Mucosal MicroRNAs Expression Profiles before and after Exclusive Enteral Nutrition Therapy in Adult Patients with Crohn’s Disease


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Treatment
2.3. Intestinal Tissues Collection
2.4. RNA Extraction
2.5. miRNA Microarray
2.6. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
2.7. Data Analysis
3. Results
3.1. Characteristics of Participants
3.2. Alteration of MiRNAs Expression Profiles in Terminal Ileal Mucosa of CD after EEN Treatment (EEN-M)
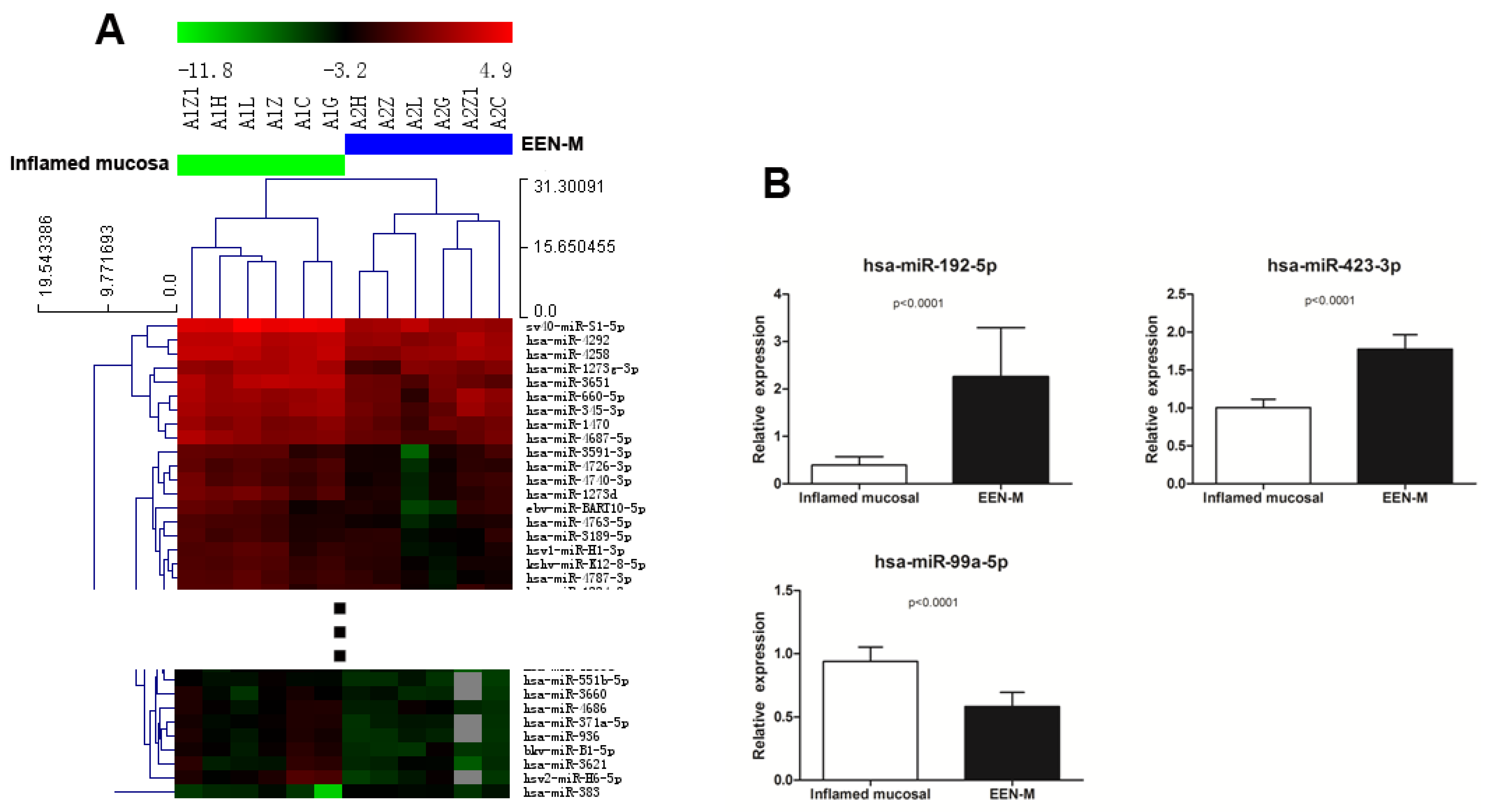
3.2.1. Comparison of miRNAs Expression Profiles in EEN-M and Inflamed Mucosa of Active CD
3.2.2. Comparison of miRNAs Expression Profiles in EEN-M and Non-inflamed Mucosa of Active CD
3.2.3. Comparison of miRNAs Expression Profiles in EEN-M and Healthy Controls
3.2.4. Several miRNAs Expression in All Groups
3.3. Correlation between miRNAs and CDAI and CRP before and after EEN
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H. The genetics and immunopathogenesis of inflammatory bowel disease. Nat. Rev. Immunol. 2008, 8, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Ventham, N.T.; Kennedy, N.A.; Nimmo, E.R.; Satsangi, J. Beyond gene discovery in inflammatory bowel disease: The emerging role of epigenetics. Gastroenterology 2013, 145, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- McKenna, L.B.; Schug, J.; Vourekas, A.; McKenna, J.B.; Bramswig, N.C.; Friedman, J.R.; Kaestner, K.H. MicroRNAs control intestinal epithelial differentiation, architecture, and barrier function. Gastroenterology 2010, 139, 1654–1664. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Guo, S.; Al-Sadi, R.; Ma, T.Y. MicroRNA regulation of intestinal epithelial tight junction permeability. Gastroenterology 2011, 141, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Fasseu, M.; Treton, X.; Guichard, C.; Pedruzzi, E.; Cazals-Hatem, D.; Richard, C.; Aparicio, T.; Daniel, F.; Soule, J.C.; Moreau, R.; et al. Identification of restricted subsets of mature microRNA abnormally expressed in inactive colonic mucosa of patients with inflammatory bowel disease. PLoS ONE 2010, 5, e13160. [Google Scholar] [CrossRef] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology 2012, 142, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Voitk, A.J.; Echave, V.; Feller, J.H.; Brown, R.A.; Gurd, F.N. Experience with elemental diet in the treatment of inflammatory bowel disease. Is this primary therapy? Arch. Surg. 1973, 107, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, E.; Gaunt, W.W.; Cardigan, T.; Garrick, V.; McGrogan, P.; Russell, R.K. The use of exclusive enteral nutrition for induction of remission in children with Crohn’s disease demonstrates that disease phenotype does not influence clinical remission. Aliment Pharmacol. Ther. 2009, 30, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Nakahigashi, M.; Saniabadi, A.R.; Iwata, T.; Maruyama, Y.; Umegae, S.; Matsumoto, K. Impacts of long-term enteral nutrition on clinical and endoscopic disease activities and mucosal cytokines during remission in patients with Crohn’s disease: A prospective study. Inflamm. Bowel Dis. 2007, 13, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Hiwatashi, N. Enteral nutrition for Crohn’s disease in Japan. Dis. Colon. Rectum. 1997, 40, S48–S53. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Fukuda, Y.; Tanida, N.; Hosomi, M.; Shimoyama, T. Home elemental enteral hyperalimentation (HEEH) for the maintenance of remission in patients with Crohn’s disease. Gastroenterology 1993, 28, 379–384. [Google Scholar]
- Yamamoto, T.; Nakahigashi, M.; Umegae, S.; Kitagawa, T.; Matsumoto, K. Impact of elemental diet on mucosal inflammation in patients with active Crohn’s disease: Cytokine production and endoscopic and histological findings. Inflamm Bowel Dis. 2005, 11, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Buchman, A.L. DNA-driven nutritional therapy of inflammatory bowel disease. Nutrition 2009, 25, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, M.T. Understanding the nutrigenomic definitions and concepts at the food-genome junction. OMICS. 2008, 12, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Segura, L.; Perez-Andrade, M.; Miranda-Rios, J. The emerging role of MicroRNAs in the regulation of gene expression by nutrients. J. Nutr. Nutr. 2013, 6, 16–31. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wu, R.; Gong, J.; Zhu, W.; Li, Y.; Wang, Z.; Li, N.; Li, J. Altered microRNAs expression in inflamed and non-inflamed terminal ileal mucosa of adult patients with active Crohn’s disease. J. Gastroenterol Hepatol. 2015, 30, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wu, R.; Zhu, W.; Gong, J.; Zhang, W.; Li, Y.; Gu, L.; Li, N.; Li, J. Effect of exclusive enteral nutrition on health-related quality of life for adults with active Crohn’s disease. Nutr. Clin. Pract. 2013, 28, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.F.; Liston, A. MicroRNA in the immune system, microRNA as an immune system. Immunology 2009, 127, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Pekow, J.R.; Kwon, J.H. MicroRNAs in inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, S.; Dassopoulos, T.; Harris, M.L.; Bayless, T.M.; Meltzer, S.J.; Brant, S.R.; Kwon, J.H. Identification of microRNAs associated with ileal and colonic Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Matsui, T.; Yao, T.; Takagi, Y.; Hirai, F.; Aoyagi, K.; Okada, M. Short-term efficacy of enteral nutrition in the treatment of active Crohn’s disease: A randomized, controlled trial comparing nutrient formulas. JPEN J. Parenter. Enter. Nutr. 2002, 26, 98–103. [Google Scholar] [CrossRef]
- Gearry, R.B.; Day, A.S.; Wall, C.L. Use of exclusive enteral nutrition in adults with Crohn’s disease: A review. World J. Gastroenterol. 2013, 19, 7652–7660. [Google Scholar]
- Burgess, L. Exclusive enteral nutrition and induction of remission of active Crohn’s disease in children. Expert Rev. Clin. Immunol. 2013, 9, 375–384. [Google Scholar]
- Feng, Y.; Li, Y.; Mei, S.; Zhang, L.; Gong, J.; Gu, J.; Zhang, W.; Zhu, W.; Li, N.; Li, J. Exclusive enteral nutrition ameliorates mesenteric adipose tissue alterations in patients with active Crohn’s disease. Clin. Nutr. 2014, 33, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Shi, T.; Zhou, Q.; Shi, S.; Zhao, R.; Shi, H.; Dong, L.; Zhang, C.; Zeng, K.; Chen, J.; et al. miR-141 Regulates colonic leukocytic trafficking by targeting CXCL12beta during murine colitis and human Crohn’s disease. Gut 2014, 63, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, C.; Liu, Y.; Tang, L.; Zheng, M.; Xu, C.; Song, J.; Meng, X. miR-122 targets NOD2 to decrease intestinal epithelial cell injury in Crohn’s disease. Biochem. Biophys. Res. Commun. 2013, 438, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ge, W.; Xu, L.; Qu, C.; Zhu, M.; Zhang, W.; Xiao, Y. miR-200b is involved in intestinal fibrosis of Crohn’s disease. Int. J. Mol. Med. 2012, 29, 601–606. [Google Scholar] [PubMed]
- Ludwig, K.; Fassan, M.; Mescoli, C.; Pizzi, M.; Balistreri, M.; Albertoni, L.; Pucciarelli, S.; Scarpa, M.; Sturniolo, G.C.; Angriman, I.; et al. PDCD4/miR-21 dysregulation in inflammatory bowel disease-associated carcinogenesis. Virchows Arch. 2013, 462, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Olaru, A.V.; Selaru, F.M.; Mori, Y.; Vazquez, C.; David, S.; Paun, B.; Cheng, Y.; Jin, Z.; Yang, J.; Agarwal, R.; et al. Dynamic changes in the expression of MicroRNA-31 during inflammatory bowel disease-associated neoplastic transformation. Inflamm. Bowel Dis. 2011, 17, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Chuang, A.Y.; Chuang, J.C.; Zhai, Z.; Wu, F.; Kwon, J.H. NOD2 expression is regulated by microRNAs in colonic epithelial HCT116 cells. Inflamm. Bowel Dis. 2014, 20, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Koukos, G.; Polytarchou, C.; Kaplan, J.L.; Morley-Fletcher, A.; Gras-Miralles, B.; Kokkotou, E.; Baril-Dore, M.; Pothoulakis, C.; Winter, H.S.; Iliopoulos, D. MicroRNA-124 regulates STAT3 expression and is down-regulated in colon tissues of pediatric patients with ulcerative colitis. Gastroenterology 2013, 145, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Shay, N.F.; Banz, W.J. Regulation of gene transcription by botanicals: Novel regulatory mechanisms. Annu. Rev. Nutr. 2005, 25, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Bull, C.; Fenech, M. Genome-health nutrigenomics and nutrigenetics: Nutritional requirements or ‘nutriomes’ for chromosomal stability and telomere maintenance at the individual level. Proc. Nutr. Soc. 2008, 67, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, D.; Deval, C.; Gouranton, E.; Landrier, J.F.; Scalbert, A.; Morand, C.; Mazur, A. Modulation of miRNA expression by dietary polyphenols in apoE deficient mice: A new mechanism of the action of polyphenols. PLoS ONE 2012, 7, e29837. [Google Scholar] [CrossRef] [PubMed]
- Davidson, L.A.; Wang, N.; Shah, M.S.; Lupton, J.R.; Ivanov, I.; Chapkin, R.S. n-3 Polyunsaturated fatty acids modulate carcinogen-directed non-coding microRNA signatures in rat colon. Carcinogenesis 2009, 30, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Nakamichi, I.; Esaki, M.; Asano, K.; Matsumoto, T.; Kitazono, T. Serum microRNA levels in patients with Crohn’s disease during induction therapy by infliximab. J. Gastroenterol. Hepatol. 2014, 29, 1207–1214. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer |
|---|---|
| hsa-miR-21-5p | GSP:5′GGGGGGTAGCTTATCAGACTG3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-31-5p | GSP:5′ GGGAGGCAAGATGCTGGC3′ R:5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-192-5p | GSP:5′GGGGCTGACCTATGAATTG3′ R:5′CAGTGCGTGTCGTGGAGT3′ |
| hsa-miR-423-3p | GSP:5′TAAGCTCGGTCTGAGGC3′ R:5′CAGTGCGTGTCGTGGA3′ |
| hsa-let-7b-5p | GSP:5′GGGGTGAGGTAGTAGGTTG3′ R:5′CAGTGCGTGTCGTGGA3′ |
| hsa-let-7a-2-3p | GSP:5′GCCCCTGTACAGCCTCCTA3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-495-5p | GSP:5′GGGGAAGTTGCCCATGTTA3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-199b-5p | GSP:5′GGGACCCCAGTGTTTAGACTAT3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-124-3p | GSP:5′GGGTAAGGCACGCGGT3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-301a-5p | GSP:5′GGGGGCTCTGACTTTATTGC3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| hsa-miR-99a-5p | GSP:5′GCCAACCCGTAGATCCGAT3′ R: 5′GTGCGTGTCGTGGAGTCG3′ |
| U6 | F:5′GCTTCGGCAGCACATATACTAAAAT3′ R:5′CGCTTCACGAATTTGCGTGTCAT3′ |
| CD Patients N = 25 | Controls N = 10 | |
|---|---|---|
| Male/Female | 17/8 | 4/6 |
| Age (years) | 27 (4.4) | 42 (10.1) |
| Duration of disease (months) | 14.6 (7.7) | n.a |
| Location | ||
| ileum | 6 | n.a |
| ileocolon | 19 | n.a |
| Behavior (non-stricturing, non-penetrating) | 25 | n.a |
| CDAI before EEN | 269.4 (55.5) | n.a |
| CRP before EEN | 33.2 (18.5) | 1.6 (1.3) |
| CDAI after EEN | 77.7 (20.6) *** | n.a |
| CRP after EEN | 3.6 (2.4) *** | n.a |
| Medication (sulfasalazine or mesalazine) | 10 | 0 |
| Smoker | 0 | 0 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Gong, J.; Li, Y.; Gu, L.; Cao, L.; Wang, Z.; Zhu, W.; Li, J. Mucosal MicroRNAs Expression Profiles before and after Exclusive Enteral Nutrition Therapy in Adult Patients with Crohn’s Disease. Nutrients 2016, 8, 519. https://doi.org/10.3390/nu8080519
Guo Z, Gong J, Li Y, Gu L, Cao L, Wang Z, Zhu W, Li J. Mucosal MicroRNAs Expression Profiles before and after Exclusive Enteral Nutrition Therapy in Adult Patients with Crohn’s Disease. Nutrients. 2016; 8(8):519. https://doi.org/10.3390/nu8080519
Chicago/Turabian StyleGuo, Zhen, Jianfeng Gong, Yi Li, Lili Gu, Lei Cao, Zhiming Wang, Weiming Zhu, and Jieshou Li. 2016. "Mucosal MicroRNAs Expression Profiles before and after Exclusive Enteral Nutrition Therapy in Adult Patients with Crohn’s Disease" Nutrients 8, no. 8: 519. https://doi.org/10.3390/nu8080519
APA StyleGuo, Z., Gong, J., Li, Y., Gu, L., Cao, L., Wang, Z., Zhu, W., & Li, J. (2016). Mucosal MicroRNAs Expression Profiles before and after Exclusive Enteral Nutrition Therapy in Adult Patients with Crohn’s Disease. Nutrients, 8(8), 519. https://doi.org/10.3390/nu8080519