Maternal Dietary Vitamin D Does Not Program Systemic Inflammation and Bone Health in Adult Female Mice Fed an Obesogenic Diet

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
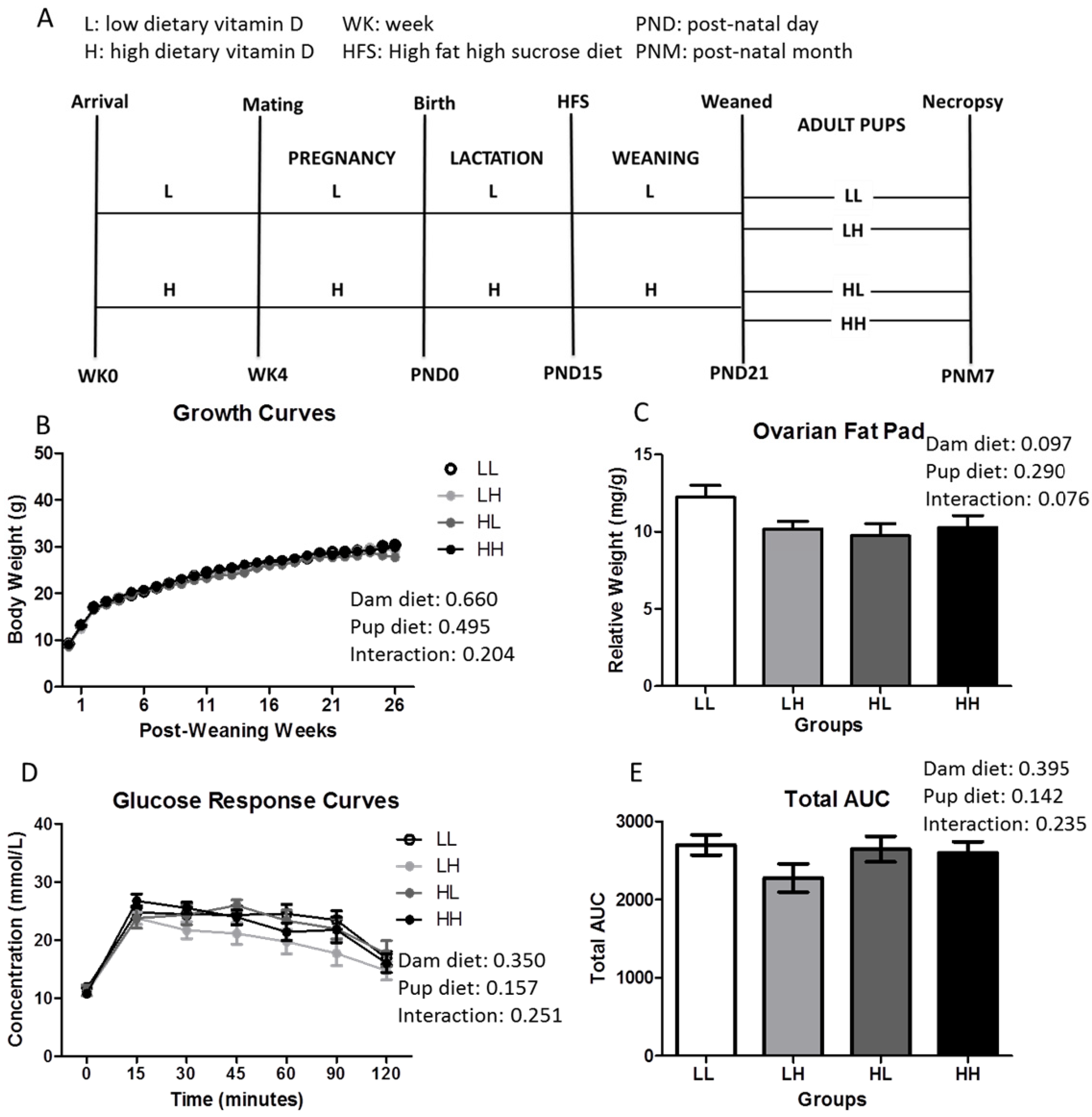
2.1. Animal Study
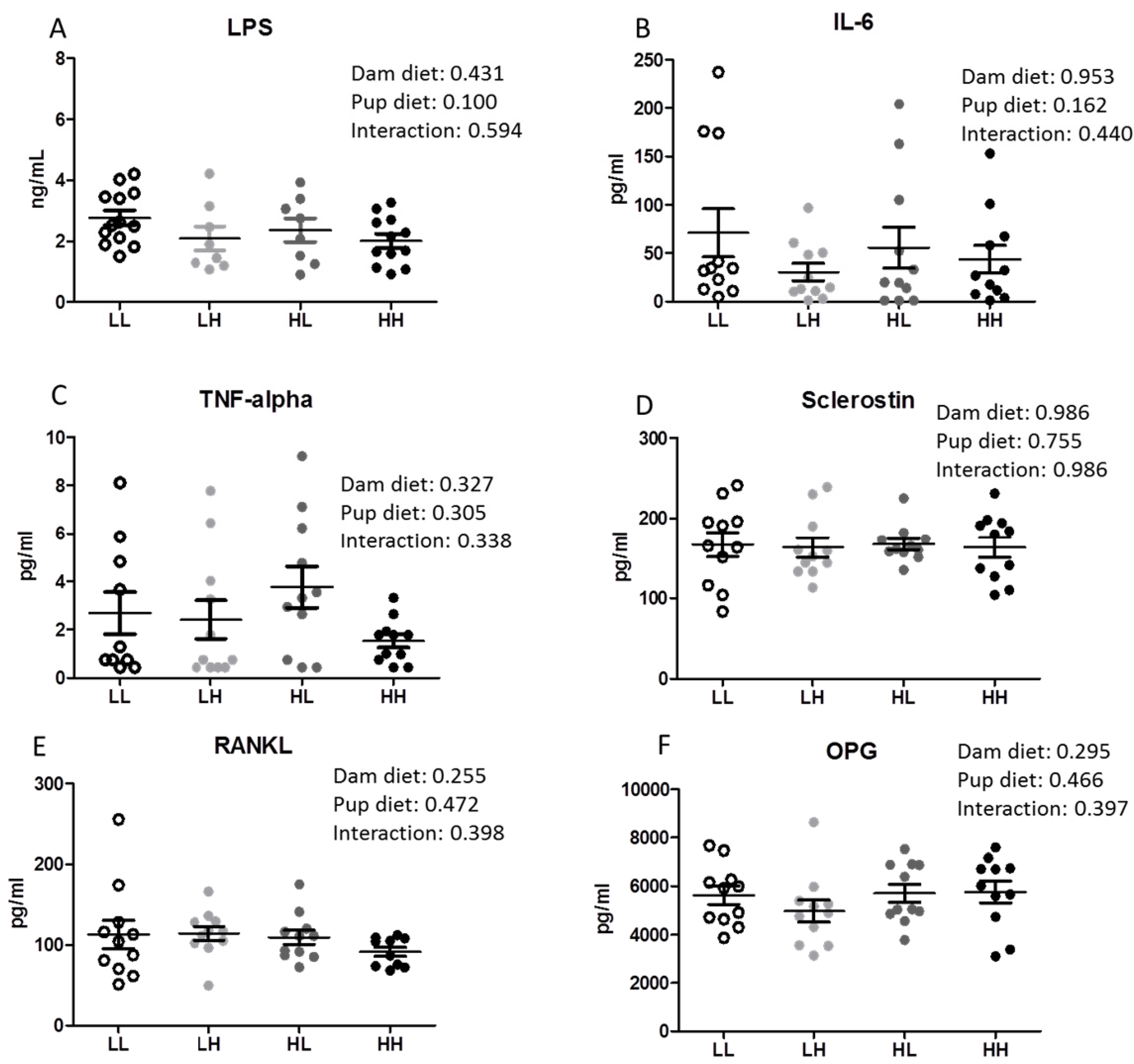
2.2. Biochemical Analyses
2.3. Glucose Response
2.4. Bone Mineral Content (BMC), Bone Mineral Density (BMD), and Biomechanical Strength of Femurs and Lumbar Vertebrae
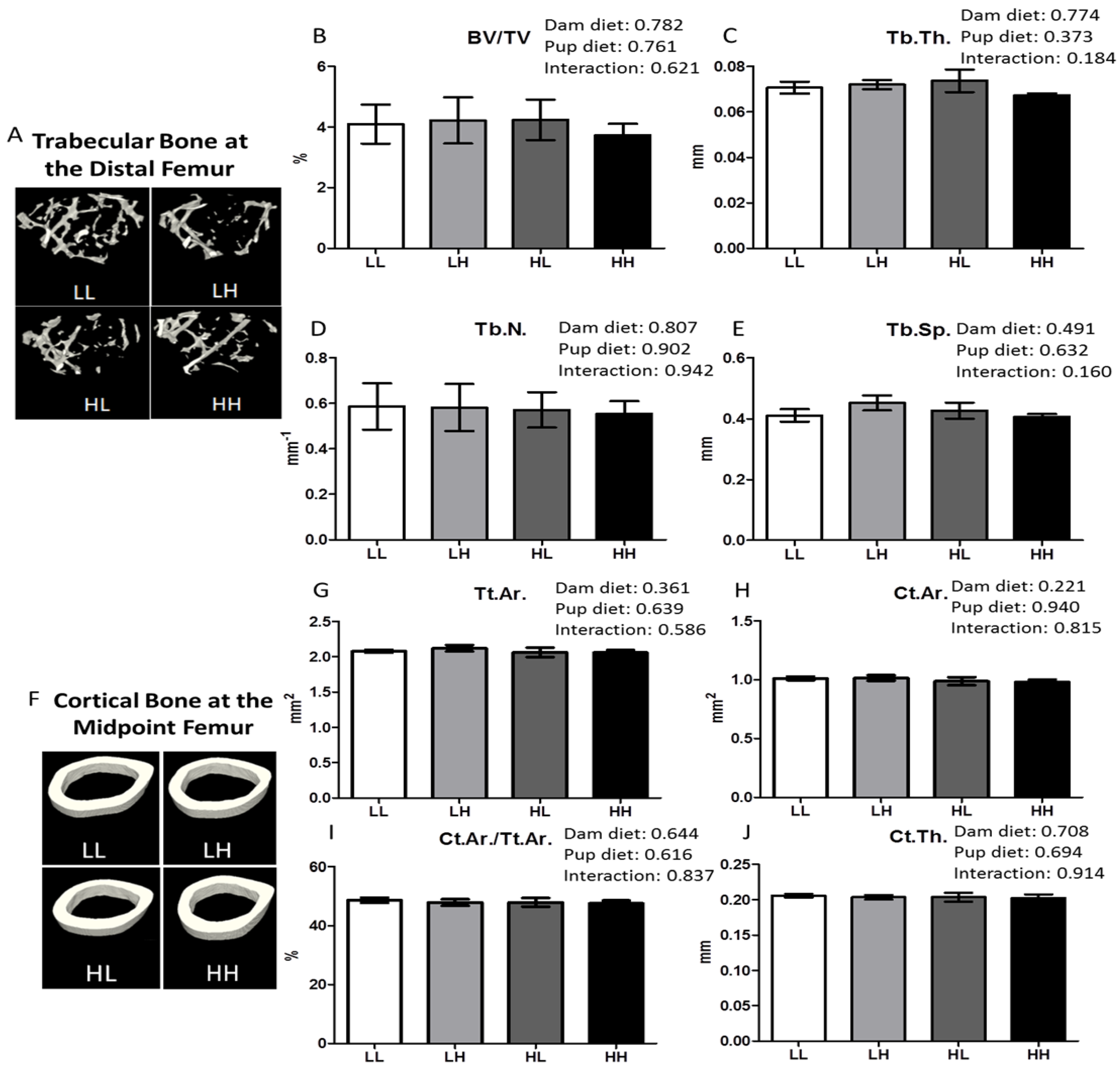
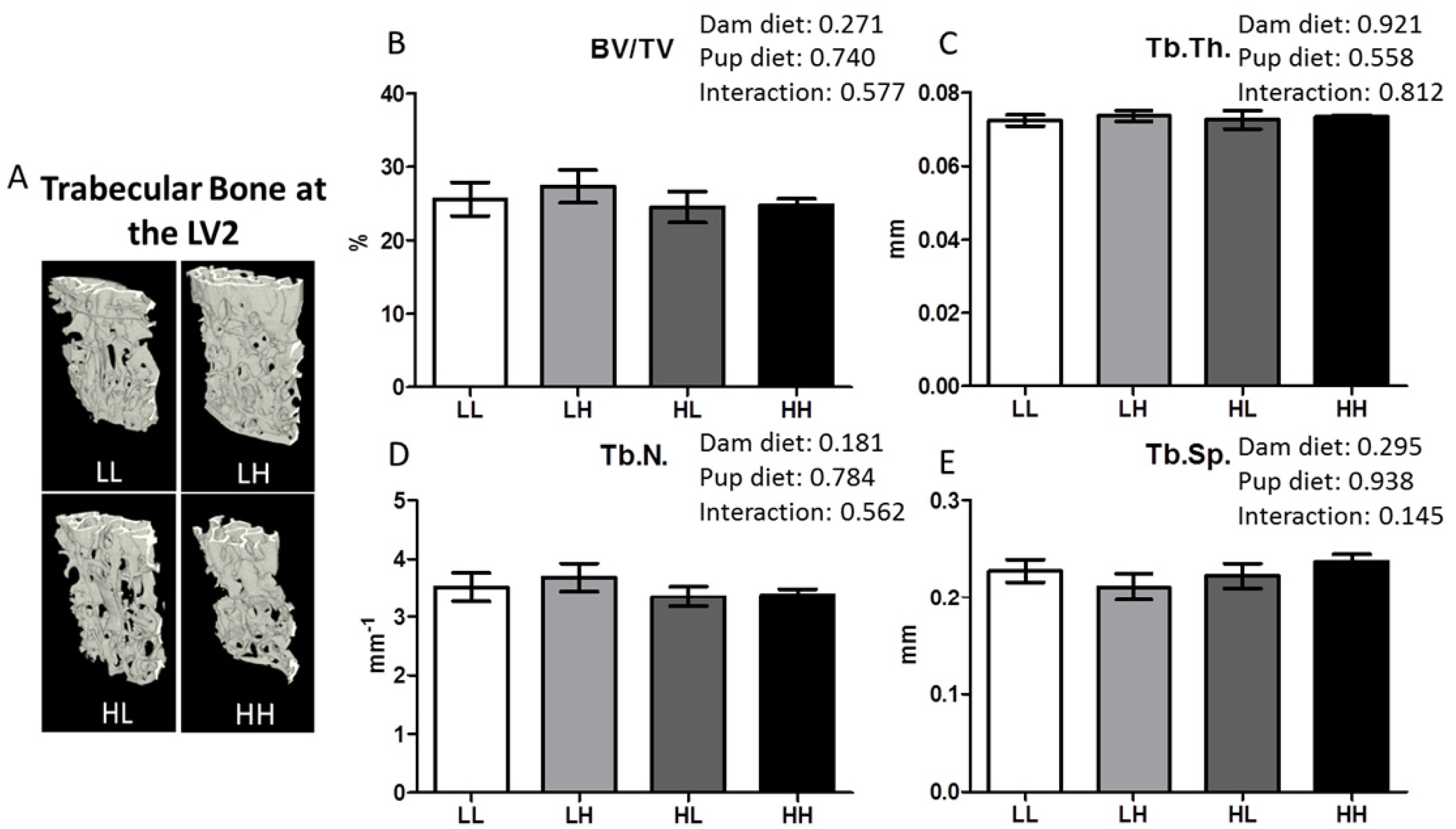
2.5. Structure of Femurs and Lumbar Vertebrae Using Micro Computed Tomography
2.5.1. Trabecular Bone Structure
2.5.2. Cortical Bone Structure
2.6. Statistics
3. Results
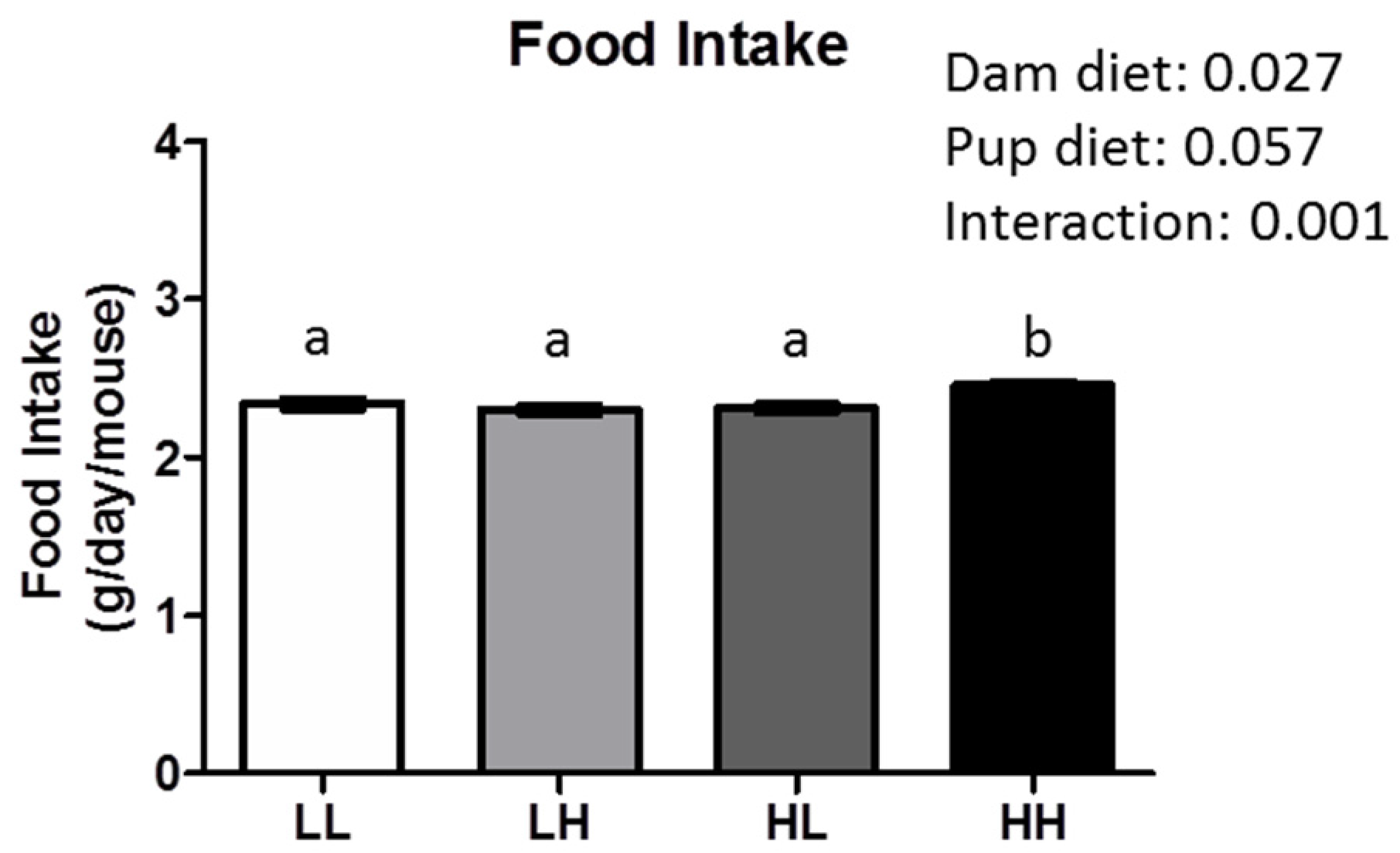
3.1. High Dietary Vitamin D, Pre- and/or Post-Weaning, Does Not Affect Weight, Adiposity, or Glycemia at Seven Months of Age
3.2. High Dietary Vitamin D Does Not Program Systemic Inflammation
3.3. Post-Weaning, but Not Pre-Weaning, Consumption of High Dietary Vitamin D Improves Mineralization in the Lumbar Vertebra but Does Not Increase Strength of Trabecular or Cortical Bone
3.4. High Dietary Vitamin D, Pre- and/or Post-Weaning, Does Not Affect Bone Quality in the Trabecular Bone of the Metaphyseal or the Diaphyseal Region of the Femur
3.5. High Dietary Vitamin D, Pre- and/or Post-Weaning, Does Not Impact Trabecular Bone Quality at the Lumbar Vertebrae
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Formula | g/kg |
| Casein, “Vitamin-Free” Test | 245.0 |
| L-Cystine | 3.5 |
| Corn Starch | 85.0 |
| Maltodextrin | 115.0 |
| Sucrose | 199.7 |
| Vegetable Shortening, hydrogenated (Primex) | 195.0 |
| Soybean Oil | 30.0 |
| Cellulose | 58.0 |
| Mineral Mix, AIN-93G-MX (94046) | 43.0 |
| Calcium Phosphate, dibasic | 3.4 |
| Vitamin Mix, AIN-93 w/o A, D, E (120379) | 19.0 |
| Vitamin A Palmitate (500,000 IU/g) | 0.015 |
| Vitamin D3, cholecalciferol (50,000 IU/g in sucrose) | 0.1 † or 0.0005 †† |
| Vitamin E, DL-alpha tocopheryl acetate (500 IU/g) | 0.285 |
| Choline Bitartrate | 3.0 |
| Vitamin Mix | g/kg |
| Niacin | 3.0 |
| Calcium Pantothenate | 1.6 |
| Pyridoxine HCl | 0.7 |
| Thiamine HCl | 0.6 |
| Riboflavin | 0.6 |
| Folic Acid | 0.2 |
| Biotin | 0.02 |
| Vitamin B12 (0.1% in mannitol) | 2.5 |
| Vitamin K1, phylloquinone | 0.075 |
| Sucrose, fine ground | 990.705 |
| Mineral Mix | g/kg |
| Calcium Carbonate | 357.0 |
| Potassium Phosphate, monobasic | 196.0 |
| Potassium Citrate, monohydrate | 70.78 |
| Sodium Chloride | 74.0 |
| Potassium Sulfate | 46.6 |
| Magnesium Oxide | 24.3 |
| Ferric Citrate | 6.06 |
| Zinc Carbonate | 1.65 |
| Manganous Carbonate | 0.63 |
| Cupric Carbonate | 0.31 |
| Potassium Iodate | 0.01 |
| Sodium Selenate | 0.0103 |
| Ammonium Paramolybdate, tetrahydrate | 0.008 |
| Sodium Meta-Silicate, nonahydrate | 1.45 |
| Chromium Potassium Sulfate, dodecahydrate | 0.275 |
| Lithium Chloride | 0.0174 |
| Boric Acid | 0.0815 |
| Sodium Fluoride | 0.0635 |
| Nickel Carbonate Hydroxide, tetrahydrate | 0.0318 |
| Ammonium Meta-Vanadate | 0.0066 |
| Sucrose, fine ground | 220.7159 |
| Macronutrient | % kcal |
| Protein | 19.6 |
| Fat | 44.2 |
| Carbohydrate (Sucrose) | 36.2 (19.8) |

References
- World Health Organization. Obesity and Overweight: Fact Sheet. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 16 June 2016).
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Morrison, H.; de Groh, M.; Waters, C.; DesMeules, M.; Jones-McLean, E.; Ugnat, A.M.; Desjardins, S.; Lim, M.; Mao, Y. The burden of adult obesity in Canada. Chronic Dis. Can. 2007, 27, 135–144. [Google Scholar] [PubMed]
- Bray, G.A. Medical consequences of obesity. J. Clin. Endocrinol. Metab. 2004, 89, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Compston, J.E.; Watts, N.B.; Chapurlat, R.; Cooper, C.; Boonen, S.; Greenspan, S.; Pfeilschifter, J.; Silverman, S.; Diez-Perez, A.; Lindsay, R.; et al. Obesity is not protective against fracture in postmenopausal women: GLOW. Am. J. Med. 2011, 124, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Premaor, M.O.; Compston, J.E.; Fina Aviles, F.; Pages-Castella, A.; Nogues, X.; Diez-Perez, A.; Prieto-Alhambra, D. The association between fracture site and obesity in men: A population-based cohort study. J. Bone Miner. Res. 2013, 28, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Karim, L.; Bouxsein, M.L. Effect of type 2 diabetes-related non-enzymatic glycation on bone biomechanical properties. Bone 2016, 82, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Farr, J.N.; Khosla, S. Determinants of bone strength and quality in diabetes mellitus in humans. Bone 2016, 82, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.S.; Wu, W.; Chumlea, W.C.; Roche, A.F. Predicting overweight and obesity in adulthood from body mass index values in childhood and adolescence. Am. J. Clin. Nutr. 2002, 76, 653–658. [Google Scholar] [PubMed]
- Viljakainen, H.T.; Pekkinen, M.; Saarnio, E.; Karp, H.; Lamberg-Allardt, C.; Makitie, O. Dual effect of adipose tissue on bone health during growth. Bone 2011, 48, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Javaid, M.K.; Crozier, S.R.; Harvey, N.C.; Gale, C.R.; Dennison, E.M.; Boucher, B.J.; Arden, N.K.; Godfrey, K.M.; Cooper, C. Maternal vitamin D status during pregnancy and childhood bone mass at age 9 years: A longitudinal study. Lancet 2006, 367, 36–43. [Google Scholar] [CrossRef]
- Zhu, K.; Whitehouse, A.J.; Hart, P.H.; Kusel, M.; Mountain, J.; Lye, S.; Pennell, C.; Walsh, J.P. Maternal vitamin D status during pregnancy and bone mass in offspring at 20 years of age: A prospective cohort study. J. Bone Miner. Res. 2014, 29, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.C.; Sheppard, A.; Godfrey, K.M.; McLean, C.; Garratt, E.; Ntani, G.; Davies, L.; Murray, R.; Inskip, H.M.; Gluckman, P.D.; et al. Childhood bone mineral content is associated with methylation status of the RXRA promoter at birth. J. Bone Miner. Res. 2014, 29, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Soder, O. Perinatal imprinting by estrogen and adult prostate disease. Proc. Natl. Acad. Sci. USA 2005, 102, 1269–1270. [Google Scholar] [CrossRef] [PubMed]
- Nutrition UNESCO. Double burden of malnutrition—A common agenda. In Proceedings of the 33rd Annual Session of the Standing Committee on Nutrition, Geneva, Switzerland, 13–17 March 2006.
- Lucas, A. Long-term programming effects of early nutrition—Implications for the preterm infant. J. Perinatol. 2005, 25, S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Nielson, C.M.; Srikanth, P.; Orwoll, E.S. Obesity and fracture in men and women: An epidemiologic perspective. J. Bone Miner. Res. 2012, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mosca, L.N.; Goldberg, T.B.; da Silva, V.N.; da Silva, C.C.; Kurokawa, C.S.; Bisi Rizzo, A.C.; Corrente, J.E. Excess body fat negatively affects bone mass in adolescents. Nutrition 2014, 30, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Jahani, R.; Fielding, K.A.; Chen, J.; Villa, C.R.; Castelli, L.M.; Ward, W.E.; Comelli, E.M. Low vitamin D status throughout life results in an inflammatory prone status but does not alter bone mineral or strength in healthy 3-month-old CD-1 male mice. Mol. Nutr. Food Res. 2014, 58, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.J.; Lerner, M.R.; Bu, S.Y.; Lucas, E.A.; Hanas, J.S.; Lightfoot, S.A.; Postier, R.G.; Bronze, M.S.; Brackett, D.J. Systemic bone loss and induction of coronary vessel disease in a rat model of chronic inflammation. Bone 2006, 38, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.P.; Bellido, T.; Manolagas, S.C. Down-regulation of NF-kappa B protein levels in activated human lymphocytes by 1,25-dihydroxyvitamin D3. Proc. Natl. Acad. Sci. USA 1995, 92, 10990–10994. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Leung, D.Y.; Richers, B.N.; Liu, Y.; Remigio, L.K.; Riches, D.W.; Goleva, E. Vitamin D inhibits monocyte/macrophage proinflammatory cytokine production by targeting MAPK phosphatase-1. J. Immunol. 2012, 188, 2127–2135. [Google Scholar] [CrossRef] [PubMed]
- Villa, C.R.; Chen, J.; Wen, B.; Sacco, S.M.; Taibi, A.; Ward, W.E.; Comelli, E.M. Maternal vitamin D beneficially programs metabolic, gut and bone health of mouse male offspring in an obesogenic environment. Int. J. Obes. 2016. [Google Scholar] [CrossRef] [PubMed]
- Fleet, J.C.; Gliniak, C.; Zhang, Z.; Xue, Y.; Smith, K.B.; McCreedy, R.; Adedokun, S.A. Serum metabolite profiles and target tissue gene expression define the effect of cholecalciferol intake on calcium metabolism in rats and mice. J. Nutr. 2008, 138, 1114–1120. [Google Scholar] [PubMed]
- Kaludjerovic, J.; Ward, W.E. Neonatal exposure to daidzein, genistein, or the combination modulates bone development in female CD-1 mice. J. Nutr. 2009, 139, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Otsu, N. A threshold selection method from gray-level histograms. IEEE Trans. Syst. Man Cybern. 1979, 9, 62–66. [Google Scholar] [CrossRef]
- Huot, P.S.; Ly, A.; Szeto, I.M.; Reza-Lopez, S.A.; Cho, D.; Kim, Y.I.; Anderson, G.H. Maternal and postweaning folic acid supplementation interact to influence body weight, insulin resistance, and food intake regulatory gene expression in rat offspring in a sex-specific manner. Appl. Physiol. Nutr. Metab. 2016, 41, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Kaludjerovic, J.; Ward, W.E. Adequate but not supplemental folic acid combined with soy isoflavones during early life improves bone health at adulthood in male mice. J. Nutr. Biochem. 2013, 24, 1691–1696. [Google Scholar] [CrossRef] [PubMed]
- Kaludjerovic, J.; Ward, W.E. Bone-specific gene expression patterns and whole bone tissue of female mice are programmed by early life exposure to soy isoflavones and folic acid. J. Nutr. Biochem. 2015, 26, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Tailor, A.M.; Peeters, P.H.; Norat, T.; Vineis, P.; Romaguera, D. An update on the prevalence of the metabolic syndrome in children and adolescents. Int. J. Pediatr. Obes. 2010, 5, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Dimitri, P.; Bishop, N.; Walsh, J.S.; Eastell, R. Obesity is a risk factor for fracture in children but is protective against fracture in adults: A paradox. Bone 2012, 50, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, P.; Huen, K.; Dave, V.; Barcellos, L.; Eskenazi, B.; Holland, N. Sex differences in DNA methylation assessed by 450 K BeadChip in newborns. BMC Genom. 2015, 16, 911. [Google Scholar] [CrossRef] [PubMed]
- Body Weight Information for 000664. Available online: https://www.jax.org/jax-mice-and-services/strain-data-sheet-pages/body-weight-chart-000664 (accessed on 7 June 2016).
- Villa, C.R.; Ward, W.E.; Comelli, E.M. Gut Microbiota-bone Axis. Crit. Rev. Food Sci. Nutr. 2015. [Google Scholar] [CrossRef] [PubMed]
- Langsetmo, L.; Goltzman, D.; Kovacs, C.S.; Adachi, J.D.; Hanley, D.A.; Kreiger, N.; Josse, R.; Papaioannou, A.; Olszynski, W.P.; Jamal, S.A.; et al. Repeat low-trauma fractures occur frequently among men and women who have osteopenic BMD. J. Bone Miner. Res. 2009, 24, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Siris, E.S.; Chen, Y.T.; Abbott, T.A.; Barrett-Connor, E.; Miller, P.D.; Wehren, L.E.; Berger, M.L. Bone mineral density thresholds for pharmacological intervention to prevent fractures. Arch. Intern. Med. 2004, 164, 1108–1112. [Google Scholar] [CrossRef] [PubMed]
- Cranney, A.; Jamal, S.A.; Tsang, J.F.; Josse, R.G.; Leslie, W.D. Low bone mineral density and fracture burden in postmenopausal women. Can. Med. Assoc. J. 2007, 177, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.L.; Felton, S.K.; Riek, A.E.; Bernal-Mizrachi, C. Implications of vitamin D deficiency in pregnancy and lactation. Am. J. Obstet. Gynecol. 2010, 202, 429.e1–429.e9. [Google Scholar] [CrossRef] [PubMed]




| Dam Diet | Pup Diet | p-Value | ||||
|---|---|---|---|---|---|---|
| Low | High | Dam Diet | Pup Diet | Dam Diet × Pup Diet | ||
| Bone mineral | ||||||
| Whole femur | ||||||
| BMC, mg | Low | 25.5 ± 0.39 | 25.7 ± 0.36 | |||
| High | 26.1 ± 0.51 | 26.3 ± 0.43 | NS | NS | NS | |
| BMD, mg/cm2 | Low | 62.8 ± 0.77 | 62.7 ± 0.67 | |||
| High | 64.2 ± 0.71 | 63.7 ± 0.86 | NS | NS | NS | |
| 1/3 Proximal femur | ||||||
| BMC, mg | Low | 9.4 ± 0.20 | 9.9 ± 0.19 | |||
| High | 9.7 ± 0.23 | 9.8 ± 0.25 | NS | NS | NS | |
| BMD, mg/cm2 | Low | 67.9 ± 1.1 | 68.9 ± 0.85 | |||
| High | 68.9 ± 1.2 | 68.8 ± 1.10 | NS | NS | NS | |
| LV3 | ||||||
| BMC, mg | Low | 22.3 ± 0.48 | 22.9 ± 0.82 | |||
| High | 21.8 ± 0.60 | 24.2 ± 0.81 | NS | 0.037 | NS | |
| BMD, mg/cm2 | Low | 60.3 ± 1.02 | 61.7 ± 0.76 | |||
| High | 60.0 ± 0.90 | 62.5 ± 0.87 | NS | 0.015 | NS | |
| Biomechanical strength | ||||||
| Femur midpoint | ||||||
| Peak load, N | Low | 21.9 ± 0.85 | 21.8 ± 0.83 | |||
| High | 20.2 ± 0.53 | 21.1 ± 0.48 | NS | NS | NS | |
| Femur neck | ||||||
| Peak load, N | Low | 15.3 ± 0.90 | 16.6 ± 0.91 | |||
| High | 15.1 ± 1.95 | 14.1 ± 1.23 | NS | NS | NS | |
| LV3 | ||||||
| Peak load, N | Low | 42.8 ± 2.87 | 41.6 ± 3.60 | |||
| High | 37.7 ± 3.60 | 37.5 ± 3.74 | NS | NS | NS | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villa, C.R.; Chen, J.; Wen, B.; Sacco, S.M.; Taibi, A.; Ward, W.E.; Comelli, E.M. Maternal Dietary Vitamin D Does Not Program Systemic Inflammation and Bone Health in Adult Female Mice Fed an Obesogenic Diet. Nutrients 2016, 8, 675. https://doi.org/10.3390/nu8110675
Villa CR, Chen J, Wen B, Sacco SM, Taibi A, Ward WE, Comelli EM. Maternal Dietary Vitamin D Does Not Program Systemic Inflammation and Bone Health in Adult Female Mice Fed an Obesogenic Diet. Nutrients. 2016; 8(11):675. https://doi.org/10.3390/nu8110675
Chicago/Turabian StyleVilla, Christopher R., Jianmin Chen, Bijun Wen, Sandra M. Sacco, Amel Taibi, Wendy E. Ward, and Elena M. Comelli. 2016. "Maternal Dietary Vitamin D Does Not Program Systemic Inflammation and Bone Health in Adult Female Mice Fed an Obesogenic Diet" Nutrients 8, no. 11: 675. https://doi.org/10.3390/nu8110675
APA StyleVilla, C. R., Chen, J., Wen, B., Sacco, S. M., Taibi, A., Ward, W. E., & Comelli, E. M. (2016). Maternal Dietary Vitamin D Does Not Program Systemic Inflammation and Bone Health in Adult Female Mice Fed an Obesogenic Diet. Nutrients, 8(11), 675. https://doi.org/10.3390/nu8110675