Cuminaldehyde from Cinnamomum verum Induces Cell Death through Targeting Topoisomerase 1 and 2 in Human Colorectal Adenocarcinoma COLO 205 Cells

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Cell Viability XTT Test
2.4. Lactate Dehydrogenase Cytotoxicity Test
2.5. Test for Nuclear Fragmentation
2.6. Comet Test
2.7. Test for Volume of Acidic Compartments
2.8. Mitochondrial Membrane Potential Test
2.9. Caspase Activity Test
2.10. Test for Topoisomerase I and II Activities
2.11. In Vivo Tumor Xenograft Study
2.12. Statistical Analysis
3. Results
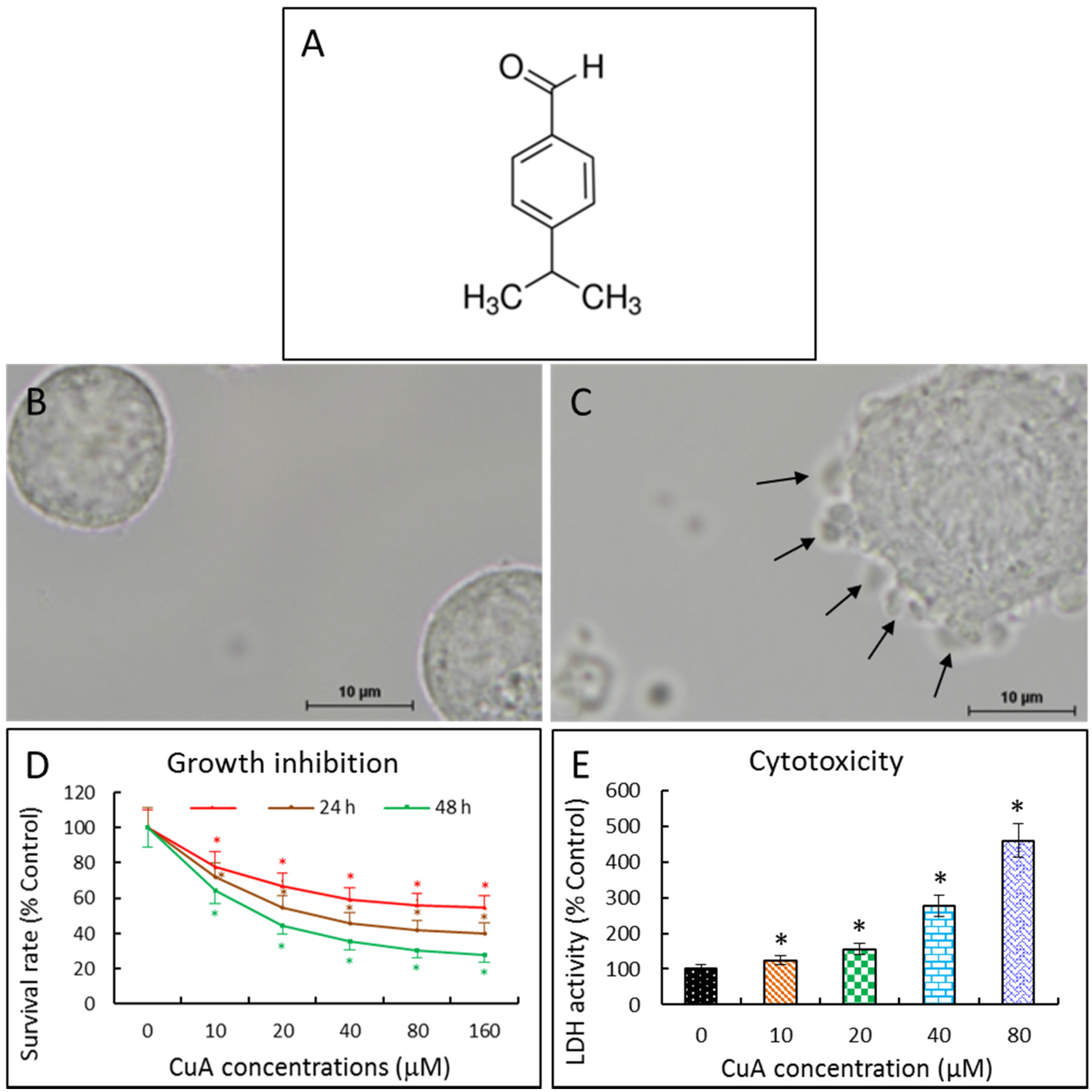
3.1. Cuminaldehyde’s Effects on Cell Morphological Changes
3.2. Cuminaldehyde Inhibited Human Colorectal COLO 205 Cell Proliferation
3.3. Cuminaldehyde Caused Cytotoxicity in Human Colorectal COLO 205 Cells
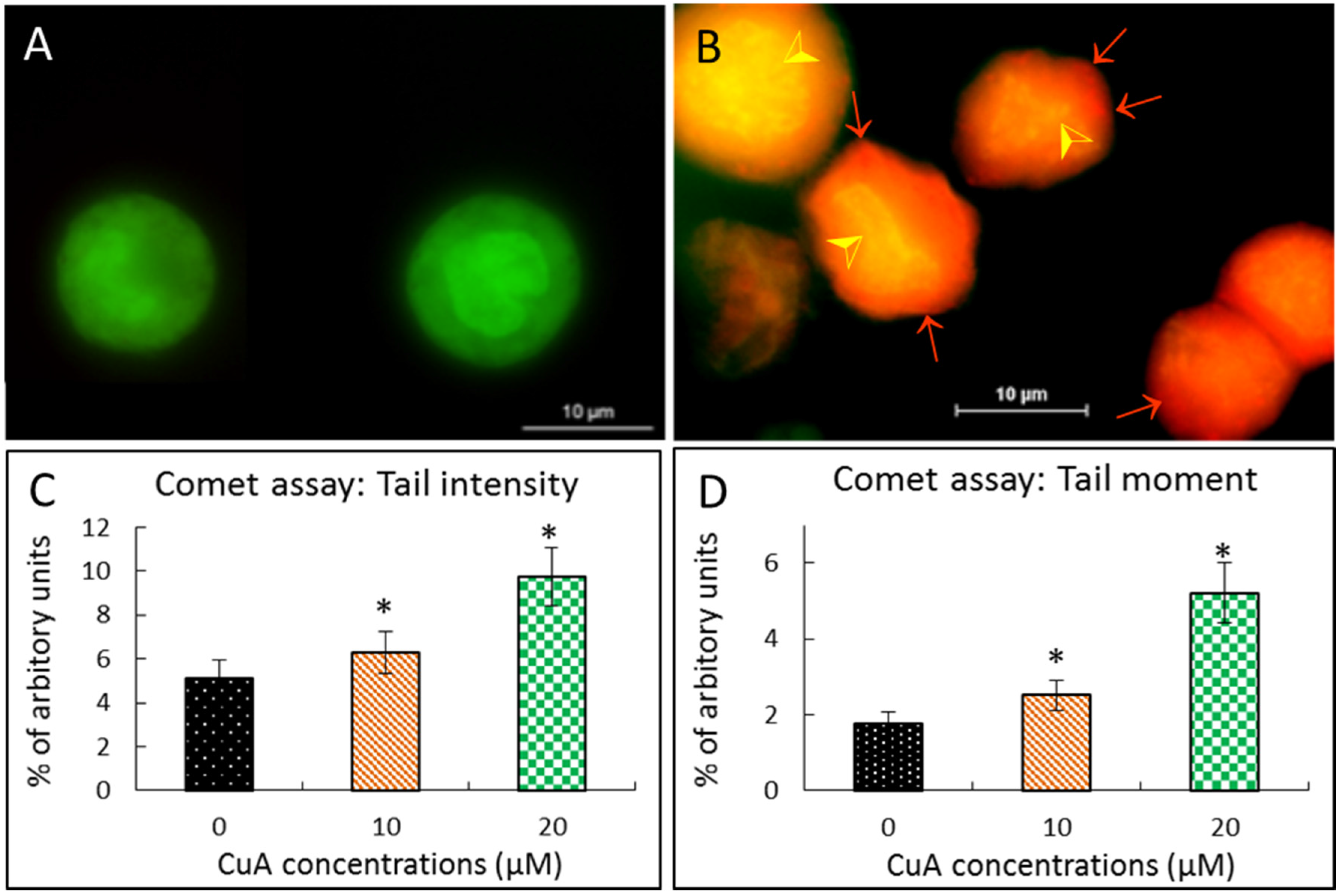
3.4. Cuminaldehyde Caused Nuclear Fragmentation in Human Colorectal COLO 205 Cells
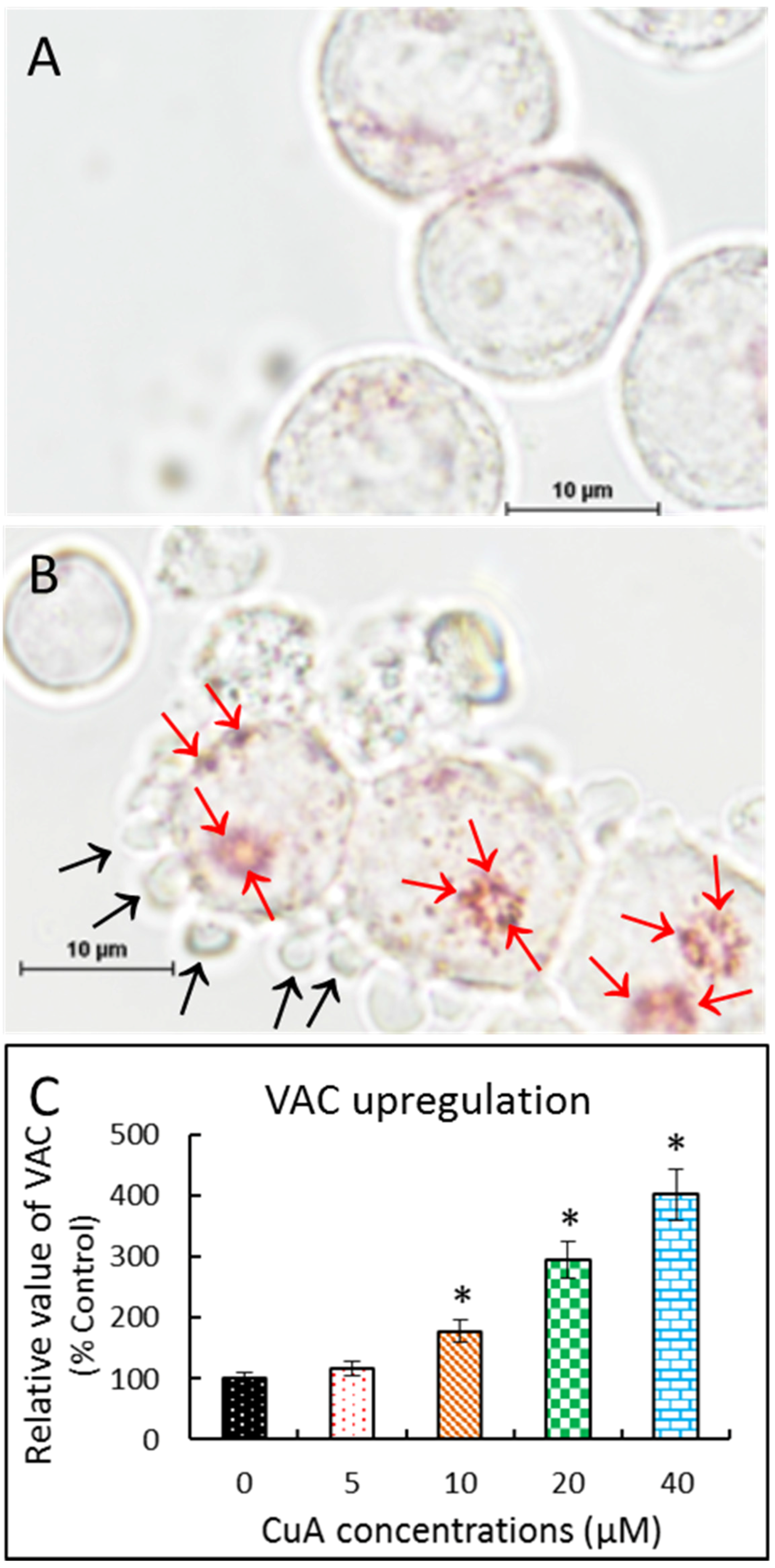
3.5. Cuminaldehyde Increased Volume of Acidic Compartment in Human Colorectal COLO 205 Cells
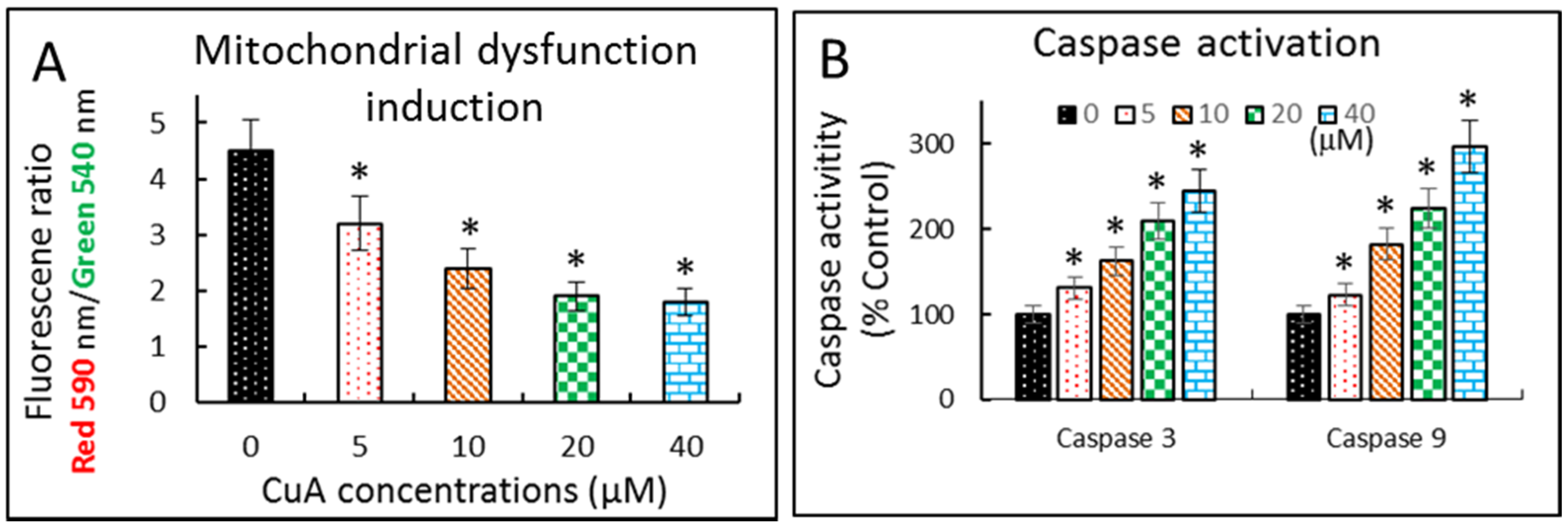
3.6. Cuminaldehyde Caused Apoptosis via the Mitochondrial Pathway in Human Colorectal COLO 205 Cells
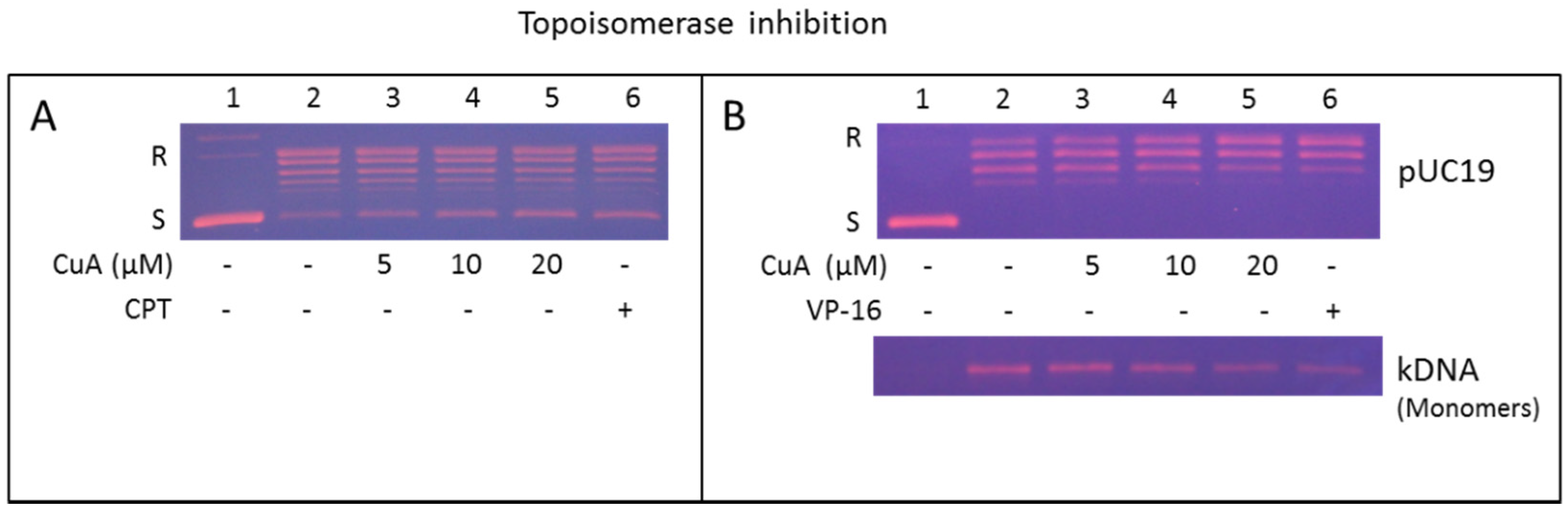
3.7. Cuminaldehyde Suppressed Topoisomerase I Activity in Human Colorectal COLO 205 Cells
3.8. Cuminaldehyde Suppressed Activity of Topoisomerase II in Human Colorectal COLO 205 Cells
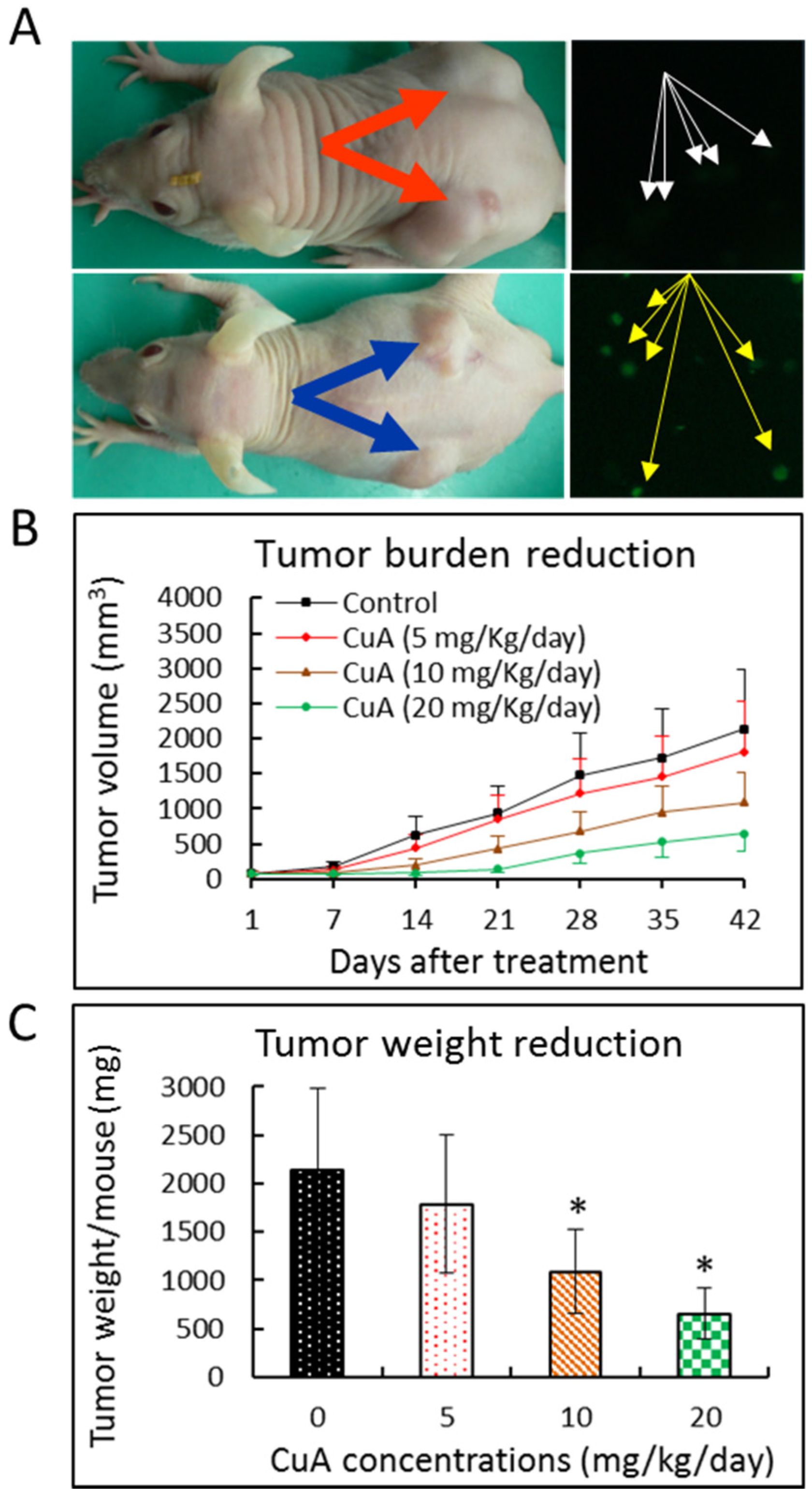
3.9. Cuminaldehyde Suppressed Growth of Human Colorectal COLO 205 Xenograft in a Nude Mice Model
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tanzer, M.; Liebl, M.; Quante, M. Molecular biomarkers in esophageal, gastric, and colorectal adenocarcinoma. Pharmacol. Ther. 2013, 140, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Yoon, Y.H.; Fukui, H.; Tabata, M.; Akira, T.; Okano, K.; Iwai, M.; Iga, Y.; Yokoyama, K. Antiulcerogenic compounds isolated from chinese cinnamon. Planta Medica 1989, 55, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.M.; Seo, J.H.; Ryu, S.Y.; Kim, Y.S.; Kim, Y.S.; Min, K.R.; Kim, Y. Cinnamaldehyde and 2-methoxycinnamaldehyde as NF-κB inhibitors from Cinnamomum cassia. Planta Medica 2004, 70, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Meeran, S.M.; Katiyar, S.K. Epigenetic regulation by selected dietary phytochemicals in cancer chemoprevention. Cancer Lett. 2014, 355, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, R.V.; Nagini, S. Cancer chemoprevention by dietary phytochemicals: Promises and pitfalls. Curr. Pharm. Biotechnol. 2012, 13, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Landau, J.M.; Huang, M.T.; Newmark, H.L. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [PubMed]
- Watson, W.H.; Cai, J.; Jones, D.P. Diet and apoptosis. Annu. Rev. Nutr. 2000, 20, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Duke, J.A. Dr. Duke’s Phytochemical and Ethnobotanical Databases. Available online: https://phytochem.nal.usda.gov/phytochem/chemicals/show/6429?et=C (accessed on 21 May 2016).
- Artandi, S.E.; DePinho, R.A. Telomeres and telomerase in cancer. Carcinogenesis 2010, 31, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Aleo, E.; Henderson, C.J.; Fontanini, A.; Solazzo, B.; Brancolini, C. Identification of new compounds that trigger apoptosome-independent caspase activation and apoptosis. Cancer Res. 2006, 66, 9235–9244. [Google Scholar] [CrossRef] [PubMed]
- Pop, C.; Timmer, J.; Sperandio, S.; Salvesen, G.S. The apoptosome activates caspase-9 by dimerization. Mol. Cell 2006, 22, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Henzel, W.J.; Liu, X.; Lutschg, A.; Wang, X. Apaf-1, a human protein homologous to C. elegans CED-4, participates in cytochrome c-dependent activation of caspase-3. Cell 1997, 90, 405–413. [Google Scholar] [CrossRef]
- Li, P.; Nijhawan, D.; Budihardjo, I.; Srinivasula, S.M.; Ahmad, M.; Alnemri, E.S.; Wang, X. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997, 91, 479–489. [Google Scholar] [CrossRef]
- McClendon, A.K.; Osheroff, N. DNA topoisomerase II, genotoxicity, and cancer. Mutat. Res. 2007, 623, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Heck, M.M.; Earnshaw, W.C. Topoisomerase II: A specific marker for cell proliferation. J. Cell Biol. 1986, 103, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Husain, I.; Mohler, J.L.; Seigler, H.F.; Besterman, J.M. Elevation of topoisomerase I messenger RNA, protein, and catalytic activity in human tumors: Demonstration of tumor-type specificity and implications for cancer chemotherapy. Cancer Res. 1994, 54, 539–546. [Google Scholar] [PubMed]
- Negri, F.V.; Azzoni, C.; Bottarelli, L.; Campanini, N.; Mandolesi, A.; Wotherspoon, A.; Cunningham, D.; Scartozzi, M.; Cascinu, S.; Tinelli, C.; et al. Thymidylate synthase, topoisomerase-1 and microsatellite instability: Relationship with outcome in mucinous colorectal cancer treated with fluorouracil. Anticancer Res. 2013, 33, 4611–4617. [Google Scholar] [PubMed]
- Sonderstrup, I.M.; Nygard, S.B.; Poulsen, T.S.; Linnemann, D.; Stenvang, J.; Nielsen, H.J.; Bartek, J.; Brunner, N.; Norgaard, P.; Riis, L. Topoisomerase-1 and -2A gene copy numbers are elevated in mismatch repair-proficient colorectal cancers. Mol. Oncol. 2015, 9, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Azzoni, C.; Bottarelli, L.; Cecchini, S.; Ziccarelli, A.; Campanini, N.; Bordi, C.; Sarli, L.; Silini, E.M. Role of topoisomerase I and thymidylate synthase expression in sporadic colorectal cancer: Associations with clinicopathological and molecular features. Pathol. Res. Pract. 2014, 210, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Naowaratwattana, W.; De-Eknamkul, W.; De Mejia, E.G. Phenolic-containing organic extracts of mulberry (Morus alba L.) leaves inhibit HepG2 hepatoma cells through G2/M phase arrest, induction of apoptosis, and inhibition of topoisomerase IIα activity. J. Med. Food 2010, 13, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Baikar, S.; Malpathak, N. Secondary metabolites as DNA topoisomerase inhibitors: A new era towards designing of anticancer drugs. Pharmacogn. Rev. 2010, 4, 12–26. [Google Scholar] [PubMed]
- Bandele, O.J.; Clawson, S.J.; Osheroff, N. Dietary polyphenols as topoisomerase II poisons: B ring and C ring substituents determine the mechanism of enzyme-mediated DNA cleavage enhancement. Chem. Res. Toxicol. 2008, 21, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Sudan, S.; Rupasinghe, H.P. Flavonoid-enriched apple fraction AF4 induces cell cycle arrest, DNA topoisomerase II inhibition, and apoptosis in human liver cancer HepG2 cells. Nutr. Cancer 2014, 66, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- El-Awady, R.A.; Ali, M.M.; Saleh, E.M.; Ghaleb, F.M. Apoptosis is the most efficient death-pathway in tumor cells after topoisomerase II inhibition. Saudi Med. J. 2008, 29, 558–564. [Google Scholar] [PubMed]
- Fan, C.; Wang, W.; Zhao, B.; Zhang, S.; Miao, J. Chloroquine inhibits cell growth and induces cell death in A549 lung cancer cells. Bioorg. Med. Chem. 2006, 14, 3218–3222. [Google Scholar] [CrossRef] [PubMed]
- Olive, P.L.; Banath, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Wang, X.; Han, J. Resistance to tumor necrosis factor-induced cell death mediated by PMCA4 deficiency. Mol. Cell Biol. 2001, 21, 8276–8288. [Google Scholar] [CrossRef] [PubMed]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (Δψm) in apoptosis; an update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Reers, M.; Smiley, S.T.; Mottola-Hartshorn, C.; Chen, A.; Lin, M.; Chen, L.B. Mitochondrial membrane potential monitored by JC-1 dye. Methods Enzymol. 1995, 260, 406–417. [Google Scholar] [PubMed]
- Martin, E.J.; Forkert, P.G. Evidence that 1,1-dichloroethylene induces apoptotic cell death in murine liver. J. Pharmacol. Exp. Ther. 2004, 310, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Fesik, S.W.; Shi, Y. Controlling the caspases. Science 2001, 294, 1477–1478. [Google Scholar] [CrossRef] [PubMed]
- Deveraux, Q.L.; Reed, J.C. Iap family proteins—Suppressors of apoptosis. Genes Dev. 1999, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Har-Vardi, I.; Mali, R.; Breietman, M.; Sonin, Y.; Albotiano, S.; Levitas, E.; Potashnik, G.; Priel, E. DNA topoisomerases I and II in human mature sperm cells: Characterization and unique properties. Hum. Reprod. 2007, 22, 2183–2189. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.; Crisol, L.; Prado, R.; Boyano, M.D.; Arluzea, J.; Arechaga, J. Plasma membrane and nuclear envelope integrity during the blebbing stage of apoptosis: A time-lapse study. Biol. Cell 2010, 102, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Lobner, D. Comparison of the LDH and MTT assays for quantifying cell death: Validity for neuronal apoptosis? J. Neurosci. Methods 2000, 96, 147–152. [Google Scholar] [CrossRef]
- Koh, J.Y.; Choi, D.W. Quantitative determination of glutamate mediated cortical neuronal injury in cell culture by lactate dehydrogenase efflux assay. J. Neurosci. Methods 1987, 20, 83–90. [Google Scholar] [CrossRef]
- Gwag, B.J.; Lobner, D.; Koh, J.Y.; Wie, M.B.; Choi, D.W. Blockade of glutamate receptors unmasks neuronal apoptosis after oxygen-glucose deprivation in vitro. Neuroscience 1995, 68, 615–619. [Google Scholar] [CrossRef]
- Koh, J.Y.; Gwag, B.J.; Lobner, D.; Choi, D.W. Potentiated necrosis of cultured cortical neurons by neurotrophins. Science 1995, 268, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J. Inhibition of apoptosis by ginsenoside RG1 in cultured cortical neurons. Chin. Med. J. (Engl.) 1997, 110, 535–539. [Google Scholar] [PubMed]
- Miura, T.; Chiba, M.; Kasai, K.; Nozaka, H.; Nakamura, T.; Shoji, T.; Kanda, T.; Ohtake, Y.; Sato, T. Apple procyanidins induce tumor cell apoptosis through mitochondrial pathway activation of caspase-3. Carcinogenesis 2008, 29, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.R.; Dong, H.W.; Chen, B.Q.; Zhao, P.; Liu, R.H. Fresh apples suppress mammary carcinogenesis and proliferative activity and induce apoptosis in mammary tumors of the sprague-dawley rat. J. Agric. Food Chem. 2009, 57, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Liu, R.H. Effect of selected phytochemicals and apple extracts on NF-κB activation in human breast cancer MCF-7 cells. J. Agric. Food Chem. 2007, 55, 3167–3173. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.Q.; Qiao, B.; Wang, M.; Tao, Q. Mechanisms of apple polyphenols-induced proliferation inhibiting and apoptosis in a metastatic oral adenoid cystic carcinoma cell line. Kaohsiung J. Med. Sci. 2013, 29, 239–245. [Google Scholar] [CrossRef] [PubMed]
- White, K.; Grether, M.E.; Abrams, J.M.; Young, L.; Farrell, K.; Steller, H. Genetic control of programmed cell death in drosophila. Science 1994, 264, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, Z. Differential staining of DNA and RNA in intact cells and isolated cell nuclei with acridine orange. Methods Cell Biol. 1990, 33, 285–298. [Google Scholar] [PubMed]
- Wyllie, A.H.; Kerr, J.F.; Currie, A.R. Cell death: The significance of apoptosis. Int. Rev. Cytol. 1980, 68, 251–306. [Google Scholar] [PubMed]
- Cover, T.L.; Puryear, W.; Perez-Perez, G.I.; Blaser, M.J. Effect of urease on HeLa cell vacuolation induced by Helicobacter pylori cytotoxin. Infect. Immun. 1991, 59, 1264–1270. [Google Scholar] [PubMed]
- Patel, H.K.; Willhite, D.C.; Patel, R.M.; Ye, D.; Williams, C.L.; Torres, E.M.; Marty, K.B.; MacDonald, R.A.; Blanke, S.R. Plasma membrane cholesterol modulates cellular vacuolation induced by the Helicobacter pylori vacuolating cytotoxin. Infect. Immun. 2002, 70, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y. Diversity of DNA topoisomerases I and inhibitors. Biochimie 1998, 80, 255–270. [Google Scholar] [CrossRef]
- Li, T.K.; Liu, L.F. Tumor cell death induced by topoisomerase-targeting drugs. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 53–77. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K. Role of spices beyond food flavoring: Nutraceuticals with multiple health effects. Food Rev. Int. 2005, 21, 167–188. [Google Scholar] [CrossRef]
- Duke, J.A.; Duke, P.-A.K.; duCellier, J.L. Duke’s Handbook of Medicinal Plants of the Bible; CRC Press: Boca Raton, NY, USA, 2008. [Google Scholar]
- Nitoda, T.; Fan, M.D.; Kubo, I. Effects of cuminaldehyde on melanoma cells. Phytother. Res. 2008, 22, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Tomy, M.J.; Dileep, K.V.; Prasanth, S.; Preethidan, D.S.; Sabu, A.; Sadasivan, C.; Haridas, M. Cuminaldehyde as a lipoxygenase inhibitor: In vitro and in silico validation. Appl. Biochem. Biotechnol. 2014, 174, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S. Cuminaldehyde: Aldose reductase and α-glucosidase inhibitor derived from Cuminum cyminum L. seeds. J. Agric. Food Chem. 2005, 53, 2446–2450. [Google Scholar] [CrossRef] [PubMed]
- Morshedi, D.; Aliakbari, F.; Tayaranian-Marvian, A.; Fassihi, A.; Pan-Montojo, F.; Perez-Sanchez, H. Cuminaldehyde as the major component of Cuminum cyminum, a natural aldehyde with inhibitory effect on alpha-synuclein fibrillation and cytotoxicity. J. Food Sci. 2015, 80, 2336–2345. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.B.; Takalikar, S.S.; Joglekar, M.M.; Haldavnekar, V.S.; Arvindekar, A.U. Insulinotropic and β-cell protective action of cuminaldehyde, cuminol and an inhibitor isolated from Cuminum cyminum in streptozotocin-induced diabetic rats. Br. J. Nutr. 2013, 110, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Aruna, K.; Sivaramakrishnan, V.M. Anticarcinogenic effects of the essential oils from cumin, poppy and basil. Phytother. Res. 1996, 10, 577–580. [Google Scholar] [CrossRef]
- Allahghadri, T.; Rasooli, I.; Owlia, P.; Nadooshan, M.J.; Ghazanfari, T.; Taghizadeh, M.; Astaneh, S.D. Antimicrobial property, antioxidant capacity, and cytotoxicity of essential oil from cumin produced in Iran. J. Food Sci. 2010, 75, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Hu, X.; Li, J.; Liu, P.; Yang, Y.; Ni, Y. Preparative isolation and purification of cuminaldehyde and p-menta-1,4-dien-7-al from the essential oil of Cuminum cyminum L. by high-speed counter-current chromatography. Anal. Chim. Acta 2011, 689, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, E.L.; Osheroff, N. Etoposide, topoisomerase II and cancer. Curr. Med. Chem. Anticancer Agents 2005, 5, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, D.C.; Chalmers, A.J.; El-Khamisy, S.F. Topoisomerase I inhibition in colorectal cancer: Biomarkers and therapeutic targets. Br. J. Cancer 2012, 106, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.H.; Christensen, I.J.; Jensen, N.F.; Markussen, B.; Romer, M.U.; Nygard, S.B.; Muller, S.; Nielsen, H.J.; Brunner, N.; Nielsen, K.V. Mechanisms of topoisomerase I (TOP1) gene copy number increase in a stage III colorectal cancer patient cohort. PLoS ONE 2013, 8, e60613. [Google Scholar] [CrossRef] [PubMed]
- Denny, W.A.; Baguley, B.C. Dual topoisomerase I/II inhibitors in cancer therapy. Curr. Top. Med. Chem. 2003, 3, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Bisceglie, F.; Pinelli, S.; Alinovi, R.; Goldoni, M.; Mutti, A.; Camerini, A.; Piola, L.; Tarasconi, P.; Pelosi, G. Cinnamaldehyde and cuminaldehyde thiosemicarbazones and their copper(II) and nickel(II) complexes: A study to understand their biological activity. J. Inorg. Biochem. 2014, 140, 111–125. [Google Scholar] [CrossRef] [PubMed]






© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, K.-d.; Liu, Y.-H.; Chen, T.-W.; Yang, S.-M.; Wong, H.-Y.; Cherng, J.; Chou, K.-S.; Cherng, J.-M. Cuminaldehyde from Cinnamomum verum Induces Cell Death through Targeting Topoisomerase 1 and 2 in Human Colorectal Adenocarcinoma COLO 205 Cells. Nutrients 2016, 8, 318. https://doi.org/10.3390/nu8060318
Tsai K-d, Liu Y-H, Chen T-W, Yang S-M, Wong H-Y, Cherng J, Chou K-S, Cherng J-M. Cuminaldehyde from Cinnamomum verum Induces Cell Death through Targeting Topoisomerase 1 and 2 in Human Colorectal Adenocarcinoma COLO 205 Cells. Nutrients. 2016; 8(6):318. https://doi.org/10.3390/nu8060318
Chicago/Turabian StyleTsai, Kuen-daw, Yi-Heng Liu, Ta-Wei Chen, Shu-Mei Yang, Ho-Yiu Wong, Jonathan Cherng, Kuo-Shen Chou, and Jaw-Ming Cherng. 2016. "Cuminaldehyde from Cinnamomum verum Induces Cell Death through Targeting Topoisomerase 1 and 2 in Human Colorectal Adenocarcinoma COLO 205 Cells" Nutrients 8, no. 6: 318. https://doi.org/10.3390/nu8060318