Leucine-Enriched Essential Amino Acids Augment Mixed Protein Synthesis, But Not Collagen Protein Synthesis, in Rat Skeletal Muscle after Downhill Running


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Leucine-Enriched Essential Amino Acids
2.4. Measurements of Blood Variables
2.5. Measurement of Mixed and Collagen Protein Synthesis
2.6. Statistical Analysis
3. Results
3.1. Intracellular Proline Enrichment in Vastus Lateralis
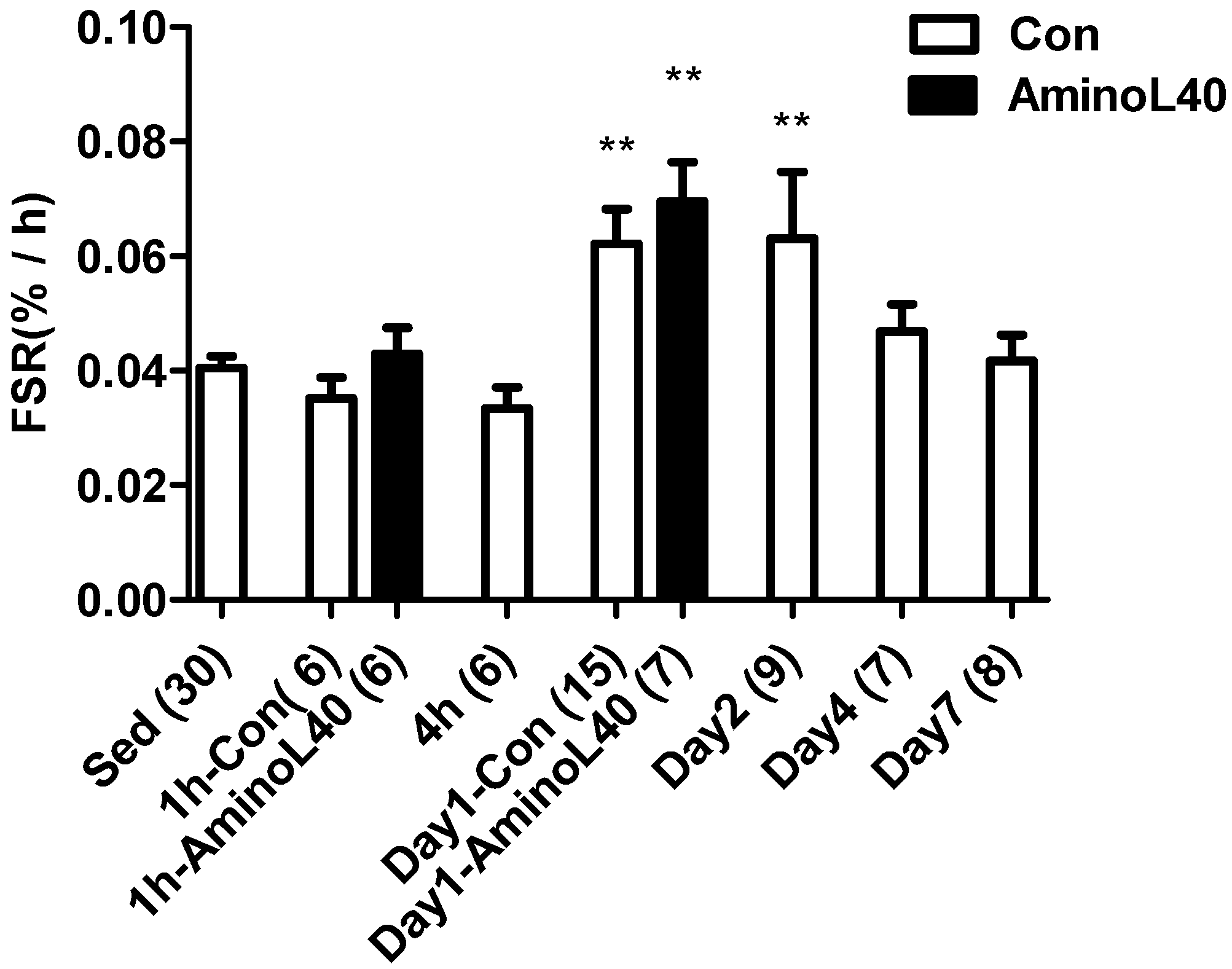
3.2. Mixed Protein Synthesis after Downhill Running Exercise
3.3. Collagen Protein Synthesis after Downhill Running Exercise
3.4. Amino Acid Concentrations after Administration of LEAAs Following Downhill Running Exercise
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| FSR | fractional synthesis rate |
| LEAAs | Leucine-enriched essential amino acids |
| MPE | mole per excess |
| mTOR | Mammalian target of rapamycin |
References
- Huijing, P.A. Muscle as a collagen fiber reinforced composite: A review of force transmission in muscle and whole limb. J. Biomech. 1999, 32, 329–345. [Google Scholar] [CrossRef]
- Kovanen, V.; Suominen, H.; Heikkinen, E. Connective tissue of “fast” and “slow” skeletal muscle in rats—Effects of endurance training. Acta Physiol. Scand. 1980, 108, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.E.; Goldspink, G. Connective tissue changes in surgically overloaded muscle. Cell Tissue Res. 1981, 221, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, M. Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 2004, 84, 649–698. [Google Scholar] [CrossRef] [PubMed]
- Stauber, W.T. Factors involved in strain-induced injury in skeletal muscles and outcomes of prolonged exposures. J. Electromyogr. Kinesiol. 2004, 14, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.R.; Phillips, S.M.; Babraj, J.A.; Smith, K.; Rennie, M.J. Myofibrillar and collagen protein synthesis in human skeletal muscle in young men after maximal shortening and lengthening contractions. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E1153–E1159. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.J.; Babraj, J.; Smith, K.; Wilkes, E.; Fedele, M.J.; Esser, K.; Rennie, M. Anabolic signaling and protein synthesis in human skeletal muscle after dynamic shortening or lengthening exercise. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E731–E738. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.F.; Olesen, J.L.; Hansen, M.; Dossing, S.; Crameri, R.M.; Welling, R.J.; Langberg, H.; Flyvbjerg, A.; Kjaer, M.; Babraj, J.A.; et al. Coordinated collagen and muscle protein synthesis in human patella tendon and quadriceps muscle after exercise. J. Physiol. 2005, 567, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.M.; Tipton, K.D.; Aarsland, A.; Wolf, S.E.; Wolfe, R.R. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. Am. J. Physiol. 1997, 273, E99–E107. [Google Scholar] [PubMed]
- Biolo, G.; Maggi, S.P.; Williams, B.D.; Tipton, K.D.; Wolfe, R.R. Increased rates of muscle protein turnover and amino acid transport after resistance exercise in humans. Am. J. Physiol. 1995, 268, E514–E520. [Google Scholar] [PubMed]
- Burd, N.A.; Tang, J.E.; Moore, D.R.; Phillips, S.M. Exercise training and protein metabolism: Influences of contraction, protein intake, and sex-based differences. J. Appl. Physiol. 2009, 106, 1692–1701. [Google Scholar] [CrossRef] [PubMed]
- Biolo, G.; Tipton, K.D.; Klein, S.; Wolfe, R.R. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am. J. Physiol. 1997, 273, E122–E129. [Google Scholar] [PubMed]
- Moore, D.R.; Robinson, M.J.; Fry, J.L.; Tang, J.E.; Glover, E.I.; Wilkinson, S.B.; Prior, T.; Tarnopolsky, M.A.; Phillips, S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am. J. Clin. Nutr. 2009, 89, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Babraj, J.A.; Cuthbertson, D.J.; Smith, K.; Langberg, H.; Miller, B.; Krogsgaard, M.R.; Kjaer, M.; Rennie, M.J. Collagen synthesis in human musculoskeletal tissues and skin. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E864–E869. [Google Scholar] [CrossRef] [PubMed]
- Mittendorfer, B.; Andersen, J.L.; Plomgaard, P.; Saltin, B.; Babraj, J.A.; Smith, K.; Rennie, M.J. Protein synthesis rates in human muscles: Neither anatomical location nor fibre-type composition are major determinants. J. Physiol. 2005, 563, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; van Hall, G.; Rose, A.J.; Miller, B.F.; Doessing, S.; Richter, E.A.; Kjaer, M. Contraction intensity and feeding affect collagen and myofibrillar protein synthesis rates differently in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E257–E269. [Google Scholar] [CrossRef] [PubMed]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. A high proportion of leucine is required for optimal stimulation of the rate of muscle protein synthesis by essential amino acids in the elderly. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E381–E387. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, H.C.; Drummond, M.J.; Pennings, B.; Fujita, S.; Glynn, E.L.; Chinkes, D.L.; Dhanani, S.; Volpi, E.; Rasmussen, B.B. Leucine-enriched essential amino acid and carbohydrate ingestion following resistance exercise enhances mtor signaling and protein synthesis in human muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E392–E400. [Google Scholar] [CrossRef] [PubMed]
- Fujita, S.; Dreyer, H.C.; Drummond, M.J.; Glynn, E.L.; Cadenas, J.G.; Yoshizawa, F.; Volpi, E.; Rasmussen, B.B. Nutrient signalling in the regulation of human muscle protein synthesis. J. Physiol. 2007, 582, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Rasmussen, B.B. Leucine-enriched nutrients and the regulation of mammalian target of rapamycin signalling and human skeletal muscle protein synthesis. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Suzuki, H.; Mimura, M.; Inoue, Y.; Sugita, M.; Suzuki, K.; Kobayashi, H. Leucine-enriched essential amino acids attenuate muscle soreness and improve muscle protein synthesis after eccentric contractions in rats. Amino Acids 2015, 47, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, D.S.; Nelson, A.R.; Raymond, F.; Metairon, S.; Mansourian, R.; Clarke, J.; Stellingwerff, T.; Phillips, S.M. Protein-Leucine Ingestion Activates a Regenerative Inflammo-Myogenic Transcriptome in Skeletal Muscle Following Intense Endurance Exercise. Physiol. Genom. 2016, 48, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Y.; Wang, W.; Komulainen, J.; Koskinen, S.O.; Kovanen, V.; Vihko, V.; Trackman, P.C.; Takala, T.E. Increased mRNAs for procollagens and key regulating enzymes in rat skeletal muscle following downhill running. Pflügers Archiv 1999, 437, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am. J. Clin. Nutr. 2014, 99, 86–95. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, R.J. Methods for measuring hydroxyproline and estimating in vivo rates of collagen synthesis and degradation. Methods Mol. Med. 2005, 117, 189–207. [Google Scholar] [PubMed]
- Laurent, G.J. Dynamic state of collagen: Pathways of collagen degradation in vivo and their possible role in regulation of collagen mass. Am. J. Physiol. 1987, 252, C1–C9. [Google Scholar] [PubMed]
- McAnulty, R.J.; Laurent, G.J. Collagen synthesis and degradation in vivo. Evidence for rapid rates of collagen turnover with extensive degradation of newly synthesized collagen in tissues of the adult rat. Coll. Relat. Res. 1987, 7, 93–104. [Google Scholar] [CrossRef]
- Laurent, G.J. Rates of collagen synthesis in lung, skin and muscle obtained in vivo by a simplified method using [3H] proline. Biochem. J. 1982, 206, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Rennie, M.J. New approaches and recent results concerning human-tissue collagen synthesis. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Dietzen, D.J.; Weindel, A.L.; Carayannopoulos, M.O.; Landt, M.; Normansell, E.T.; Reimschisel, T.E.; Smith, C.H. Rapid comprehensive amino acid analysis by liquid chromatography/tandem mass spectrometry: Comparison to cation exchange with post-column ninhydrin detection. Rapid Commun. Mass Spectrom. 2008, 22, 3481–3488. [Google Scholar] [CrossRef] [PubMed]
- Calder, A.G.; Anderson, S.E.; Grant, I.; McNurlan, M.A.; Garlick, P.J. The determination of low d5-phenylalanine enrichment (0.002–0.09 atom percent excess), after conversion to phenylethylamine, in relation to protein turnover studies by gas chromatography/electron ionization mass spectrometry. Rapid Commun. Mass Spectrom. 1992, 6, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Drummond, M.J.; Dreyer, H.C.; Fry, C.S.; Glynn, E.L.; Rasmussen, B.B. Nutritional and contractile regulation of human skeletal muscle protein synthesis and mtorc1 signaling. J. Appl. Physiol. 2009, 106, 1374–1384. [Google Scholar] [CrossRef] [PubMed]
- Crozier, S.J.; Kimball, S.R.; Emmert, S.W.; Anthony, J.C.; Jefferson, L.S. Oral leucine administration stimulates protein synthesis in rat skeletal muscle. J. Nutr. 2005, 135, 376–382. [Google Scholar] [PubMed]
- Peake, J.; Nosaka, K.; Suzuki, K. Characterization of inflammatory responses to eccentric exercise in humans. Exerc. Immunol. Rev. 2005, 11, 64–85. [Google Scholar] [PubMed]
- Duncan, M.R.; Berman, B. Stimulation of collagen and glycosaminoglycan production in cultured human adult dermal fibroblasts by recombinant human interleukin 6. J. Investig. Dermatol. 1991, 97, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Miura, K.; Nakano, S.; Suzuki, K.; Bannai, M.; Inoue, Y. Leucine-enriched essential amino acids attenuate inflammation in rat muscle and enhance muscle repair after eccentric contraction. Amino Acids 2016. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.G.; Silva, M.T.; Carlassara, E.O.; Goncalves, D.A.; Abrahamsohn, P.A.; Kettelhut, I.C.; Moriscot, A.S.; Aoki, M.S.; Miyabara, E.H. Leucine supplementation accelerates connective tissue repair of injured tibialis anterior muscle. Nutrients 2014, 6, 3981–4001. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, S.O.; Wang, W.; Ahtikoski, A.M.; Kjaer, M.; Han, X.Y.; Komulainen, J.; Kovanen, V.; Takala, T.E. Acute exercise induced changes in rat skeletal muscle mRNAs and proteins regulating type IV collagen content. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1292–R1300. [Google Scholar] [PubMed]
- Schwane, J.A.; Armstrong, R.B. Effect of training on skeletal muscle injury from downhill running in rats. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Myllyla, R.; Salminen, A.; Peltonen, L.; Takala, T.E.; Vihko, V. Collagen metabolism of mouse skeletal muscle during the repair of exercise injuries. Pflügers Archiv 1986, 407, 64–70. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Intracellular Proline Enrichment (%MPE) | |
|---|---|
| Sed (30) | 40.5 ± 0.2 |
| 1h-Con (6) | 42.2 ± 0.2 |
| 1h-AminoL40 (6) | 42.2 ± 0.3 |
| 4h (6) | 40.9 ± 0.6 |
| Day1-Con (15) | 40.9 ± 0.4 |
| Day1-AminoL40 (7) | 39.5 ± 1.0 |
| Day2 (9) | 39.4 ± 0.9 |
| Day4 (7) | 40.6 ± 0.2 |
| Day7 (8) | 39.5 ± 1.0 |
| Amino Acid | Sed (14) | Immediately after | 1 Day after | ||
|---|---|---|---|---|---|
| Con (6) | LEAA (6) | Con (7) | LEAA (8) | ||
| His | 46.6 ± 2.3 c, d | 57.3 ± 2.5 b, c | 71.0 ± 3.3 a | 39.2 ± 2.1 d | 58.2 ± 3.2 b |
| Ile | 71.5 ± 2.1 b | 84.8 ± 2.6 b | 368.3 ± 25.5 a | 62.1 ± 2.8 b | 313.5 ± 22.4 a |
| Leu | 115.2 ± 4.2 b | 140.5 ± 4.3 b | 1448.7 ± 101.3 a | 99.9 ± 5.2 b | 1304.8 ± 89.2 a |
| Lys | 574.1 ± 23.7 c | 630.9 ± 31.2 c | 1849.6 ± 91.3 a | 529.5 ± 33.2 c | 1599.4 ± 47.8 b |
| Met | 58.9 ± 1.7 c | 53.1 ± 1.9 c | 83.8 ± 4.8 b | 50.3 ± 2.2 c | 99.5 ± 4.7 a |
| Phe | 59.9 ± 2.1 b | 65.3 ± 2.4 b | 118.4 ± 7.3 a | 53.7 ± 1.7 b | 101.4 ± 5.8 a |
| Thr | 232.5 ± 10.4 b | 208.2 ± 6.4 b | 633.3 ± 70.8 a | 223.0 ± 11.6 b | 746.7 ± 46.4 a |
| Trp | 98.6 ± 4.5 b, c | 116.2 ± 13.0 a, b | 145.9 ± 10.0 a | 82.6 ± 6.6 c | 111.3 ± 4.4 b, c |
| Val | 164.0 ± 6.4 c | 195.8 ± 5.4 c | 1033.7 ± 50.5 a | 145.4 ± 6.2 c | 874.8 ± 44.9 b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, H.; Suzuki, H.; Inoue, Y.; Suzuki, K.; Kobayashi, H. Leucine-Enriched Essential Amino Acids Augment Mixed Protein Synthesis, But Not Collagen Protein Synthesis, in Rat Skeletal Muscle after Downhill Running. Nutrients 2016, 8, 399. https://doi.org/10.3390/nu8070399
Kato H, Suzuki H, Inoue Y, Suzuki K, Kobayashi H. Leucine-Enriched Essential Amino Acids Augment Mixed Protein Synthesis, But Not Collagen Protein Synthesis, in Rat Skeletal Muscle after Downhill Running. Nutrients. 2016; 8(7):399. https://doi.org/10.3390/nu8070399
Chicago/Turabian StyleKato, Hiroyuki, Hiromi Suzuki, Yoshiko Inoue, Katsuya Suzuki, and Hisamine Kobayashi. 2016. "Leucine-Enriched Essential Amino Acids Augment Mixed Protein Synthesis, But Not Collagen Protein Synthesis, in Rat Skeletal Muscle after Downhill Running" Nutrients 8, no. 7: 399. https://doi.org/10.3390/nu8070399