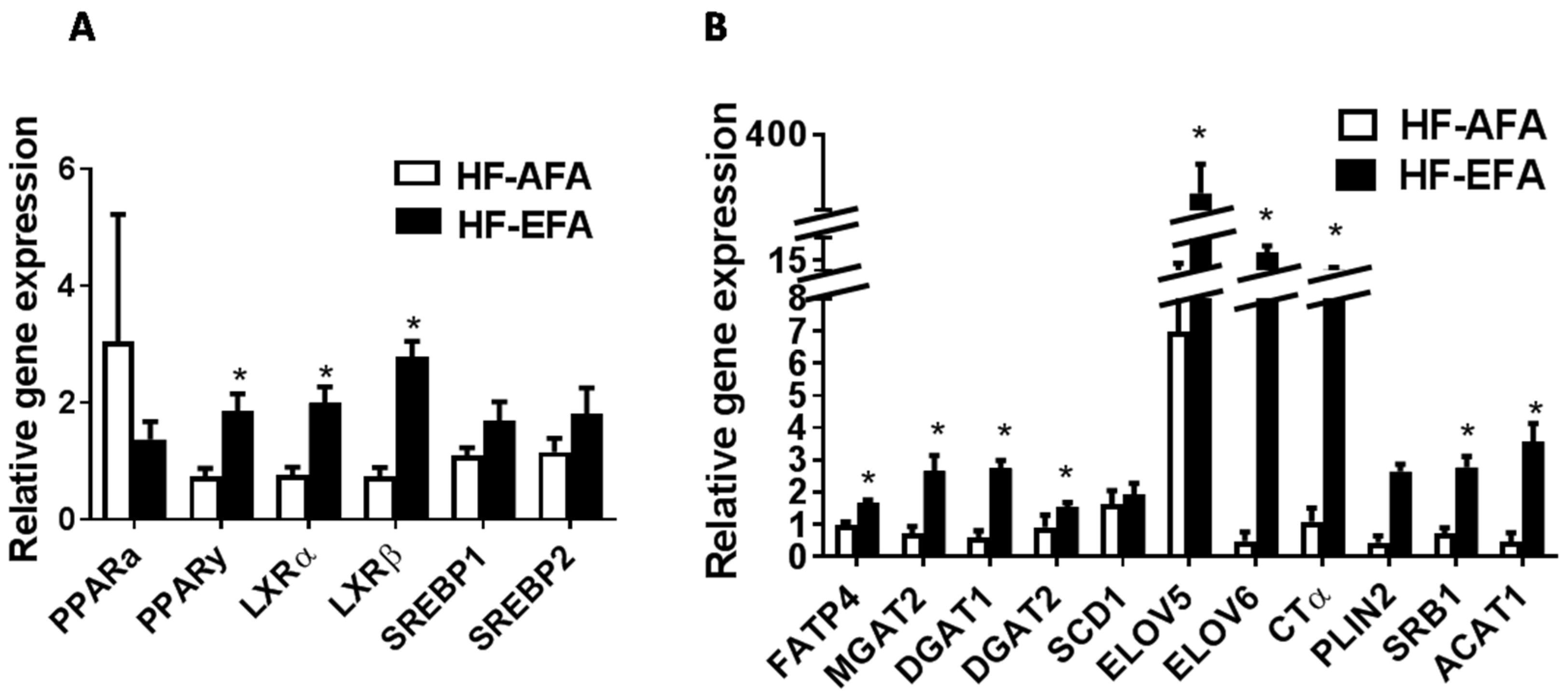
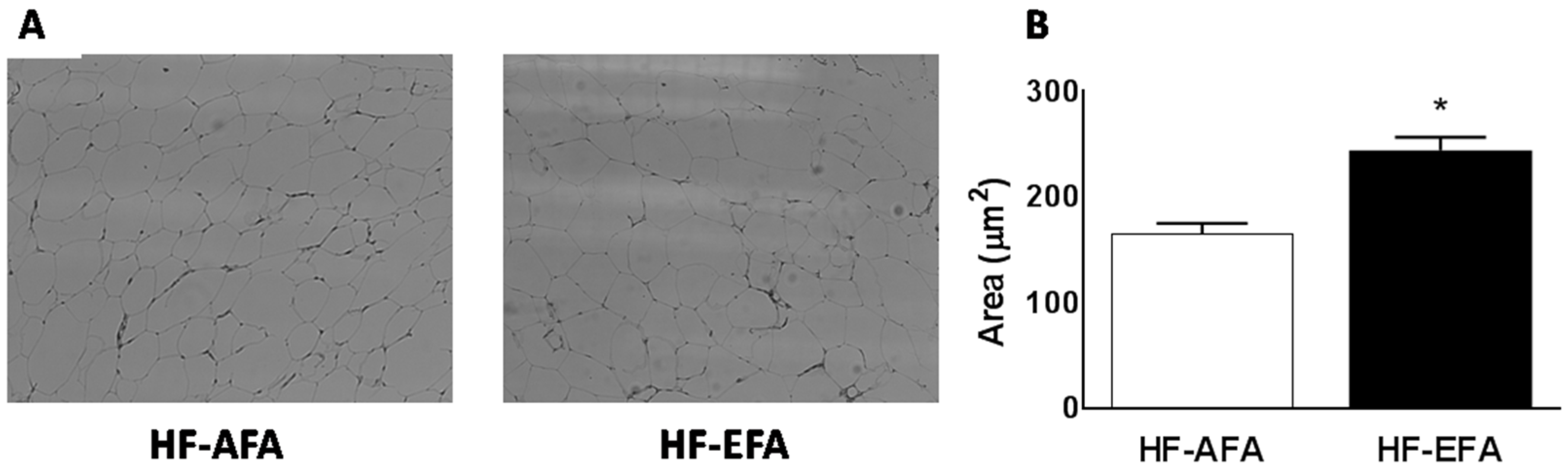
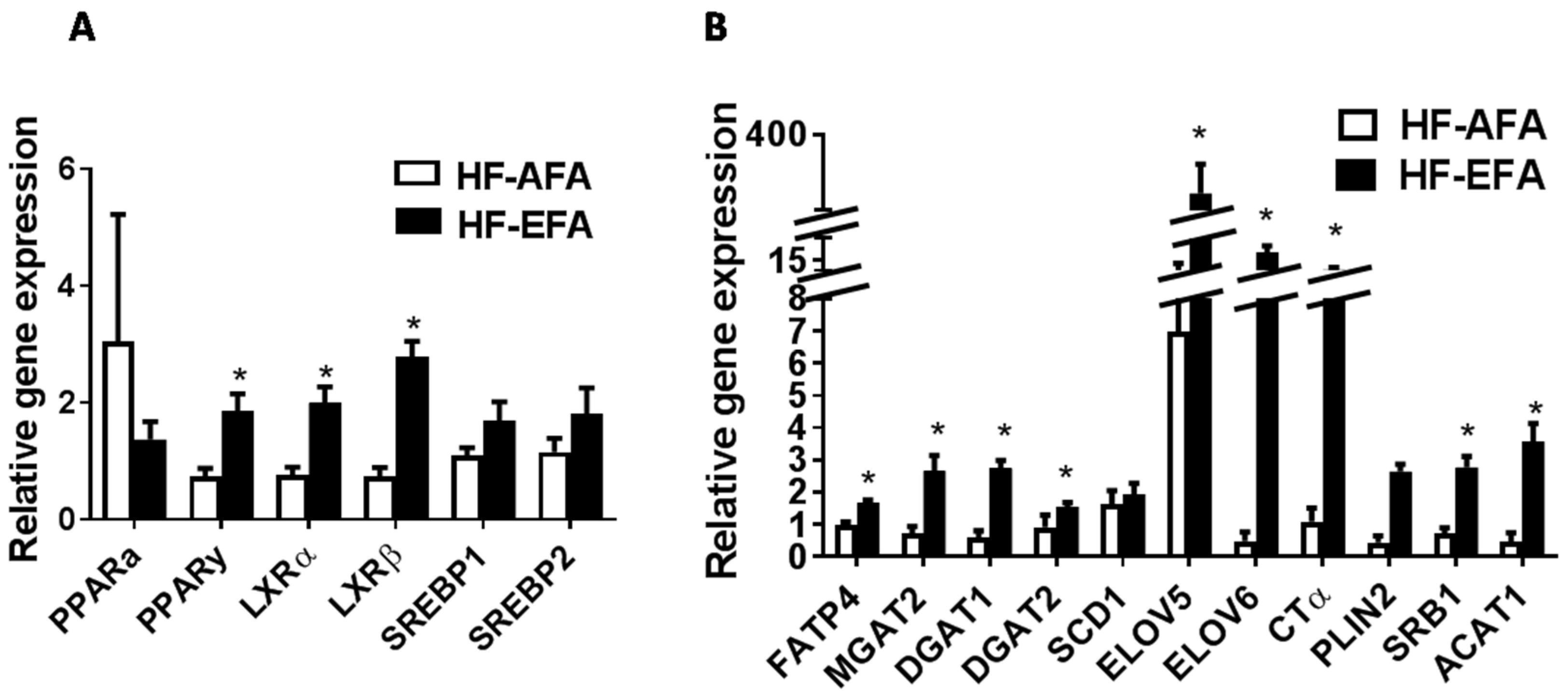
3.3. Excess Folic Acid Increases Adipose Tissue Size and Mass By Inducing Lipogenic Genes in High Fat Diet-Fed Rats
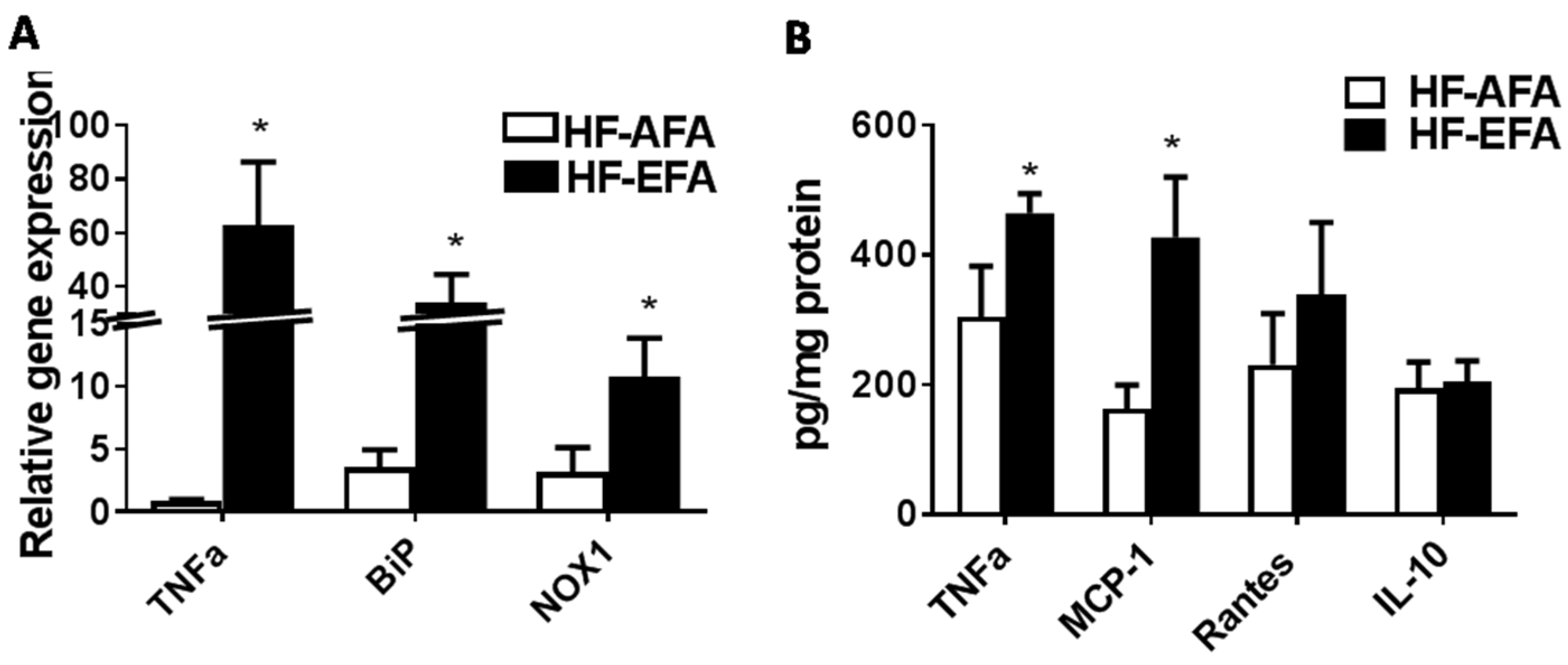
3.4. Excess Folic Acid Increases Inflammation in White Adipose Tissue
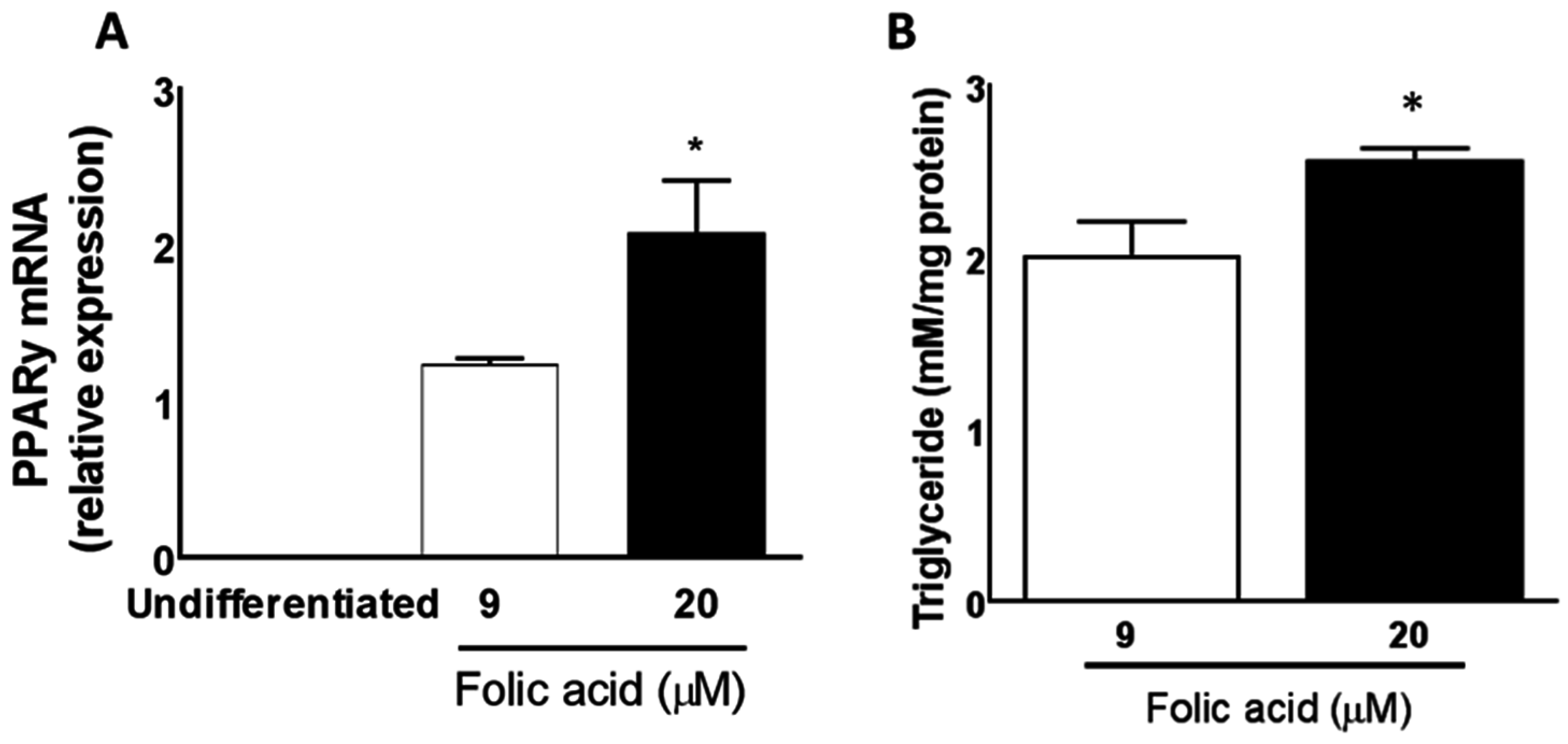
3.5. Excess Folic Acid Promotes Triglyceride Accumulation in Mature 3T3L1 Adipocytes
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- James, P.T.; Rigby, N.; Leach, R.; Force, I.O.T. The obesity epidemic, metabolic syndrome and future prevention strategies. Eur. J. Cardiovasc. Prev. Rehabil. 2004, 11, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [PubMed]
- Smith, A.D.; Kim, Y.I.; Refsum, H. Is folic acid good for everyone? Am. J. Clin. Nutr. 2008, 87, 517–533. [Google Scholar] [PubMed]
- Chan, Y.M.; Bailey, R.; O’Connor, D.L. Folate. Adv. Nutr. 2013, 4, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (U.S.) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes; Institute of Medicine (U.S.) Panel on Folate Other B Vitamins and Choline; Institute of Medicine (U.S.) Subcommittee on Upper Reference Levels of Nutrients. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; National Academy Press: Washington, DC, USA, 1998; p. 564. [Google Scholar]
- Pfeiffer, C.M.; Sternberg, M.R.; Fazili, Z.; Yetley, E.A.; Lacher, D.A.; Bailey, R.L.; Johnson, C.L. Unmetabolized folic acid is detected in nearly all serum samples from US children, adolescents, and adults. J. Nutr. 2015, 145, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.L.; Mills, J.L.; Yetley, E.A.; Gahche, J.J.; Pfeiffer, C.M.; Dwyer, J.T.; Dodd, K.W.; Sempos, C.T.; Betz, J.M.; Picciano, M.F. Serum unmetabolized folic acid in a nationally representative sample of adults ≥60 years in the United States, 2001–2002. Food Nutr. Res. 2012, 56. [Google Scholar] [CrossRef] [PubMed]
- Fayyaz, F.; Wang, F.; Jacobs, R.L.; O’Connor, D.L.; Bell, R.C.; Field, C.J.; Team, A.S. Folate, vitamin B12, and vitamin B6 status of a group of high socioeconomic status women in the Alberta Pregnancy Outcomes and Nutrition (APrON) cohort. Appl. Physiol. Nutr. Metab. 2014, 39, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- McNulty, B.; McNulty, H.; Marshall, B.; Ward, M.; Molloy, A.M.; Scott, J.M.; Dornan, J.; Pentieva, K. Impact of continuing folic acid after the first trimester of pregnancy: Findings of a randomized trial of Folic Acid Supplementation in the Second and Third Trimesters. Am. J. Clin. Nutr. 2013, 98, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Pentieva, K.; Selhub, J.; Paul, L.; Molloy, A.M.; McNulty, B.; Ward, M.; Marshall, B.; Dornan, J.; Reilly, R.; Parle-McDermott, A.; et al. Evidence from a Randomized Trial That Exposure to Supplemental Folic Acid at Recommended Levels during Pregnancy Does Not Lead to Increased Unmetabolized Folic Acid Concentrations in Maternal or Cord Blood. J. Nutr. 2016, 146, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Fife, J.; Raniga, S.; Hider, P.N.; Frizelle, F.A. Folic acid supplementation and colorectal cancer risk: A meta-analysis. Colorectal Dis. 2011, 13, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Hultdin, J.; Van Guelpen, B.; Bergh, A.; Hallmans, G.; Stattin, P. Plasma folate, vitamin B12, and homocysteine and prostate cancer risk: A prospective study. Int. J. Cancer 2005, 113, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Troen, A.M.; Mitchell, B.; Sorensen, B.; Wener, M.H.; Johnston, A.; Wood, B.; Selhub, J.; McTiernan, A.; Yasui, Y.; Oral, E.; et al. Unmetabolized folic acid in plasma is associated with reduced natural killer cell cytotoxicity among postmenopausal women. J. Nutr. 2006, 136, 189–194. [Google Scholar] [PubMed]
- Sawaengsri, H.; Wang, J.; Reginaldo, C.; Steluti, J.; Wu, D.; Meydani, S.N.; Selhub, J.; Paul, L. High folic acid intake reduces natural killer cell cytotoxicity in aged mice. J. Nutr. Biochem. 2016, 30, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Hebert, L.E.; Scherr, P.A.; Schneider, J.A. Dietary folate and vitamin B12 intake and cognitive decline among community-dwelling older persons. Arch. Neurol. 2005, 62, 641–645. [Google Scholar] [CrossRef] [PubMed]
- Bird, J.K.; Ronnenberg, A.G.; Choi, S.W.; Du, F.; Mason, J.B.; Liu, Z. Obesity is associated with increased red blood cell folate despite lower dietary intakes and serum concentrations. J. Nutr. 2015, 145, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.E.; Sánchez-Hernández, D.; Reza-López, S.A.; Huot, P.S.; Kim, Y.I.; Anderson, G.H. High folate gestational and post-weaning diets alter hypothalamic feeding pathways by DNA methylation in Wistar rat offspring. Epigenetics 2013, 8, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Hoile, S.P.; Lillycrop, K.A.; Grenfell, L.R.; Hanson, M.A.; Burdge, G.C. Increasing the folic acid content of maternal or post-weaning diets induces differential changes in phosphoenolpyruvate carboxykinase mRNA expression and promoter methylation in rats. Br. J. Nutr. 2012, 108, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Szeto, I.M.; Aziz, A.; Das, P.J.; Taha, A.Y.; Okubo, N.; Reza-Lopez, S.; Giacca, A.; Anderson, G.H. High multivitamin intake by Wistar rats during pregnancy results in increased food intake and components of the metabolic syndrome in male offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R575–R582. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; He, Y.; Sun, X.; Li, Y.; Sun, C. Maternal high folic acid supplement promotes glucose intolerance and insulin resistance in male mouse offspring fed a high-fat diet. Int. J. Mol. Sci. 2014, 15, 6298–6313. [Google Scholar] [CrossRef] [PubMed]
- Szeto, I.M.; Das, P.J.; Aziz, A.; Anderson, G.H. Multivitamin supplementation of Wistar rats during pregnancy accelerates the development of obesity in offspring fed an obesogenic diet. Int. J. Obes. (Lond.) 2009, 33, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Yajnik, C.S.; Deshpande, S.S.; Jackson, A.A.; Refsum, H.; Rao, S.; Fisher, D.J.; Bhat, D.S.; Naik, S.S.; Coyaji, K.J.; Joglekar, C.V.; et al. Vitamin B12 and folate concentrations during pregnancy and insulin resistance in the offspring: The Pune Maternal Nutrition Study. Diabetologia 2008, 51, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Lillycrop, K.A.; Phillips, E.S.; Slater-Jefferies, J.L.; Jackson, A.A.; Hanson, M.A. Folic acid supplementation during the juvenile-pubertal period in rats modifies the phenotype and epigenotype induced by prenatal nutrition. J. Nutr. 2009, 139, 1054–1060. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.A.; Mathers, J.C. Diet induced epigenetic changes and their implications for health. Acta Physiol. (Oxf.) 2011, 202, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.E.; Mikael, L.G.; Leung, K.Y.; Lévesque, N.; Deng, L.; Wu, Q.; Malysheva, O.V.; Best, A.; Caudill, M.A.; Greene, N.D.; et al. High folic acid consumption leads to pseudo-MTHFR deficiency, altered lipid metabolism, and liver injury in mice. Am. J. Clin. Nutr. 2015, 101, 646–658. [Google Scholar] [CrossRef] [PubMed]
- Stabler, S.P.; Allen, R.H. Quantification of serum and urinary S-adenosylmethionine and S-adenosylhomocysteine by stable-isotope-dilution liquid chromatography-mass spectrometry. Clin. Chem. 2004, 50, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Myher, J.J.; Kuksis, A. Determination of plasma total lipid profiles by capillary gas-liquid chromatography. J. Biochem. Biophys. Methods 1984, 10, 13–23. [Google Scholar] [CrossRef]
- Da Silva, R.P.; Kelly, K.B.; Leonard, K.A.; Jacobs, R.L. Creatine reduces hepatic TG accumulation in hepatocytes by stimulating fatty acid oxidation. Biochim. Biophys. Acta 2014, 1841, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Korf, H.; Vander Beken, S.; Romano, M.; Steffensen, K.R.; Stijlemans, B.; Gustafsson, J.A.; Grooten, J.; Huygen, K. Liver X receptors contribute to the protective immune response against Mycobacterium tuberculosis in mice. J. Clin. Investig. 2009, 119, 1626–1637. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Tontonoz, P. Endocrine functions of adipose tissue. Annu. Rev. Pathol. 2007, 2, 31–56. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Ogawa, Y. Adipose tissue macrophages: Their role in adipose tissue remodeling. J. Leukoc. Biol. 2010, 88, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Pannia, E.; Cho, C.E.; Kubant, R.; Sánchez-Hernández, D.; Huot, P.S.; Chatterjee, D.; Fleming, A.; Anderson, G.H. A high multivitamin diet fed to Wistar rat dams during pregnancy increases maternal weight gain later in life and alters homeostatic, hedonic and peripheral regulatory systems of energy balance. Behav. Brain Res. 2015, 278, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Symonds, M.E. Adipose Tissue Biology; Springer: New York, NY, USA, 2012; p. 413. [Google Scholar]
- Sie, K.K.; Li, J.; Ly, A.; Sohn, K.J.; Croxford, R.; Kim, Y.I. Effect of maternal and postweaning folic acid supplementation on global and gene-specific DNA methylation in the liver of the rat offspring. Mol. Nutr. Food Res. 2013, 57, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Ehara, T.; Kamei, Y.; Takahashi, M.; Yuan, X.; Kanai, S.; Tamura, E.; Tanaka, M.; Yamazaki, T.; Miura, S.; Ezaki, O.; et al. Role of DNA methylation in the regulation of lipogenic glycerol-3-phosphate acyltransferase 1 gene expression in the mouse neonatal liver. Diabetes 2012, 61, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Hunter, D.; Huber, R.; Lemieux, J.; Slaymaker, S.; Vaddi, K.; Charo, I.; Leibel, R.L.; Ferrante, A.W. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J. Clin. Investig. 2006, 116, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Molnar, C.; Geiger, S.; Graziadei, I.; Ebenbichler, C.F.; Weiss, H.; Kaser, S.; Kaser, A.; Tilg, H. Anti-inflammatory effects of excessive weight loss: Potent suppression of adipose interleukin 6 and tumour necrosis factor alpha expression. Gut 2010, 59, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, R.A.; Sheedfar, F.; Morrison, M.C.; Hommelberg, P.P.; Kor, D.; Kloosterhuis, N.J.; Gruben, N.; Youssef, S.A.; de Bruin, A.; Hofker, M.H.; et al. High-fat diet induced obesity primes inflammation in adipose tissue prior to liver in C57BL/6j mice. Aging 2015, 7, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.P.; Kelly, K.B.; Al Rajabi, A.; Jacobs, R.L. Novel insights on interactions between folate and lipid metabolism. Biofactors 2014, 40, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Qiu, A.; Jansen, M.; Sakaris, A.; Min, S.H.; Chattopadhyay, S.; Tsai, E.; Sandoval, C.; Zhao, R.; Akabas, M.H.; Goldman, I.D. Identification of an intestinal folate transporter and the molecular basis for hereditary folate malabsorption. Cell 2006, 127, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.W.; Ayling, J.E. The extremely slow and variable activity of dihydrofolate reductase in human liver and its implications for high folic acid intake. Proc. Natl. Acad. Sci. USA 2009, 106, 15424–15429. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | LF-AFA | LF-EFA | HF-AFA | HF-EFA |
|---|---|---|---|---|
| Folic acid (mg) | 0.75 | 7.5 | 0.75 | 7.5 |
| l-cysteine (g) | 3 | 3 | 4 | 4 |
| Corn starch (g) | 263.7 | 263.7 | - | - |
| Sucrose (g) | 209.7 | 209.7 | 106.3 | 106.3 |
| Maltodextrin (g) | 130 | 130 | 160 | 160 |
| Soybean Oil (g) | 60 | 60 | 30 | 30 |
| Lard (g) | - | - | 310 | 310 |
| Cellulose (g) | 50 | 50 | 20 | 20 |
| Pectin (g) | 50 | 50 | 50 | 50 |
| Succinylsulphathiazole (g) | 10 | 10 | 10 | 10 |
| Vitamin-free casein (g) | 195 | 195 | 265 | 265 |
| Mineral Mix, AIN-93G (g) | 35 | 35 | 48 | 48 |
| Tertiary-Butylhydroquinone (mg) | 12 | 12 | 3400 | 3400 |
| Choline bitartrate (g) | 2.5 | 2.5 | 3 | 3 |
| Niacin (mg) | 30 | 30 | 63 | 63 |
| Calcium pantothenate (mg) | 16 | 16 | 34 | 34 |
| Pyridoxine HCl (μg) | 7 | 7 | 15 | 15 |
| Thiamin HCl (μg) | 6 | 6 | 13 | 13 |
| Riboflavin (mg) | 6 | 6 | 13 | 13 |
| Biotin (μg) | 200 | 200 | 400 | 400 |
| Vitamin B12 (μg) | 25 | 25 | 40 | 40 |
| dl-alpha tocopheryl acetate (500 IU/g) (mg) | 150 | 150 | 315 | 315 |
| Vitamin A palmitate (500,000 IU/g) (mg) | 8 | 8 | 17 | 17 |
| Cholecalciferol (500,000 IU/g) (mg) | 2 | 2 | 4 | 4 |
| Phylloquinone (μg) | 800 | 800 | 1600 | 1600 |
| HF-AFA | HF-EFA | |
|---|---|---|
| Plasma one carbon metabolites | ||
| Folate (nmol/L) | 41.56 ± 0.50 | 41.30 ± 0.72 |
| Homocysteine (μM) | 3.28 ± 0.17 | 2.650 ± 0.14 * |
| Methionine (μM) | 69.73 ± 2.93 | 62.58 ± 1.01 * |
| Dimethylglycine (μM) | 13.10 ± 0.98 | 15.68 ± 0.95 |
| Methylglycine (μM) | 7.19 ± 0.48 | 6.21 ± 0.31 |
| Glycine (μM) | 425.0 ± 21.6 | 345.5 ± 24.0 * |
| Serine (μM) | 339.2 ± 11.2 | 364.5 ± 10.9 |
| Cystathionine (nM) | 914.7 ± 69.2 | 763.3 ± 37.2 |
| Cysteine (nM) | 311.3 ± 4.5 | 305.7 ± 9.7 |
| α-aminobutyrate (μM) | 27.68 ± 2.6 | 35.60 ± 4.0 |
| Plasma lipids | ||
| Triglyceride (µg/mL) | 292.6 ± 43.26 | 211.8 ±46.21 |
| Cholesterol Ester (µg/mL) | 72.92 ± 5.87 | 60.18 ± 7.97 |
| Free Cholesterol (µg/mL) | 134.4 ± 18.65 | 118.1 ± 15.41 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kelly, K.B.; Kennelly, J.P.; Ordonez, M.; Nelson, R.; Leonard, K.; Stabler, S.; Gomez-Muñoz, A.; Field, C.J.; Jacobs, R.L. Excess Folic Acid Increases Lipid Storage, Weight Gain, and Adipose Tissue Inflammation in High Fat Diet-Fed Rats. Nutrients 2016, 8, 594. https://doi.org/10.3390/nu8100594
Kelly KB, Kennelly JP, Ordonez M, Nelson R, Leonard K, Stabler S, Gomez-Muñoz A, Field CJ, Jacobs RL. Excess Folic Acid Increases Lipid Storage, Weight Gain, and Adipose Tissue Inflammation in High Fat Diet-Fed Rats. Nutrients. 2016; 8(10):594. https://doi.org/10.3390/nu8100594
Chicago/Turabian StyleKelly, Karen B., John P. Kennelly, Marta Ordonez, Randal Nelson, Kelly Leonard, Sally Stabler, Antonio Gomez-Muñoz, Catherine J. Field, and René L. Jacobs. 2016. "Excess Folic Acid Increases Lipid Storage, Weight Gain, and Adipose Tissue Inflammation in High Fat Diet-Fed Rats" Nutrients 8, no. 10: 594. https://doi.org/10.3390/nu8100594