The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action

Abstract
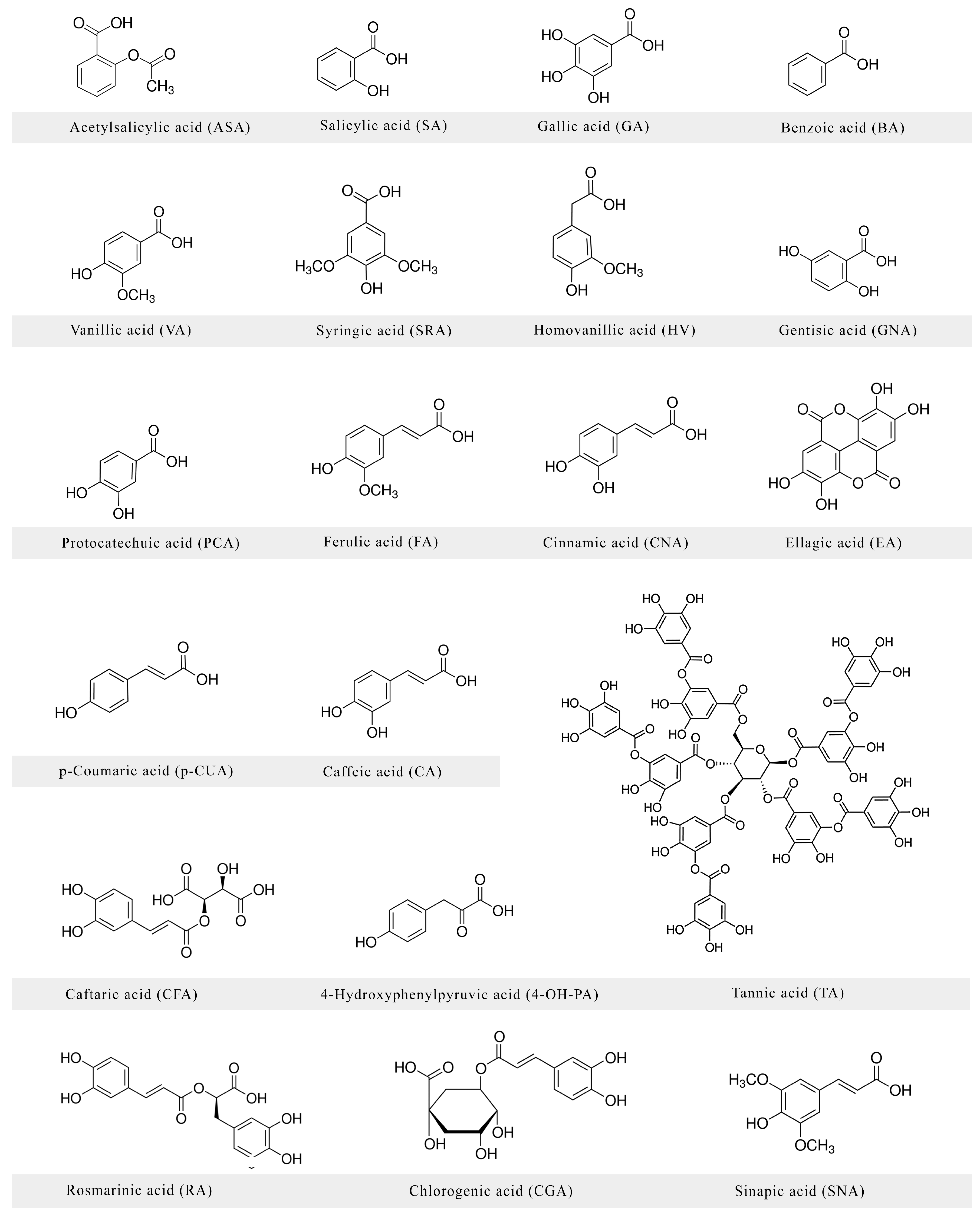
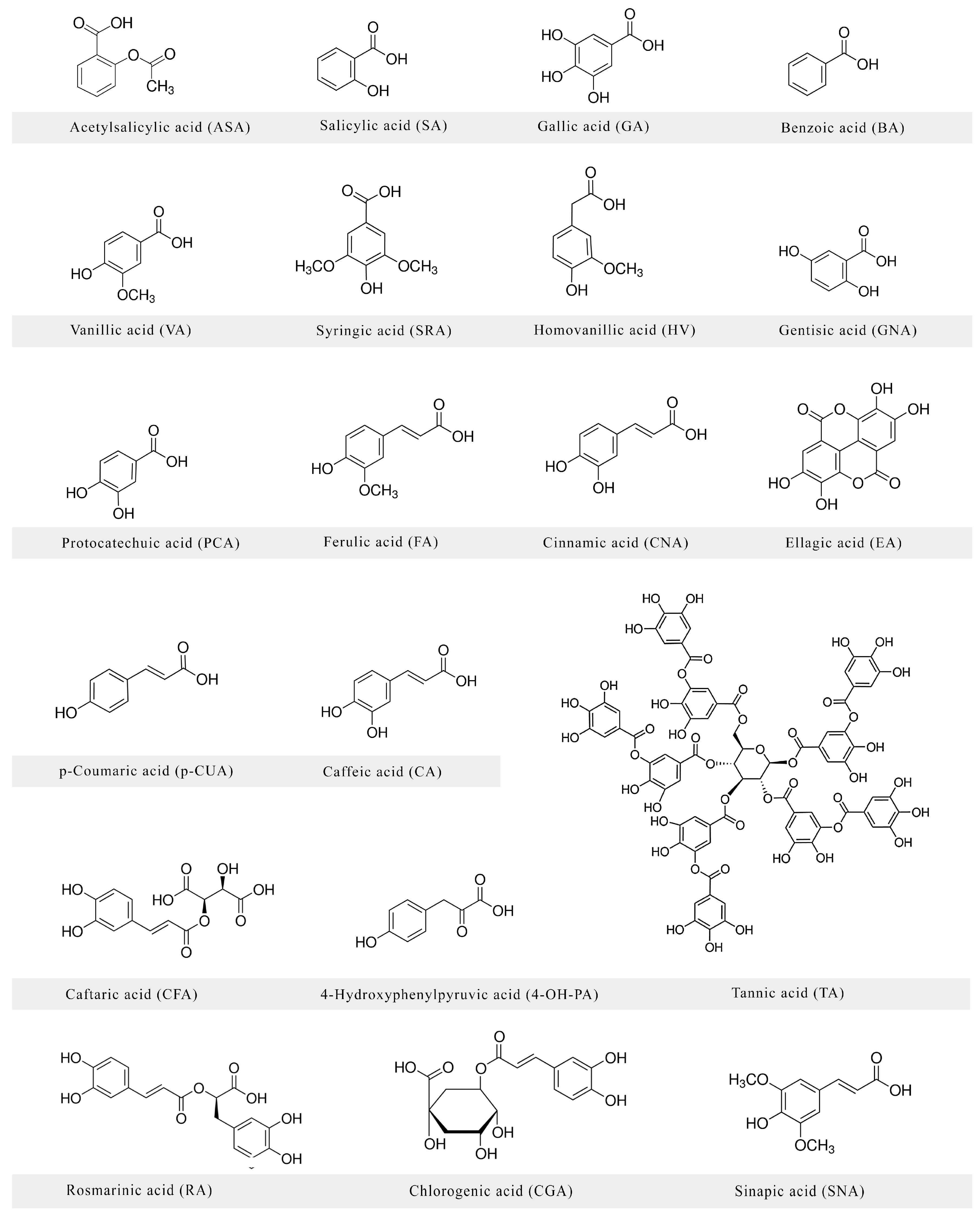
:1. Introduction
2. Neuroprotective Activities of Phenolic Acids
3. Penetration of Brain by Phenolic Acids
4. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 3NP | 3-Nitropropionic acid |
| 6-OHDA | 6-Hydroxydopamine |
| BAD | Bcl-2-associated death promoter |
| Bax | Bcl-2-like protein 4 |
| BCL | B-cell lymphoma |
| BCL-2 | B-cell lymphoma 2 |
| Bcr-Abl | fusion between break point cluster (Bcr) gene and the Abelson (Abl) tyrosine kinase gene |
| BDNF | Brain-derived neurotrophic factor |
| b.w. | Body weight |
| Casp-3 | Caspase-3 |
| Casp-8 | Caspase-8 |
| Casp-9 | Caspase-9 |
| CAT | Catalase |
| CCI | Chronic constriction injury |
| CD40 | Cluster of differentiation 40 |
| COX-2 | Cyclooxygenase-2 |
| CREB | cAMP response element-binding protein |
| CUMS | Chronic unexpected mild stress |
| EPO | Erythropoietin |
| ERK | Extracellular signal-regulated kinase, protein-serine/threonine kinase |
| ERK1 | Extracellular signal-regulated kinase 1, protein-serine/threonine kinase 1 |
| ERK2 | Extracellular signal-regulated kinase 2 protein-serine/threonine kinase 2 |
| FST | Forced swimming test |
| GFAP | Glial fibrillary acidic protein |
| GPx | Glutathione peroxidase |
| i.p. | Intraperitoneally |
| IL-1β | Interleukin 1β |
| IL-4 | Interleukin 4 |
| IL-5 | Interleukin 5 |
| IL-6 | Interleukin 6 |
| IL-13 | Interleukin 13 |
| iNOS | Inducible nitric oxide synthase |
| JNK | c-Jun N-terminal kinase |
| LDH | Lactate dehydrogenase |
| LPS | Lipopolysaccharide |
| MAO | Monoamine oxidase |
| MDA | Malondialdehyde |
| MEK | Mitogen-activated protein kinase |
| MHX | major histocompatibility complex II molecules |
| NF-κB | Nuclear factor-κB |
| NO | Nitric oxide |
| NRF1 | Nuclear respiratory factor 1 |
| Nrf2 | Nuclear factor (erythroid-derived 2)-like 2 |
| Nrf2/HO-1 | Nuclear factor (erythroid-derived 2)-like 2/heme oxygenase-1 |
| p38 | MAP Kinase (MAPK), CSBP Cytokinin-Specific Binding Protein or RK |
| p90RSK | MAPK-activated protein kinase-1 (MAPKAP-K1) |
| pCREB | phosphorylated cAMP response element-binding protein |
| PD | Parkinson disease |
| PGE2 | Prostaglandin E2 |
| p.o. | Orally |
| p-p38 | phosphorylated p38 |
| PTZ | Pentylenetetrazol |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| TBI | Traumatic brain injury |
| Th1 | T helper type 1 (Th1) cells |
| Th2 | T helper type 2 (Th2) cells |
| TNF-α | Tumor necrosis factor-α |
| TST | Tail suspension test |
References
- Mattila, P.; Hellström, J.; Törrönen, R. Phenolic acids in berries, fruits, and beverages. J. Agric. Food Chem. 2006, 54, 7193–7199. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Estruch, R. Nut consumption and age-related disease. Maturitas 2016, 84, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P. Phenolic compounds of cereals and their antioxidant capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yan, Y.; Li, F.; Zhang, D. Fruit and vegetable consumption and the risk of depression: A meta-analysis. Nutrition 2016, 32, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Micek, A.; Castellano, S.; Pajak, A.; Galvano, F. Coffee, tea, caffeine and risk of depression: A systematic review and close-response meta-analysis of observational studies. Mol. Nutr. Food Res. 2016, 60, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D.; Baranowska-Wójcik, E.; Borowiec, K. Phenolic acids exert anticholinesterase and cognition-improving effects. Curr. Alzheimer Res 2017, in press. [Google Scholar]
- Li, G.; Ruan, L.; Chen, R.; Wang, R.; Xie, X.; Zhang, M.; Chen, L.; Yan, Q.; Reed, M.; Chen, J.; et al. Synergistic antidepressant-like effect of ferulic acid in combination with piperine: Involvement of monoaminergic system. Metab. Brain Dis. 2015, 30, 1505–1514. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, D.; Zhang, C.; Li, G.; Zhang, N.; Ruan, L.; Yan, Q.; Li, J.; Yu, X.; Xie, X.; et al. Antidepressant-like effects of ferulic acid: Involvement of serotonergic and norepinergic systems. Metab. Brain Dis. 2015, 30, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, J.; Rodriguez, A.F.; Rós Ade, S.; de Castro, A.B.; de Lima, D.D.; Magro, D.D.; Zeni, A.L. Ferulic acid chronic treatment exerts antidepressant-like effect: Role of antioxidant defense system. Metab. Brain Dis. 2015, 30, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.M.; Hu, C.Y.; Shen, J.D.; Wu, S.H.; Li, Y.C.; Yi, L.T. Elevation of synaptic protein is associated with the antidepressant-like effects of ferulic acid in a chronic model of depression. Physiol. Behav. 2017, 169, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H.; Wang, T.; Jiang, N.; Yu, P.; Chong, Y.; Fu, F. Ferulic acid ameliorates nerve injury induced by cerebral ischemia in rats. Exp. Ther. Med. 2015, 9, 972–976. [Google Scholar] [CrossRef] [PubMed]
- Koh, P.O. Ferulic acid attenuates the down-regulation of MEK/ERK/p90RSK signaling pathway in focal cerebral ischemic injury. Neurosci. Lett. 2015, 588, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.J.; Fan, S.Y.; Yang, J.H.; Wei, Y.; Zhu, Z.H.; Lou, Y.J.; Guo, Y.; Wan, H.T.; Xie, Y.Q. Pharmacokinetic and pharmacodynamic analysis of ferulic acid-puerarin-astragaloside in combination with neuroprotective in cerebral ischemia/reperfusion injury in rats. Asian Pac. J. Trop. Med. 2015, 8, 299–304. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, D.; Yu, X.; Xie, X.; Wang, L.; Lian, L.; Fei, N.; Chen, J.; Zhu, N.; Wang, G.; et al. The antinociceptive effects of ferulic acid on neuropathic pain: Involvement of descending monoaminergic system and opioid receptors. Oncotarget 2016, 7, 20455–20468. [Google Scholar] [CrossRef] [PubMed]
- Aswar, M.; Patil, V. Ferulic acid ameliorates chronic constriction injury induced painful neuropathy in rats. Inflammopharmacology 2016, 24, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Chellappan, D.R.; Chinnaswamy, P.; Thulasingam, S. Ferulic acid pretreatment mitigates MPTP-induced motor impairment and histopathological alterations in C57BI/6 mice. Pharm. Biol. 2015, 53, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.; Javed, H.; Azimullah, S.; Khair, S.B.A.; Haque, M.E. Neuroprotective potential of ferulic acid in the rotenone model of Parkinson’s disease. Drug Des. Devel. Ther. 2015, 9, 5499–5510. [Google Scholar] [PubMed]
- Wu, J.L.; Shen, M.M.; Yang, S.X.; Wang, X.; Ma, Z.C. Inhibitory effect of ferulic acid on neuroinflammation in LPS-activated microglia. Chin. Pharm. Bull. 2015, 31, 97–102. [Google Scholar]
- Huang, H.; Hong, Q.; Tan, H.L.; Xiao, C.R.; Gao, Y. Ferulic acid prevents LPS-induced up-regulation of PDE4B and stimulates the cAMP/CREB signaling pathway in PC12 cells. Acta Pharm. Sin. 2016, 37, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.C.; Kuan, C.Y.; Subramaniam, S.; Zhao, J.Y.; Sivasubramaniam, S.; Chang, H.Y.; Lin, F.H. A potent inhibition of oxidative stress induced gene expression in neural cells by sustained ferulic acid release from chitosan based hydrogel. Mat. Sci. Eng. C 2015, 49, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Wang, C.C.; Huang, H.M.; Lin, C.L.; Leu, S.J.; Lee, Y.L. Ferulic acid induces Th1 responses by modulating the function of dendritic cells and ameliorates Th2-mediated allergic airway inflammation in mice. Evid-Based Compl. Alt. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Peng, Y.F.; Hou, C.W. Ferulic acid protects PC12 neurons against hypoxia by inhibiting the p-MAPKs and COX-2 pathways. Iran. J. Basic Med. Sci. 2015, 18, 478–484. [Google Scholar] [PubMed]
- Shen, Y.; Zhang, H.; Wang, L.; Qian, H.; Qi, H.; Miao, X.; Cheng, L.; Qi, X. Protective effect of ferulic acid against 2,2′-azobis(2-amidinopropane) dihydrochloride-induced oxidative stress in PC12 cells. Cell. Mol. Biol. 2016, 62, 109–116. [Google Scholar] [PubMed]
- Basu Mallik, S.; Mudgal, J.; Nampoothiri, M.; Hall, S.; Dukie, S.A.; Grant, G.; Rao, C.M.; Arora, D. Caffeic acid attenuates lipopolysaccharide-induced sickness behavior and neuroinflammation in mice. Neurosci. Lett. 2016, 632, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Fazili, N.A.; Naeem, A. Anti-fibrillation potency of caffeic acid against an antidepressant induced fibrillogenesis of human α-synuclein: Implications for Parkinson’s disease. Biochimie 2015, 108, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Baj, J.P.; Hu, X.L.; Jiang, X.W.; Tian, X.; Zhao, Q.C. Caffeic acids from roots of Arctium lappa and their neuroprotective activity. Chin. Trad. Herb. Drugs 2015, 46, 163–168. [Google Scholar]
- Coelho, V.R.; Vieira, C.G.; de Souza, L.P.; Moysés, F.; Basso, C.; Picada, J.N.; Pereira, P. Antiepileptogenic, antioxidant and genotoxic evaluation of rosmarinic acid and its metabolite caffeic acid in mice. Life Sci. 2015, 122, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Coelho, V.R.; Vieira, C.G.; de Souza, L.P.; da Silva, L.L.; Pflüger, P.; Regner, G.G.; Papke, D.K.; Picada, J.N.; Pereira, P. Behavioral and genotoxic evaluation of rosmarinic and caffeic acid in acute seizure models induced by pentylenetetrazole and pilocarpine in mice. Naunyn-Schmiedeberg's Arch. Pharmacol. 2016, 389, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Kuo, P.L.; Chen, C.W.; Wu, J.S.B.; Shen, S.C. Caffeic acid improves memory impairment and brain glucose metabolism via ameliorating cerebral insulin and leptin signaling pathways in high-fat diet–induced hyperinsulinemic rats. Food Res. Int. 2015, 77, 24–33. [Google Scholar] [CrossRef]
- Alkis, H.E.; Kuzhan, A.; Dirier, A.; Tarakcioglu, M.; Demir, E.; Saricicek, E.; Demir, T.; Ahlatci, A.; Demirci, A.; Cinar, K.; Taysi, S. Neuroprotective effects of propolis and caffeic acid phenethyl ester (CAPE) on the radiation-injured brain tissue (Neuroprotective effects of propolis and CAPE). Int. J. Radiat. Res. 2015, 13, 297–303. [Google Scholar]
- Khan, M.; Baarine, M.; Singh, I. Therapeutic potential of caffeic acid phenethyl ester in neurodegenerative diseases. In Caffeic Acid: Biological Properties, Structure and Health Effects; Nova Science Publishers: New York, NY, USA, 2015. [Google Scholar]
- Ginis, Z.; Ozturk, G.; Albayrak, A.; Kurt, S.N.; Albayrak, M.; Fadillioglu, E. Protective effects of caffeic acid phenethyl ester on ifosfamide-induced central neurotoxicity in rats. Toxicol. Ind. Health. 2016, 32, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Akyol, S.; Erdemli, H.K.; Amautou, F.; Akyol, O. In vitro and in vivo neuroprotective effect of caffeic acid phenethyl ester. J. Intercult. Ethnopharmacol. 2015, 4, 192–193. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.A.G.D.; Martins, N.M.; Silva, R.D.B.; Ferreira, R.S.; Sisti, F.M.; dos Santos, A.C. Caffeic acid phenethyl ester (CAPE) protects PC12 cells from MPP+ toxicity by inducing the expression of neuron-typical proteins. Neurotoxicology 2014, 45, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Tang, Z.J.; Han, Y.Z. Neuroprotective effects of caffeic acid phenethyl ester against sevoflurane-induced neuronal degeneration in the hippocampus of neonatal rats involve MAPK and PI3K/Akt signaling pathways. Mol. Med. Rep. 2016, 14, 3403–3412. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.F.; Kuo, Y.H.; Yeh, W.L.; Wu, C.Y.; Lin, H.Y.; Lai, S.W.; Liu, Y.S.; Wu, L.H.; Lu, J.K.; Lu, D.Y. Regulatory effects of caffeiccidphenethyl ester on neuroinflammation in microglial cells. Int. J. Mol. Sci. 2015, 16, 5572–5589. [Google Scholar] [CrossRef] [PubMed]
- Balkhi, H.M.; Gul, T.; Haq, E. Anti-neoplastic and calcium modulatory action of caffeic acid phenethyl ester and desatinib in C6 glial cells: A therapeutic perspective. CNS Neurol. Disord. Drug Targets 2016, 15, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Bak, J.; Kim, H.J.; Kim, S.Y.; Choi, Y.S. Neuroprotective effect of caffeic acid phenethyl ester in 3-nitropropionic acid-induced striatal neurotoxicity. Korean J. Physiol. Pharmacol. 2016, 20, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Yamazawa, T. Chlorogenic, a polyphenol in coffee, protects neurons against glutamate neurotoxicity. Life Sci. 2015, 139, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.J.; Luo, T.; Wu, F.; Mei, Y.W.; Peng, J.; Liu, H.; Li, H.R.; Zhang, S.L.; Dong, J.H.; Fang, Y.; et al. Involvement of TLR2 and TLR9 in the anti-inflammatory effects of chlorogenic acid in HSV-1-infected microglia. Life Sci. 2015, 127, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.M.; Chen, H.X.; Li, H.; Tang, Y.; Yang, L.; Cao, S.S.; Qin, D.L. Antidepressant potential of chlorogenic acid-enriched extract from Eucommia ulmoides Oliver bark with neuron protection and promotion of serotonin release through enhancing synapsin I expression. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Gul, Z.; Demircan, C.; Bagdas, D.; Buyukuysal, R.L. Protective effects of chlorogenic acid and its metabolites on hydrogen peroxide-induced alterations in rat brain slices: a comparative study with resveratrol. Neurochem. Res. 2016, 41, 2075–2085. [Google Scholar] [CrossRef] [PubMed]
- Aseervatham, G.S.B.; Suryakala, U.; Doulethunisha; Sundaram, S.; Bose, P.C.; Sivasudha, T. Expression pattern of NMDA receptors reveals antiepileptic potential of apigenin 8-Cglucoside and chlorogenic acid in pilocarpine induced epileptic mice. Biomed. Pharmacother. 2016, 82, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.Q.; Wang, J.X.; Wei, J.X.; Shu, Y.H.; Xiao, L.; Lu, X.M. Beneficial effects of chlorogenic acid on alcohol-induced damage in PC12 cells. Biomed. Pharmacother. 2016, 79, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Taram, F.; Winter, A.N.; Linseman, D.A. Neuroprotection comparison of chlorogenic acid and its metabolites against mechanistically distinct cell death-inducing agents in cultured cerebellar granule neurons. Brain Res. 2016, 1648, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Fetoni, A.R.; Paciello, F.; Rolesi, R.; Eramo, S.L.; Mancuso, C.; Troiani, D.; Paludetti, G. Rosmarinic acid up-regulates the noise-activated Nrf2/HO-1 pathway and protects against noise-induced injury in rat cochlea. Free Radic. Biol. Med. 2015, 85, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Xu, H.M.; Jiang, H.; Hie, J.X. Protective effects of rosmarinic acid against iron-induced neurotoxicity in SK-N-SH cells. Am. J. Hematol. 2015, 91, E95. [Google Scholar]
- Lee, A.Y.; Wu, T.T.; Hwang, B.R.; Lee, J.; Lee, M.H.; Lee, S.; Cho, E.J. The neuro-protective effect of the methanolic extract of Perillafrutescens var. japonica and rosmarinic acid against H2O2-induced oxidative stress in C6 glial cells. Biomol. Ther. 2016, 24, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.M.D.; Passos, C.D.; Kieling Rubio, M.A.; Mendonça, J.N.; Lopes, N.P.; Henriques, A.T. Combining in vitro and in silico approaches to evaluate the multifunctional profile of rosmarinic acid from Blechnum brasiliense on targets related to neurodegeneration. Chem. Biol. Interact. 2016, 254, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.J.; Yang, Y.; Wang, H.Y.; Tao, B.Z.; Wang, J.; Wang, Z.F.; Zhou, D.B. Spinal cord injury effectively ameliorated by neuroprotective effects of rosmarinic acid. Nutr. Neurosci. 2017, 20, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Khamse, S.; Sadr, S.S.; Roghani, M.; Hasanzadeh, G.; Mohammadian, M. Rosmarinic acid exerts a neuroprotective effect in the kainite rat model of temporat lobe epilepsy: Underlying mechanisms. Pharm. Biol. 2015, 53, 1818–1825. [Google Scholar] [CrossRef] [PubMed]
- Grigoletto, J.; de Oliveira, C.V.; Grauncke, A.C.; de Souza, T.L.; Souto, N.S.; de Freitas, M.L.; Furian, A.F.; Santos, A.R.S.; Oliveira, M.S. Rosmarinic acid is anticonvulsant against seizures induced by pentylenetetrazol and pilocarpine in mice. Epilepsy Behav. 2016, 62, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, R.; Singh, D.; Prakash, A.; Mishra, N. Development, characterization and nasal delivery of rosmarinic acid-loaded solid lipid nanoparticles for the effective management of Huntingtons disease. Drug Deliv. 2015, 22, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; El Omri, A.; Han, J.; Isoda, H. Antidepressant-like effects of rosmarinic acid through mitogen-activated protein kinase phosphatase-1 and brain-derived neurotrophic factor modulation. J. Funct. Food. 2015, 14, 758–766. [Google Scholar] [CrossRef]
- Shan, Y.; Wang, D.D.; Xu, Y.X.; Wang, C.; Cao, L.; Liu, Y.S.; Zhu, C.Q. Aging as a precipitating factor in chronic restraint stress-induced tau aggregation pathology, and the protective effects of rosmarinic acid. J. Alzheimers Dis. 2016, 49, 829–844. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Kim, H.B.; Choi, G.Y.; Lee, S.; Lee, S.O.; Kim, S.; Park, J.H. Acute rosmarinic acid treatment enhances long-term potentiation, BDNF and GluR-2 protein expression, and cell survival rate against scopolamine challenge in rat organotypic hippocampal slice cultures. Biochem. Biophs. Res. Commun. 2016, 475, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Ramanauskiene, K.; Raudonis, R.; Majiene, D. Rosmarinic acid and Melissa officinalis extracts differently affect glioblastoma cells. Oxid. Med. Cell. Longev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh Rahbardar, M.; Amin, B.; Mehri, S.; Mirnajafi-Zadeh, S.J.; Hosseinzadeh, H. Anti-inflammatory effects of ethanolic extract of Rosmarinus officinalis L. and rosmarinic acid in a rat model of neuropathic pain. Biomed. Pharmacother. 2016, 86, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Güven, M.; Yuksel, Y.; Sehitoglu, M.H.; Tokmak, M.; Aras, A.B.; Akman, T.; Golge, U.H.; Goksel, F.; Karavelioglu, E.; Cosar, M. The effect of coumaric acid on ischemia-reperfusion injury of sciatic nerve in rats. Inflammation 2015, 38, 2124–2132. [Google Scholar] [CrossRef] [PubMed]
- Güven, M.; Aras, A.B.; Akman, T.; Sen, H.M.; Ozkan, A.; Salis, O.; Sehitoglu, I.; Kalkan, Y.; Silan, C.; Deniz, M.; et al. Neuroprotective effect of p-coumaric acid in rat model of embolic cerebral ischemia. Iran. J. Basic Med. Sci. 2015, 18, 356–363. [Google Scholar] [PubMed]
- Güven, M.; Sehitoglu, M.H.; Yuksel, Y.; Tomkak, M.; Aras, A.B.; Akman, T.; Golge, U.H.; Karavelioglu, E.; Bal, E.; Cosar, M. The neuroprotective effect of p-coumaric acid on spinal cord ischemia/reperfusion injury in rats. Inflammation 2015, 38, 1986–1995. [Google Scholar] [CrossRef] [PubMed]
- Shailasree, S.; Venkataramana, M.; Niranjana, S.R. Cytotoxic effect of p-coumaric acid on neuroblastoma, N2a cell via generation of reactive oxygen species leading to dysfunction of mitochondria inducing apoptosis and autophagy. Mol. Neurobiol. 2015, 51, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Zare, K.; Eidi, A.; Roghani, M.; Haeri-Rohani, A. The neuroprotective potential of sinapic acid in the 6-hydroxydopamine-induced hem-parkinsonian rat. Metab. Brain Dis. 2015, 30, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Huang, H.Y.; Yang, Y.X.; Guo, J.Y. Cinnamic aldehyde treatment alleviates chronic unexpected stress-induced depressive-like behaviors via targeting cyclooxygenase-2 in mid-aged rats. J. Ethnopharmacol. 2015, 162, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Thrash-Williams, B.; Karuppagounder, S.S.; Bhattacharya, D.; Ahuja, M.; Suppiramaniam, V.; Dhanasekaran, M. Methamphetamine-induced dopaminergic toxicity prevented owing to the neuroprotective effects of salicylic acid. Life Sci. 2016, 1, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Cetin, D.; Hacimuftuoglu, A.; Tatar, A.; Turkez, H.; Togar, B. The in vitro protective effect of salicylic acid against paclitaxel and cisplatin-induced neurotoxicity. Cytotechnology 2016, 68, 1361–1367. [Google Scholar] [CrossRef] [PubMed]
- Rosińczuk, J.; Dymarek, R.; Całkosiński, I. Histopatological, ultrastructural, and immunohistochemical assessment of hippocampus structures of rats exposed to TCDD and high doses of tocopherol and acetylsalicylic acid. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, H.C.; Taha, A.Y.; Rapoport, S.I.; Yuan, Z.X. Low-dose aspirin (acetylsalicylate) prevents increases in brain PGE2, 15-epi-lipoxin A4 and 8-isoprostane in 9 month-old HIV-1 transgenic rats, a model for HIV-1 associated neurocognitive disorders. Prostag. Leukot. Ess. 2015, 96, 25–30. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Xia, C.; Zhang, J.H.; Li, X.Q.; Zhou, Z.H.; Li, F.P.; Li, W.; Lv, Y.; Chen, H.S. Clopidogrel plus aspirin versus aspirin alone for preventing early neurological deterioration in patients with acute ischemic stroke. J. Clin. Neurosci. 2015, 22, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Han, Z.; Wang, C.; Zhou, Q.; Lin, J. Statin and aspirin pretreatment are associated with lower neurological deterioration and platelet activity in patients with acute ischemic stroke. J. Stroke Cerebrovasc. Dis. 2017, 26, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.P.; Lin, F.C.; Lee, J.K.W.; Lee, C.T.C. Aspirin use associated with amyotrophic lateral sclerosis: A total population-based case control study. J. Epidemiol. 2015, 25, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Roehrich, M.E.; Wyss, J.C.; Kumar, R.; Pascual, M.; Golshayan, D.; Vssalli, G. Additive effects of rapamycin and aspirin on dendritic cell allostimulatory capacity. Immunopharm. Immunot. 2015, 37, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.X.; Du, F.; Jiang, L.R.; Gong, J.; Zhou, Y.F.; Luo, Q.Q.; Qian, Z.M.; Ke, Y. Effects of aspirin on expression of iron transport and storage proteins in BV-2 microglial cells. Neurochem. Int. 2015, 91, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Massocatto, C.L.; Moreira, N.M.; Muniz, E.; Pinge-Filho, P.; Rossi, R.M.; Araújo, E.J.; Sant’Ana, D.M. Aspirin prevents atrophy of esophageal nitrergic myenteric neurons in a mouse model of chronic Chagas disease. Dis. Esophagus 2016. [Google Scholar] [CrossRef] [PubMed]
- Abou-Abbass, H.; Bahmad, H.; Abou-El-Hassan, H.; Zhu, R.; Zhou, S.; Dong, X.; Hamade, E.; Mallah, K.; Zebian, A.; Ramadan, N.; et al. Deciphering glycomics and neuroproteomic alterations in experimental traumatic brain injury: Comparative analysis of aspirin and clopidogrel treatment. Electrophoresis 2016, 37, 1562–1576. [Google Scholar] [CrossRef]
- Lowthian, J.A.; Britt, C.J.; Rance, G.; Lin, F.R.; Woods, R.L.; Wolfe, R.; Nelson, M.R.; Dillon, H.A.; Ward, S.; Reid, C.M.; et al. Slowing the progression of age-related hearing loss: Rationale and study design of the ASPIRIN in HEARING, retinal vessels imaging and neurocognition in older generations (ASPREE-HEARING) trial. Contemp. Clin. Trials 2016, 46, 60–66. [Google Scholar] [PubMed]
- Zhang, Z.; Li, G.; Szeto, S.S.; Chong, C.M.; Quan, Q.; Huang, C.; Cui, W.; Guo, B.; Wang, Y.; Han, Y.; et al. Examining the neuroprotective effects of protocatechuic acid and chrysin on in vitro and in vivo models of Parkinson disease. Free Radic. Biol. Med. 2015, 84, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Semaming, Y.; Sripetchwandee, J.; Sa-nguanmoo, P.; Pintana, H.; Pannangpetch, P.; Chattipakorn, N.; Chattipakorn, S.C. Protocatechuic acid protects brain mitochondrial function in streptozotocin-induced diabetic rats. Appl. Physiol. Nutr. Metab. 2015, 40, 1078–1081. [Google Scholar] [CrossRef]
- Ju, D.T.; Liao, H.E.; Shibu, M.A.; Ho, T.J.; Padma, V.V.; Tsai, F.J.; Chung, L.C.; Day, C.H.; Lin, C.C.; Huang, C.Y. Nerve regeneration potential of protocatechuic acid in RSC96 Schwann cells by induction of cellular proliferation and migration through IGF-IR-PI3K-Akt signaling. Chin. J. Physiol. 2015, 58, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Ning, Q.-Q.; Liu, S.; Li, Y.-C.; Li, L.; Yu, Y.; Zhao, W.-X.; Zhang, X.-L. Protective effect of protocatechuic acid on midbrain dopaminergic neurons injured by 1-methyl-4-phenylpyridinium. Chin. Trad. Herb. Drugs 2016, 47, 2497–2501. [Google Scholar]
- Wang, H.Y.; Wang, H.; Wang, J.H.; Wang, Q.; Ma, Q.F.; Chen, Y.Y. Protocatechuic acid inhibits inflammatory responses in LPS-stimulated BV2 microglia via NF-κB and MAPKs signaling pathways. Neurochem. Res. 2015, 40, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Su, X.Y.; Wang, X.H.; Su, J.L.; Lian, Y.; Zhang, X.L. Effects of protocatechuic acid on expression of D2DR, iNOS, and TH in striatum and midbrain of Parkinson’s disease model mice. Chin. Trad. Herb. Drugs 2015, 46, 866–870. [Google Scholar]
- Ju, D.T.; Kuo, W.W.; Ho, T.J. Protocatechuic acid from Alpinia oxyphylla induces Schwann cell migration via ERK1/2, JNK and p38 activation. Am. J. Chin. Med. 2015, 43, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Naghizadeh, B.; Mansouri, M.T. Protective effects of gallic acid against streptozotocin-induced oxidative damage in rat striatum. Drug Res. 2015, 65, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Pemminati, S. Effect of gallic acid on antioxidative enzymes activities in depression. Indian J. Psychiat. 2015, 57, S125. [Google Scholar]
- Hajimoradi, M.; Fazilati, M.; Gharib-Naseri, M.K.; Sarkaki, A. Gallic acid and exercise training improve motor function, nerve conduction velocity but not pain sense reflex after experimental sciatic nerve crush in male rats. Avicenna J. Phytomed. 2015, 5, 288–297. [Google Scholar] [PubMed]
- Moghadas, M.; Edalatmanesh, M.A.; Robati, R. Histopathological analysis from gallic acid administration on hippocampal cell density, depression, and anxiety related behaviors in a trimethyltin intoxication model. Cell. J. 2016, 17, 659–667. [Google Scholar] [PubMed]
- Oyagbemi, A.A.; Omobowale, T.O.; Saba, A.B.; Olowu, E.R.; Dada, R.O.; Akinrinde, A.S. Gallic acid ameliorates cyclophosphamide-induced neurotoxicity in Wistar rats through free radical scavenging activity and improvement in antioxidant defense system. J. Diet. Suppl. 2016, 13, 402–419. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.S.; Chou, C.T.; Liao, W.C.; Shieh, P.; Kuo, D.H.; Kuo, C.C.; Jan, C.R.; Liang, W.Z. The effect of gallic acid on cytotoxicity, Ca2+ homeostasis and ROS production in DBTRG-05MG human glioblastoma cells and CTX TNA2 rat astrocytes. Chem. Biol. Interact. 2016, 252, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Tüzmen, M.N.; Yücel, N.C.; Kalburcu, T.; Demiryas, N. Effects of curcumin and tannic acid on the aluminum-and lead-induced oxidative neurotoxicity and alterations in NMDA receptors. Toxicol. Mech. Method. 2015, 25, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Ashafaq, M.; Tabassum, H.; Vishnoi, S.; Salman, M.; Raisuddin, S.; Parvez, S. Tannic acid alleviates lead acetate-induced neurochemical perturbations in rat brain. Neurosci. Lett. 2016, 617, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Sen, H.M.; Ozkan, A.; Güven, M.; Akman, T.; Aras, A.B.; Sehitoglu, I.; Alacam, H.; Silan, C.; Cosar, M.; Ozisik Karaman, H.I. Effects of tannic acid on the ischemic brain tissue of rats. Inflammation 2015, 38, 1624–1630. [Google Scholar] [CrossRef] [PubMed]
- Ashafaq, M.; Tabassum, H.; Parvez, S. Modulation of behavioral deficits and neurodegeneration by tannic acid in experimental stroke challenged Wistar rats. Mol. Neurobiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ibero-Baraibar, I.; Perez-Cornago, A.; Ramirez, M.J.; Martinez, J.A.; Zulet, M.A. An increase in plasma homovanillic acid with cocoa extract consumption is associated with the alleviation of depressive symptoms in overweight or obese adults on an energy restricted diet in a randomized controlled trial. J. Nutr. 2016, 146, 897S–904S. [Google Scholar] [CrossRef] [PubMed]
- Neider, D.; Lindstrom, L.H.; Boden, R. Risk factors for suicide among patients with schizophrenia: A cohort study focused on cerebrospinal fluid levels of homovanillic acid and 5-hydroxyindoleacetic acid. Neuropsych. Dis. Treat. 2016, 12, 1711–1714. [Google Scholar] [CrossRef] [PubMed]
- Van de Kerkhof, N.W.A.; Fekkes, D.; van der Heijden, F.M.M.A.; Egger, J.I.M.; Verhoeven, W.M.A. Relationship between plasma homovanillic acid and outcome in patients with psychosis spectrum disorders. Neuropsychobiology 2015, 71, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Güven, M.; Aras, A.B.; Topaloğlu, N.; Özkan, A.; Şen, H.M.; Kalkan, Y.; Okuyucu, A.; Akbal, A.; Gökmen, F.; Coşar, M. The protective effect of syringic acid on ischemia injury in rat brain. Turk. J. Med. Sci. 2015, 45, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Tokmak, M.; Yuksel, Y.; Sehitoglu, M.H.; Güven, M.; Akman, T.; Aras, A.B.; Cosar, M.; Abbed, K.M. The neuroprotective effect of syringic acid on spinal cord ischemia/reperfusion injury in rats. Inflammation 2015, 38, 1969–1978. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Du, Z.; Lu, G.; Li, P.; Wang, L. Syringic acid protects retinal ganglion cells against H2O2-induced apoptosis through the activation of Pl3K/Akt signaling pathway. Cell. Mol. Biol. 2016, 62, 50–54. [Google Scholar] [PubMed]
- Cao, Y.; Zhang, L.; Sun, S.; Yi, Z.; Jiang, X.; Jia, D. Neuroprotective effects of syringic acid against OGD/R-induced injury in cultured hippocampal neuronal cells. Int. J. Mol. Med. 2016, 38, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Alfredsson, C.F.; Ding, M.; Liang, Q.-L.; Sundstrӧm, B.E.; Nånberg, E. Ellagic acid induces a dose-and time-dependent depolarization of mitochondria and activation of caspase-9 and -3 in human neuroblastoma cells. Biomed. Pharmacother. 2014, 68, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Alfredsson, C.F.; Rendel, F.; Liang, Q.L.; Sundstrӧm, B.E.; Nånberg, E. Altered sensitivity to ellagic acid in neuroblastoma cells undergoing differentiation with 12-O-tetradecynoylphorbol-13-acetate and all-trans retinoic acid. Biomed. Pharmacother. 2015, 76, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Q.; Liu, B.; Li, Y.; Tan, Y.; Yang, B. Ellagic acid inhibits proliferation and induces apoptosis in human glioblastoma cells. Acta Cir. Bras. 2016, 31, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Farbood, Y.; Sarkaki, A.; Dolatshahi, M.; Mansouri, S.M.T.; Khodadadi, A. Ellagic acid protects the brain against 6-hydroxydopamine induced neuroinflammation in a rat model of Parkinson’s disease. Basic Clin. Neurosci. 2015, 6, 15–22. [Google Scholar]
- Khatri, D.; Juvekar, A. Abrogation of locomotor impairment in a rotenone-induced Drosophila melanogaster and zebrafish model of Parkinson’s disease by ellagic acid and curcumin. Int. J. Nutr. Pharm. Neurol. Dis. 2016, 6, 90–96. [Google Scholar]
- Mansouri, M.T.; Farbood, Y.; Naghizadeh, B.; Shabani, S.; Mirshekar, M.A.; Sarkaki, A. Beneficial effects of ellagic acid against animal models of scopolamine- and diazepam-induced cognitive impairments. Pharm. Biol. 2016, 54, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.P.; Ren, T.W.; Yan, W.J.; Liu, J.; Liu, R.B. Ellagic acid alleviates inflammatory pain and paclitaxel-induced neuropathic pain in murine models. Int. J. Clin. Exp. Med. 2016, 9, 12514–12520. [Google Scholar]
- Chen, S.Y.; Zheng, K.; Wang, Z.Q. Neuroprotective effects of ellagic acid on neonatal hypoxic brain injury via inhibition of inflammatory mediators and down-regulation of JNK/p38 MAPK activation. Trop. J. Pharm. Res. 2016, 15, 241–251. [Google Scholar] [CrossRef]
- Liu, Q.S.; Li, S.R.; Li, K.; Li, X.; Yin, X.; Pang, Z. Ellagic acid improves endogenous neural stem cells proliferation and neurorestoration through Wnt/β-catenin signaling in vivo and in vitro. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K. Occurrence and content of hydroxycinnamic and hydroxybenzoic acid compounds in foods. Crit. Rev. Sci. Food Nutr. 1989, 28, 315–347. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [PubMed]
- Bourne, L.C.; Rice-Evans, C. Bioavailablility of ferulic acid. Biochem. Biophys. Res. Commun. 1998, 253, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.C.; et al. Dietary polyphenol intake in Europe: The European prospective investigation into cancer and nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Tresserra-Rimbau, A.; Medina-Remón, A.; Pérez-Jiménez, J.; Martínez-González, M.A.; Covas, M.I.; Corella, D.; Salas-Salvadó, J.; Gómez-Gracia, E.; Lapetra, J.; Arós, F.; et al. Dietary intake and major food sources of polyphenols in a Spanish population at high cardiovascular risk: The PREDIMED study. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Stepaniak, U.; Topor-Mądry, R.; Szafraniec, K.; Pająk, A. Estimated dietary intake and major food sources of polyphenols in the Polish arm of the HAPIEE study. Nutrition 2014, 30, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Ferruzzi, M.G.; Lobo, J.K.; Janle, E.M.; Cooper, B.; Simon, J.E.; Wu, Q.L.; Welch, C.; Ho, L.; Weaver, C.; Pasinetti, G.M. Bioavailability of gallic acid and catechins from grape seed polyphenol extract is improved by repeated dosing in rats: implications for treatment in Alzheimer’s disease. J. Alzheimers Dis. 2009, 18, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Wu, L.; Zhang, Q.L.; Li, J.; Yin, F.X.; Yuan, Y. Pharmacokinetics of phenolic compounds of Danshen extract in rat blood and brain by microdialysis sampling. J. Ethnopharmacol. 2011, 136, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Margalef, M.; Pons, Z.; Bravo, F.I.; Muguerza, B.; Arola-Arnal, A. Tissue distribution of rat flavanol metabolites at different doses. J. Nutr. Biochem. 2015, 26, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Del Bo, C.; Ciapellano, S.; Klimis-Zacas, D.; Martini, D.; Gardana, C.; Riso, P.; Porrini, M. Anthocyanin absorption, metabolism, and distribution from a wild blueberry-enriched diet (Vaccinium angustifolium) is affected by diet duration in the Sprague-Dawley rat. J. Agric. Food Chem. 2010, 58, 2491–2497. [Google Scholar] [PubMed]
- Wang, D.; Ho, L.; Faith, J.; Ono, K.; Janle, E.M.; Lachcik, P.J.; Cooper, B.R.; Jannasch, A.H.; D’Arcy, B.R.; Williams, B.A.; et al. Role of intestinal microbiota in the generation of polyphenol derived phenolic acid mediated attenuation of Alzheimer’s disease β-amyloid oligomerization. Mol. Nutr. Food. Res. 2015, 59, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.X.; Xu, L.Y.; Tao, J.S.; Feng, Y. Metabolism and pharmacokinetics of ferulic acid in rats. China J. Chin. Mater. Medica 1993, 18, 300–2319. [Google Scholar]
- Wu, K.; Wang, Z.Z.; Liu, D.; Qi, X.R. Pharmacokinetics, brain distribution, release and blood-brain barrier transport of Shunaoxin pills. J. Ethnopharmacol. 2014, 151, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro Fernandes, F.D.; Fontenele Menezes, A.P.; de Sousa Neves, J.C.; Fonteles, A.A.; da Silva, A.T.; de Araújo, R.P.; Santos do Carmo, M.R.; de Souza, C.M.; de Andrade, G.M. Caffeic acid protects mice from memory deficits induced by focal cerebral ischemia. Behav. Pharmacol. 2014, 25, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Ritschel, W.A.; Starzacher, A.; Sabouni, A.; Hussain, A.S.; Koch, H.P. Percutaneous absorption of rosmarinic acid in the rat. Method. Find. Exp. Clin. Pharmacol. 1989, 11, 345–352. [Google Scholar]
- Yan, J.J.; Cho, J.Y.; Kim, H.S.; Kim, K.L.; Jung, J.S.; Huh, S.O.; Suh, H.W.; Kim, Y.H.; Song, D.K. Protection against b-amyloid peptide toxicity in vivo with long-term administration of ferulic acid. Br. J. Pharmacol. 2001, 133, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Pati, S.; Redell, J.B.; Zhang, M.; Moore, A.N.; Dash, P.K. Caffeic acid phenethyl ester protects blood-brain barrier integrity and reduces contusion volume in rodent models of traumatic brain injury. J. Neurotraum. 2012, 29, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Vafeiadou, K.; Corona, G.; Pollard, S.E.; Tzounis, X.; Spencer, J.P. Champagne wine polyphenols protect primary cortical neurons against peroxynitrite-induced injury. J. Agric. Food Chem. 2007, 55, 2854–2860. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.; Kan, Z.; Xu, Y.; Lv, C.; Jiang, W. Rosmarinic acid protects against experimental diabetes with cerebral ischemia: relation to inflammation response. J. Neuroinflamm. 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Tarozzi, A.; Morroni, F.; Hrelia, S.; Angeloni, C.; Marchesi, A.; Cantelli-Forti, G.; Hrelia, P. Neuroprotective effects of anthocyanins and their in vivo metabolites in SH-SY5Y cells. Neurosci. Lett. 2007, 424, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Scapagnini, G.; Foresti, R.; Calabrese, V.; Giuffrida Stella, A.M.; Green, C.J.; Motterlini, R. Caffeic acid phenethyl ester and curcumin: A novel class of heme oxygenase-1 inducers. Mol. Pharmacol. 2002, 3, 554–561. [Google Scholar] [CrossRef]
- Scapagnini, G.; Butterfield, D.A.; Colombrita, C.; Sultana, R.; Pascale, A.; Calabrese, V. Ethyl ferulate, a lipophilic polyphenol, induces HO-1 and protects rat neurons against oxidative stress. Antioxid. Redox Signal. 2004, 6, 811–818. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Ferulic Acid | Antidepressant-like Effect:
|
| Caffeic Acid | Inflammation:
|
| Caffeic Acid Phenethyl Ester | Antioxidant Activity:
|
| Chlorogenic Acid | Reversing of the Glutamate-induced Toxicity:
|
| Chlorogenic Acid and Its Metabolites | Reversing of the Glutamate-induced Toxicity:
|
| Rosmarinic Acid | Antioxidant Activity:
|
| P-Coumaric Acid | Protection from Ischemia/ Reperfusion Injury:
|
| Sinapic Acid | PD:
|
| Cinnamic Aldehyde | Inflammation and Cognition:
|
| Salicylic Acid | Antioxidant Activity:
|
| Acetylsalicylic Acid | Inflammation and Antioxidant Activity:
|
| Protocatechuic Acid | Antioxidant Activity:
|
| Gallic Acid | Antioxidant Activity:
|
| Tannic Acid | Antioxidant Activity:
|
| Homovanillic Acid | Antidepressant Effect:
|
| Syringic Acid | Protection from Ischemia/reperfusion Injury:
|
| Ellagic Acid | Anticancer Activity:
|
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szwajgier, D.; Borowiec, K.; Pustelniak, K. The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action. Nutrients 2017, 9, 477. https://doi.org/10.3390/nu9050477
Szwajgier D, Borowiec K, Pustelniak K. The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action. Nutrients. 2017; 9(5):477. https://doi.org/10.3390/nu9050477
Chicago/Turabian StyleSzwajgier, Dominik, Kamila Borowiec, and Katarzyna Pustelniak. 2017. "The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action" Nutrients 9, no. 5: 477. https://doi.org/10.3390/nu9050477
APA StyleSzwajgier, D., Borowiec, K., & Pustelniak, K. (2017). The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action. Nutrients, 9(5), 477. https://doi.org/10.3390/nu9050477