Association between Metabolite Profiles, Metabolic Syndrome and Obesity Status

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Anthropometric Measurements
2.3. Biochemical Parameters
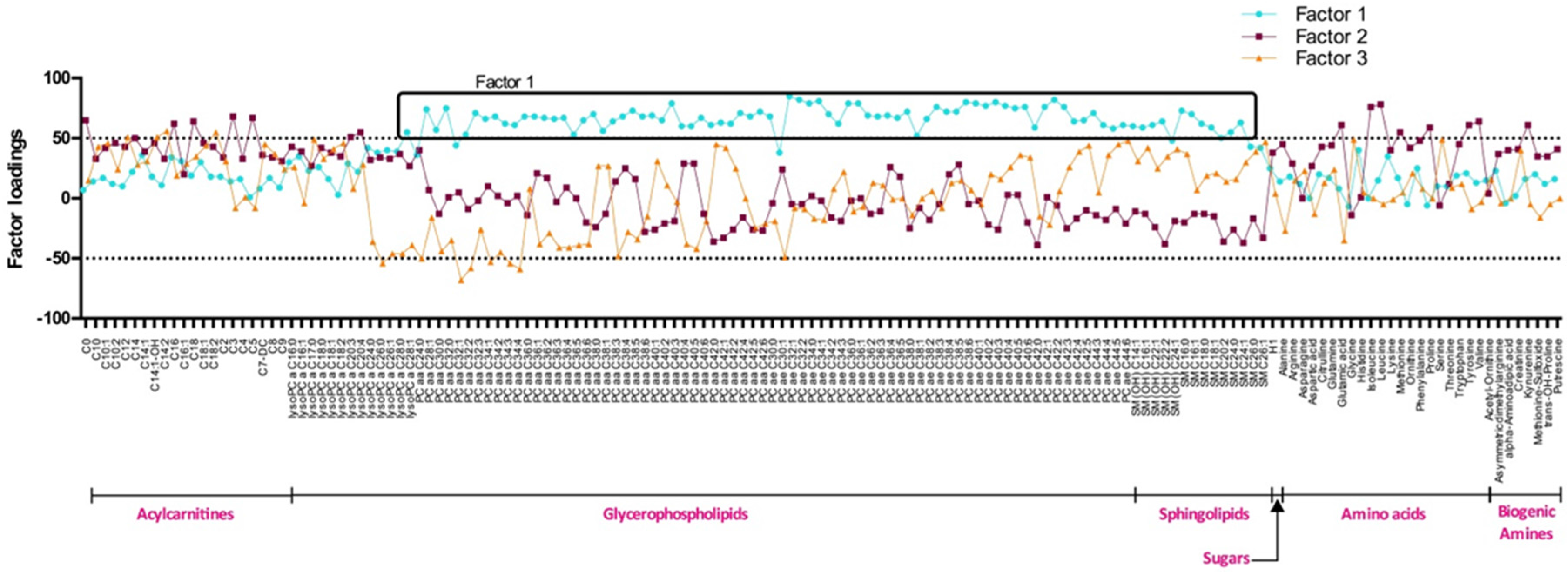
2.4. Metabolite Profiling
2.5. Statistical Analyses
3. Results
3.1. Descriptive Characteristics
3.2. Serum Metabolite Score
3.3. Relation between Cardiometabolic Risk Factors and PCA Factors
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Expert Panel on Detection, Evaluation. Executive summary of the third report of the national cholesterol education program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (Adult Treatment Panel III). JAMA 2001, 285, 2486–2497. [Google Scholar]
- Sowers, J.R. Obesity as a cardiovascular risk factor. Am. J. Med. 2003, 115, S37–S41. [Google Scholar] [CrossRef]
- James, P.T. Obesity: The worldwide epidemic. Clin. Dermatol. 2004, 22, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Karelis, A.D.; St-Pierre, D.H.; Conus, F.; Rabasa-Lhoret, R.; Poehlman, E.T. Metabolic and body composition factors in subgroups of obesity: What do we know? J. Clin. Endocrinol. Metab. 2004, 89, 2569–2575. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Salinas, C.A.; Garcia, E.G.; Robles, L.; Riano, D.; Ruiz-Gomez, D.G.; Garcia-Ulloa, A.C.; Melgarejo, M.A.; Zamora, M.; Guillen-Pineda, L.E.; Mehta, R.S.; et al. High adiponectin concentrations are associated with the metabolically healthy obese phenotype. J. Clin. Endocrinol. Metab. 2008, 93, 4075–4079. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Stamatakis, E. Metabolically healthy obesity and risk of all-cause and cardiovascular disease mortality. J. Clin. Endocrinol. Metab. 2012, 97, 2482–2488. [Google Scholar] [CrossRef] [PubMed]
- Wildman, R.P.; Muntner, P.; Reynolds, K.; McGinn, A.P.; Rajpathak, S.; Wylie-Rosett, J.; Sowers, M.R. The obese without cardiometabolic risk factor clustering and the normal weight with cardiometabolic risk factor clustering: Prevalence and correlates of 2 phenotypes among the US population (NHANES 1999–2004). Arch. Intern. Med. 2008, 168, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.R.; Stevens, R.D.; Wenner, B.R.; Ilkayeva, O.; Muoio, D.M.; Newgard, C.B. Metabolomics applied to diabetes research: Moving from information to knowledge. Diabetes 2009, 58, 2429–2443. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Jang, H.B.; Ra, M.; Choi, Y.; Lee, H.J.; Park, J.Y.; Kang, J.H.; Park, K.H.; Park, S.I.; Song, J. Prediction of future risk of insulin resistance and metabolic syndrome based on Korean boy’s metabolite profiling. Obes. Res. Clin. Pract. 2014, 9, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Jourdan, C.; Petersen, A.K.; Gieger, C.; Doring, A.; Illig, T.; Wang-Sattler, R.; Meisinger, C.; Peters, A.; Adamski, J.; Prehn, C.; et al. Body fat free mass is associated with the serum metabolite profile in a population-based study. PLoS ONE 2012, 7, e40009. [Google Scholar] [CrossRef] [PubMed]
- Paradis, A.M.; Perusse, L.; Godin, G.; Vohl, M.C. Validity of a self-reported measure of familial history of obesity. Nutr. J. 2008, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Frankenfield, D.C.; Rowe, W.A.; Cooney, R.N.; Smith, J.S.; Becker, D. Limits of body mass index to detect obesity and predict body composition. Nutrition 2001, 17, 26–30. [Google Scholar] [CrossRef]
- Albers, J.J.; Warnick, G.R.; Wiebe, D.; King, P.; Steiner, P.; Smith, L.; Breckenridge, C.; Chow, A.; Kuba, K.; Weidman, S.; et al. Multi-laboratory comparison of three heparin-Mn2+ precipitation procedures for estimating cholesterol in high-density lipoprotein. Clin. Chem. 1978, 24, 853–856. [Google Scholar] [PubMed]
- Kubo, N. Evaluation of LDL-cholesterol measurement. Rinsho Byori 2002, 50, 242–247. [Google Scholar] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Desbuquois, B.; Aurbach, G.D. Use of polyethylene glycol to separate free and antibody-bound peptide hormones in radioimmunoassays. J. Clin. Endocrinol. Metab. 1971, 33, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Richterich, R.; Kuffer, H.; Lorenz, E.; Colombo, J.P. The determination of glucose in plasma and serum (hexokinase-glucose-6-phosphate dehydrogenase method) with the Greiner electronic selective analyzer GSA II (author’s transl). Z. Klin. Chem. Klin. Biochem. 1974, 12, 5–13. [Google Scholar] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Despres, J.P.; Lemieux, I.; Bergeron, J.; Pibarot, P.; Mathieu, P.; Larose, E.; Rodes-Cabau, J.; Bertrand, O.F.; Poirier, P. Abdominal obesity and the metabolic syndrome: Contribution to global cardiometabolic risk. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Real, J.M.; Menendez, J.A.; Moreno-Navarrete, J.M.; Bluher, M.; Vazquez-Martin, A.; Vazquez, M.J.; Ortega, F.; Dieguez, C.; Fruhbeck, G.; Ricart, W.; et al. Extracellular fatty acid synthase: A possible surrogate biomarker of insulin resistance. Diabetes 2010, 59, 1506–1511. [Google Scholar] [CrossRef] [PubMed]
- Andreoli, A.; Garaci, F.; Cafarelli, F.P.; Guglielmi, G. Body composition in clinical practice. Eur. J. Radiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Heilbronn, L.K.; Coster, A.C.; Campbell, L.V.; Greenfield, J.R.; Lange, K.; Christopher, M.J.; Meikle, P.J.; Samocha-Bonet, D. The effect of short-term overfeeding on serum lipids in healthy humans. Obesity 2013, 21, E649–E659. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, K.; Krautbauer, S.; Hebel, T.; Schmitz, G.; Aslanidis, C.; Liebisch, G.; Buechler, C. Lipidomic analysis of the liver from high-fat diet induced obese mice identifies changes in multiple lipid classes. Exp. Mol. Pathol. 2014, 97, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, B. Plasmalogens: Targets for oxidants and major lipophilic antioxidants. Biochem. Soc. Trans. 2004, 32, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta 2012, 1822, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Kelly, A.S.; Jacobs, D.R.; Sinaiko, A.R., Jr.; Moran, A.; Steffen, L.M.; Steinberger, J. Relation of circulating oxidized LDL to obesity and insulin resistance in children. Pediatr. Diabetes 2010, 11, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Pietilainen, K.H.; Sysi-Aho, M.; Rissanen, A.; Seppanen-Laakso, T.; Yki-Jarvinen, H.; Kaprio, J.; Oresic, M. Acquired obesity is associated with changes in the serum lipidomic profile independent of genetic effects—A monozygotic twin study. PLoS ONE 2007, 2, e218. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Tseng, Y.J.; Wang, S.Y.; Tsai, Y.S.; Chang, C.S.; Kuo, T.C.; Yao, W.J.; Shieh, C.C.; Wu, C.H.; Kuo, P.H. The metabolome profiling and pathway analysis in metabolic healthy and abnormal obesity. Int. J. Obes. 2015, 39, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Yu, Z.; Kleber, M.; Singmann, P.; Holzapfel, C.; He, Y.; Mittelstrass, K.; Polonikov, A.; Prehn, C.; Romisch-Margl, W.; et al. Childhood obesity is associated with changes in the serum metabolite profile. Obes. Facts 2012, 5, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Shah, S.H.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.; Slentz, C.A.; Tanner, C.J.; Kuchibhatla, M.; Houmard, J.A.; Newgard, C.B.; et al. Relationships between circulating metabolic intermediates and insulin action in overweight to obese, inactive men and women. Diabetes Care 2009, 32, 1678–1683. [Google Scholar] [CrossRef] [PubMed]
- Oberbach, A.; Bluher, M.; Wirth, H.; Till, H.; Kovacs, P.; Kullnick, Y.; Schlichting, N.; Tomm, J.M.; Rolle-Kampczyk, U.; Murugaiyan, J.; et al. Combined proteomic and metabolomic profiling of serum reveals association of the complement system with obesity and identifies novel markers of body fat mass changes. J. Proteom. Res. 2011, 10, 4769–4788. [Google Scholar] [CrossRef] [PubMed]
- Layman, D.K. The role of leucine in weight loss diets and glucose homeostasis. J. Nutr. 2003, 133, S261–S267. [Google Scholar]
- Cota, D.; Proulx, K.; Smith, K.A.; Kozma, S.C.; Thomas, G.; Woods, S.C.; Seeley, R.J. Hypothalamic mTOR signaling regulates food intake. Science 2006, 312, 927–930. [Google Scholar] [CrossRef] [PubMed]
- Fajans, S.S. Leucine-Induced hypoglycemia. N. Engl. J. Med. 1965, 272, 1224–1227. [Google Scholar] [CrossRef] [PubMed]
- Hay, N.; Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.H.; Crosslin, D.R.; Haynes, C.S.; Nelson, S.; Turer, C.B.; Stevens, R.D.; Muehlbauer, M.J.; Wenner, B.R.; Bain, J.R.; Laferrere, B.; et al. Branched-chain amino acid levels are associated with improvement in insulin resistance with weight loss. Diabetologia 2012, 55, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.W.; Meigs, J.B.; Sullivan, L.; Fox, C.S.; Nathan, D.M.; D’Agostino, R.B., Sr. Prediction of incident diabetes mellitus in middle-aged adults: The Framingham Offspring Study. Arch. Intern. Med. 2007, 167, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite profiles and the risk of developing diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Superko, H.R.; Krauss, R.M. Coronary artery disease regression. Convincing evidence for the benefit of aggressive lipoprotein management. Circulation 1994, 90, 1056–1069. [Google Scholar] [CrossRef] [PubMed]
- Graessler, J.; Bornstein, T.D.; Goel, D.; Bhalla, V.P.; Lohmann, T.; Wolf, T.; Koch, M.; Qin, Y.; Licinio, J.; Wong, M.L.; et al. Lipidomic profiling before and after Roux-en-Y gastric bypass in obese patients with diabetes. Pharmacogenom. J. 2014, 14, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Doi, M.; Yamaoka, I.; Nakayama, M.; Mochizuki, S.; Sugahara, K.; Yoshizawa, F. Isoleucine, a blood glucose-lowering amino acid, increases glucose uptake in rat skeletal muscle in the absence of increases in AMP-activated protein kinase activity. J. Nutr. 2005, 135, 2103–2108. [Google Scholar] [PubMed]
- Doi, M.; Yamaoka, I.; Nakayama, M.; Sugahara, K.; Yoshizawa, F. Hypoglycemic effect of isoleucine involves increased muscle glucose uptake and whole body glucose oxidation and decreased hepatic gluconeogenesis. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1683–E1693. [Google Scholar] [CrossRef] [PubMed]
- Bouchard-Mercier, A.; Rudkowska, I.; Lemieux, S.; Couture, P.; Vohl, M.C. The metabolic signature associated with the Western dietary pattern: A cross-sectional study. Nutr. J. 2013, 12, 158. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| MetS− | MetS+ | p-Value | ||
|---|---|---|---|---|
| Ov/Ob (n = 83) | NW (n = 65) | Ov/Ob (n = 46) | ||
| Age (years) | 35.7 ± 10.4 2 | 28.9 ± 7.4 1 | 37.9 ± 10.0 3 | <0.0001 |
| BMI (kg/m2) | 31.4 ± 4.2 2 | 22.2 ± 1.8 1 | 34.3 ± 4.9 3 | <0.0001 |
| Waist circumference (cm) a | 97.8 ± 10.8 2 | 74.7 ± 5.7 1 | 109.0 ± 11.9 3 | <0.0001 |
| Fat mass (kg) | 31.3 ± 9.0 2 | 14.5 ± 4.3 1 | 36.5 ± 10.5 3 | <0.0001 |
| Lean mass (kg) | 57.5 ± 11.1 2 | 48.9 ± 8.9 1 | 65.3 ± 12.1 3 | <0.0001 |
| Systolic blood pressure (mmHg) | 119.9 ± 9.0 2 | 115.9 ± 9.8 1 | 130.1 ± 10.6 3 | <0.0001 |
| Diastolic blood pressure (mmHg) | 77.7 ± 7.7 1 | 74.0 ± 9.9 1 | 83.5 ± 9.5 2 | <0.0001 |
| Fasting glucose (mmol/L) | 5.4 ± 0.5 2 | 5.8 ± 1.3 1 | 6.1 ± 1.1 3 | 0.002 |
| Fasting insulin (pmol/L) | 86.1 ± 56.3 2 | 48.7 ± 17.1 1 | 133.1 ± 72.3 3 | <0.0001 |
| HOMA-IR | 2.98 ± 1.77 2 | 1.84 ± 0.94 1 | 5.33 ± 3.70 3 | <0.0001 |
| Total-C (mmol/L) | 4.57 ± 1.00 2 | 4.13 ± 0.67 1 | 4.84 ± 1.14 2 | 0.02 |
| LDL-C (mmol/L) | 2.81 ± 0.91 | 2.52 ± 0.70 | 2.99 ± 1.23 | 0.29 |
| HDL-C (mmol/L) | 1.32 ± 0.30 2 | 1.60 ± 0.45 1 | 0.99 ± 0.24 3 | <0.0001 |
| TG (mmol/L) | 1.15 ± 0.56 2 | 0.77 ± 0.31 1 | 2.16 ± 1.21 3 | <0.0001 |
| MetS− | MetS+ | p-Value | ||
|---|---|---|---|---|
| Ov/Ob (n = 83) | NW (n = 65) | Ov/Ob (n = 46) | ||
| Factor 1 | 0.01 ± 1.01 2 | −0.31 ± 0.84 3 | 0.36 ± 1.01 1 | 0.003 |
| Factor 2 | −0.05 ± 1.04 2 | 0.14 ± 1.03 3 | −0.22 ± 0.73 1 | 0.02 |
| Factor 3 | −0.01 ± 0.99 2 | 0.34 ± 0.95 3 | −0.36 ± 0.95 1 | 0.03 |
| MetS− | MetS+ | p-Value | ||
|---|---|---|---|---|
| Ob (n = 84) | NW (n = 65) | Ob (n = 36) | ||
| Factor 1 | 0.03 ± 0.11 3 | −0.35 ± 0.14 2 | 0.30 ± 0.16 1 | 0.005 |
| Factor 2 | −0.20 ± 0.11 1 | 0.29 ± 0.14 2 | −0.17 ± 0.16 1 | 0.03 |
| Factor 3 | −0.08 ± 0.12 1 | 0.12 ± 0.15 1 | 0.03 ± 0.17 1 | 0.59 |
| MetS− | MetS+ | |||
|---|---|---|---|---|
| Ov/Ob (n = 83) | NW (n = 65) | Ov/Ob (n = 46) | ||
| WC | r | 0.14 | 0.23 | 0.01 |
| p | 0.23 | 0.07 | 0.95 | |
| Total-C | r | 0.32 | 0.38 | 0.63 |
| p | 0.003 | 0.003 | <0.0001 | |
| TG | r | 0.61 | 0.45 | 0.44 |
| p | <0.0001 | 0.0002 | 0.003 | |
| HDL-C | r | 0.27 | 0.26 | 0.19 |
| p | 0.01 | 0.04 | 0.22 | |
| LDL-C | r | 0.11 | 0.22 | 0.31 |
| p | 0.33 | 0.09 | 0.04 | |
| Glucose | r | −0.03 | 0.03 | 0.16 |
| p | 0.81 | 0.81 | 0.32 | |
| Insulin | r | 0.12 | −0.05 | −0.03 |
| p | 0.39 | 0.69 | 0.86 | |
| SBP | r | 0.03 | −0.07 | 0.23 |
| p | 0.81 | 0.64 | 0.19 | |
| DBP | r | −0.1 | −0.04 | 0.16 |
| p | 0.39 | 0.74 | 0.29 | |
| HOMA-IR | r | 0.08 | −0.03 | 0.002 |
| p | 0.47 | 0.82 | 0.91 | |
| MetS− | MetS+ | |||
|---|---|---|---|---|
| Ov/Ob (n = 83) | NW (n = 65) | Ov/Ob (n = 46) | ||
| WC | r | −0.10 | 0.09 | −0.36 |
| p | 0.35 | 0.47 | 0.02 | |
| Total-C | r | 0.34 | 0.18 | 0.17 |
| p | 0.002 | 0.17 | 0.28 | |
| TG | r | 0.01 | −0.09 | 0.05 |
| p | 0.91 | 0.46 | 0.75 | |
| HDL-C | r | 0.09 | 0.34 | 0.12 |
| p | 0.42 | 0.008 | 0.53 | |
| LDL-C | r | 0.30 | −0.02 | 0.16 |
| p | 0.007 | 0.91 | 0.31 | |
| Glucose | r | −0.02 | 0.002 | 0.14 |
| p | 0.83 | 0.99 | 0.36 | |
| Insulin | r | 0.08 | −0.25 | 0.15 |
| p | 0.49 | 0.05 | 0.32 | |
| SBP | r | 0.05 | −0.04 | 0.09 |
| p | 0.67 | 0.79 | 0.58 | |
| DBP | r | 0.09 | 0.08 | 0.06 |
| p | 0.44 | 0.56 | 0.71 | |
| HOMA-IR | r | 0.07 | −0.14 | 0.16 |
| p | 0.54 | 0.30 | 0.30 | |
| MetS− | MetS+ | |||
|---|---|---|---|---|
| Ov/Ob (n = 83) | NW (n = 65) | Ov/Ob (n = 46) | ||
| WC | r | −0.07 | 0.02 | 0.17 |
| p | 0.55 | 0.89 | 0.28 | |
| Total-C | r | −0.06 | 0.14 | 0.13 |
| p | 0.62 | 0.29 | 0.41 | |
| TG | r | −0.16 | 0.04 | −0.22 |
| p | 0.16 | 0.75 | 0.15 | |
| HDL-C | r | 0.16 | 0.22 | 0.14 |
| p | 0.16 | 0.09 | 0.36 | |
| LDL-C | r | −0.08 | 0.01 | 0.11 |
| p | 0.46 | 0.93 | 0.48 | |
| Glucose | r | 0.0009 | −0.10 | −0.32 |
| p | 0.99 | 0.42 | 0.05 | |
| Insulin | r | −0.27 | −0.21 | −0.1 |
| p | 0.02 | 0.09 | 0.52 | |
| SBP | r | 0.07 | 0.06 | −0.06 |
| p | 0.51 | 0.67 | 0.71 | |
| DBP | r | −0.04 | 0.01 | 0.25 |
| p | 0.71 | 0.92 | 0.11 | |
| HOMA-IR | r | −0.24 | −0.23 | −0.17 |
| p | 0.03 | 0.08 | 0.26 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allam-Ndoul, B.; Guénard, F.; Garneau, V.; Cormier, H.; Barbier, O.; Pérusse, L.; Vohl, M.-C. Association between Metabolite Profiles, Metabolic Syndrome and Obesity Status. Nutrients 2016, 8, 324. https://doi.org/10.3390/nu8060324
Allam-Ndoul B, Guénard F, Garneau V, Cormier H, Barbier O, Pérusse L, Vohl M-C. Association between Metabolite Profiles, Metabolic Syndrome and Obesity Status. Nutrients. 2016; 8(6):324. https://doi.org/10.3390/nu8060324
Chicago/Turabian StyleAllam-Ndoul, Bénédicte, Frédéric Guénard, Véronique Garneau, Hubert Cormier, Olivier Barbier, Louis Pérusse, and Marie-Claude Vohl. 2016. "Association between Metabolite Profiles, Metabolic Syndrome and Obesity Status" Nutrients 8, no. 6: 324. https://doi.org/10.3390/nu8060324
APA StyleAllam-Ndoul, B., Guénard, F., Garneau, V., Cormier, H., Barbier, O., Pérusse, L., & Vohl, M.-C. (2016). Association between Metabolite Profiles, Metabolic Syndrome and Obesity Status. Nutrients, 8(6), 324. https://doi.org/10.3390/nu8060324