Combined Cytotoxicity of the Phycotoxin Okadaic Acid and Mycotoxins on Intestinal and Neuroblastoma Human Cell Models


Abstract
:
1. Introduction
2. Results
2.1. Individual Cytotoxicity of Algal Toxin and Mycotoxin
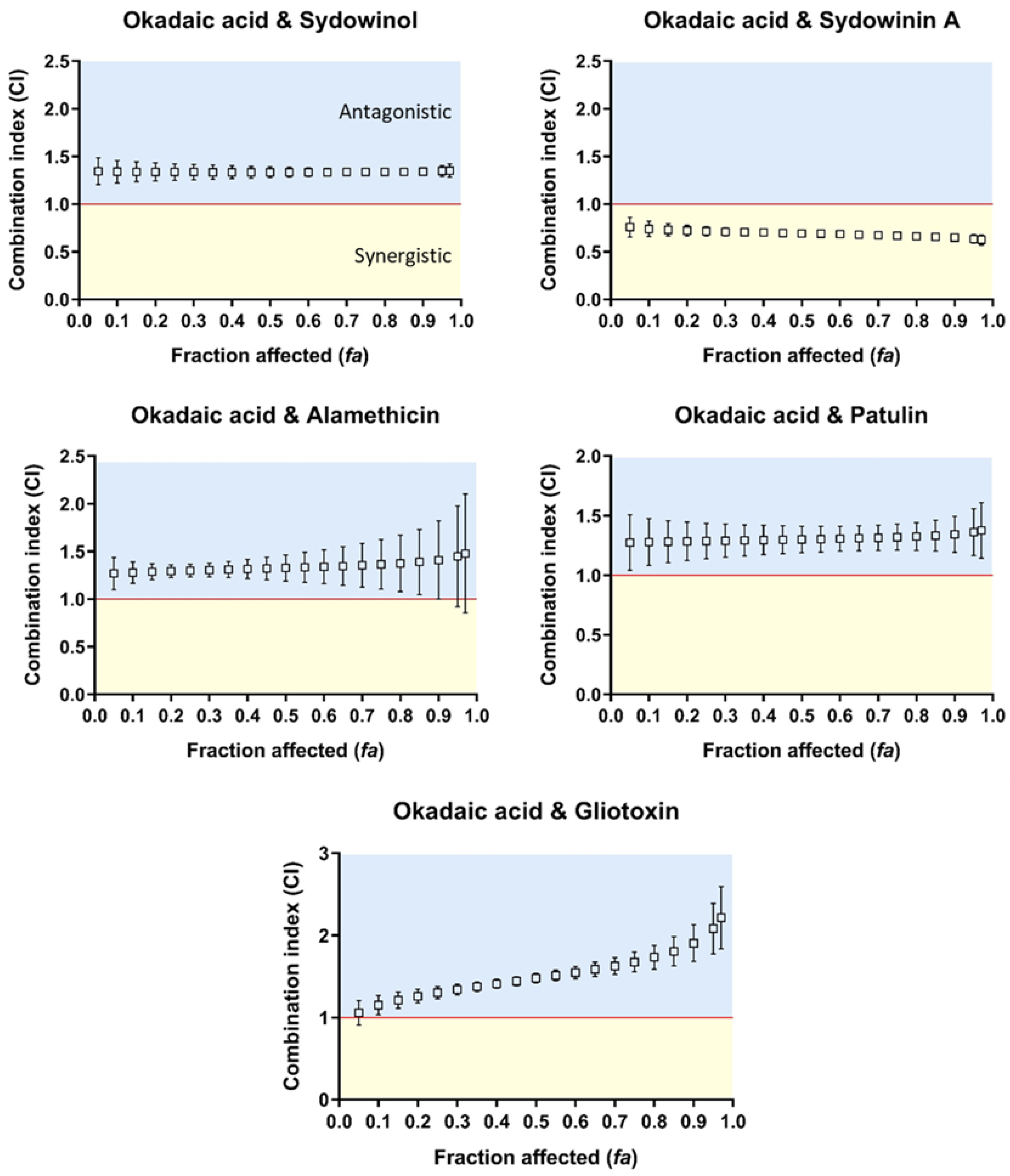
2.2. Combined Cytotoxicity of Mycotoxins and Algal Toxin
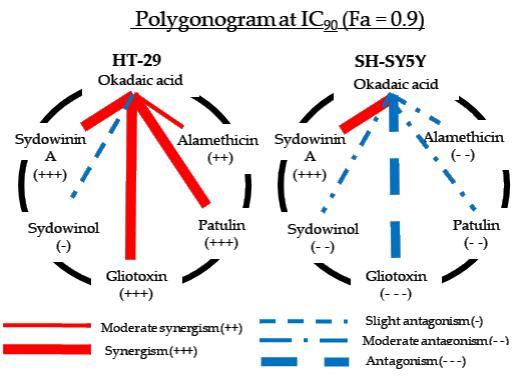
2.3. Okadaic Acid and Mycotoxins on Human Intestinal HT-29 Cells
2.4. Okadaic Acid and Mycotoxins on Human Neuroblastoma SH-SY5Y Cells
3. Discussion
3.1. Synergisms between Okadaic Acid and Mycotoxins
3.2. Antagonisms between Okadaic Acid and Mycotoxin on SH-SY5Y
4. Conclusions
5. Materials and Methods
5.1. Cell Line Cultures
5.2. Mycotoxin and Phycotoxin Toxins
5.3. Cytotoxicity Bioassays
5.4. Statistical Analysis of Cytotoxicity of Individual Mycotoxin and Algal Toxin
5.5. Median Effect and Combination Index Analysis of Mycotoxin and Algal Toxin Mixture
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sarmiento-Ramírez, J.M.; Abella, E.; Martín, M.P.; Tellería, M.T.; López-Jurado, L.F.; Marco, A.; Diéguez-Uribeondo, J. Fusarium solani is responsible for mass mortalities in nests of loggerhead sea turtle, Caretta caretta, in Boavista, Cape Verde. FEMS Microbiol. Lett. 2010, 312, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.W.; Ives, L.D.; Nagelkerken, I.A.; Richie, K.B. Caribbean sea-fan mortalities. Nature 1996, 383, 487. [Google Scholar] [CrossRef]
- Grovel, O.; Pouchus, Y.F.; Verbist, J.F. Accumulation of gliotoxin, a cytotoxic mycotoxin from Aspergillus fumigatus, in blue mussel (Mytilus edulis). Toxicon 2003, 42, 297–300. [Google Scholar] [CrossRef]
- Pang, K.L.; Overy, D.P.; Jones, E.B.G.; da Luz Calado, M.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.J.; Bills, G.F. ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 1–13. [Google Scholar] [CrossRef]
- Hallegraeff, G.; Coman, F.; Davies, C.; Hayashi, A.; McLeod, D.; Slotwinski, A.; Whittock, L.; Richardson, A.J. Australian Dust Storm Associated with Extensive Aspergillus sydowii Fungal “Bloom” in Coastal Waters. Appl. Environ. Microbiol. 2014, 80, 3315–3320. [Google Scholar] [CrossRef] [PubMed]
- Harvell, C.D.; Kim, K.; Burkholder, J.M.; Colwell, R.R.; Epstein, P.R.; Grimes, D.J.; Hofmann, E.E.; Lipp, E.K.; Osterhaus, A.D.; Overstreet, R.M.; et al. Emerging marine diseases—Climate links and anthropogenic factors. Science 1999, 285, 1505–1510. [Google Scholar] [CrossRef]
- Sallenave-Namont, C.; Pouchus, Y.F.; Robiou Du Pont, T.; Lassus, P.; Verbist, J.F. Toxigenic saprophytic fungi in marine shellfish farming areas. Mycopathologia 2000, 149, 21–25. [Google Scholar] [CrossRef]
- Brewer, D.; Greenwell, M.; Taylor, A. Studies of trichoderma isolates from mytilus edulis collected on the shores of Cape Breton and Prince Edward islands. Proc. N. S. Inst. Sci. 1993, 40, 29–40. [Google Scholar]
- Matallah-Boutiba, A.; Ruiz, N.; Sallenave-Namont, C.; Grovel, O.; Amiard, J.C.C.; Pouchus, Y.F.; Boutiba, Z. Screening for toxigenic marine-derived fungi in Algerian mussels and their immediate environment. Aquaculture 2012, 342–343, 75–79. [Google Scholar] [CrossRef]
- Zvereva, L.V.; Vysotskaya, M.A. Filamentous fungi associated with bivalve mollusks from polluted biotopes of Ussuriiskii Bay, Sea of Japan. Russ. J. Mar. Biol. 2005, 31, 382–385. [Google Scholar] [CrossRef]
- Santos, A.; Hauser-Davis, R.A.; Santos, M.J.S.; De Simone, S.G. Potentially toxic filamentous fungi associated to the economically important Nodipecten nodosus (Linnaeus, 1758) scallop farmed in southeastern Rio de Janeiro, Brazil. Mar. Pollut. Bull. 2017, 115, 75–79. [Google Scholar] [CrossRef]
- Greco, G.; Cecchi, G.; Di Piazza, S.; Cutroneo, L.; Capello, M.; Zotti, M. Fungal characterisation of a contaminated marine environment: The case of the Port of Genoa (North-Western Italy). Webbia 2018, 7792, 1–10. [Google Scholar] [CrossRef]
- Marrouchi, R.; Benoit, E.; Le Caer, J.P.; Belayouni, N.; Belghith, H.; Molgó, J.; Kharrat, R. Toxic C17-Sphinganine Analogue Mycotoxin, Contaminating Tunisian Mussels, Causes Flaccid Paralysis in Rodents. Mar. Drugs 2013, 11, 4724–4740. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.A.; Naehrer, K.; Santos, G.A. Occurrence of mycotoxins in commercial aquafeeds in Asia and Europe: A real risk to aquaculture? Rev. Aquac. 2016, 1–18. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Vansteelandt, M.; Kerzaon, I.; Blanchet, E.; Fossi Tankoua, O.; Robiou Du Pont, T.; Joubert, Y.; Monteau, F.; Le Bizec, B.; Frisvad, J.C.; Pouchus, Y.F.; et al. Patulin and secondary metabolite production by marine-derived Penicillium strains. Fungal Biol. 2012, 116, 954–961. [Google Scholar] [CrossRef]
- Poirier, L.; Montagu, M.; Landreau, A.; Mohamed-Benkada, M.; Grovel, O.; Sallenave-Namont, C.; Biard, J.F.; Amiard-Triquet, C.; Amiard, J.C.; Pouchus, Y.F. Peptaibols: Stable Markers of Fungal Development in the Marine Environment. Chem. Biodivers. 2007, 4, 1116–1128. [Google Scholar] [CrossRef]
- Petit, K.E.; Mondeguer, F.; Roquebert, M.F.; Biard, J.F.F.; Pouchus, Y.F. Detection of griseofulvin in a marine strain of Penicillium waksmanii by ion trap mass spectrometry. J. Microbiol. Methods 2004, 58, 59–65. [Google Scholar] [CrossRef]
- Sallenave, C.; Pouchus, Y.F.; Bardouil, M.; Lassus, P.; Roquebert, M.F.; Verbist, J.F. Bioaccumulation of mycotoxins by shellfish: Contamination of mussels by metabolites of a Trichoderma koningii strain isolated in the marine environment. Toxicon 1999, 37, 77–83. [Google Scholar] [CrossRef]
- Ruiz, N.; Petit, K.; Vansteelandt, M.; Kerzaon, I.; Baudet, J.; Amzil, Z.; Biard, J.F.; Grovel, O.; Pouchus, Y.F. Enhancement of domoic acid neurotoxicity on Diptera larvae bioassay by marine fungal metabolites. Toxicon 2010, 55, 805–810. [Google Scholar] [CrossRef]
- Campbell, A.; Hudson, D.; McLeod, C.; Nicholls, C.; Pointon, A. Tactical Research Fund: Review of the 2012 paralytic shellfish toxin event in Tasmania associated with the dinoflagellate alga, Alexandrium tamarense. In FRDC Project 2012/060 Appendix to the Final Report; SafeFish: Adelaide, Australia, 2013. [Google Scholar]
- Stobo, L.A.; Lacaze, J.P.C.L.; Scott, A.C.; Petrie, J.; Turrell, E.A. Surveillance of algal toxins in shellfish from Scottish waters. Toxicon 2008, 51, 635–648. [Google Scholar] [CrossRef]
- Gonçalves, R.A.; Schatzmayr, D.; Hofstetter, U.; Santos, G.A. Occurrence of mycotoxins in aquaculture: Preliminary overview of Asian and European plant ingredients and finished feeds. World Mycotoxin J. 2017, 10, 183–194. [Google Scholar] [CrossRef]
- Hayashi, A.; Crombie, A.; Lacey, E.; Richardson, A.; Vuong, D.; Piggott, A.; Hallegraeff, G. Aspergillus Sydowii Marine Fungal Bloom in Australian Coastal Waters, Its Metabolites and Potential Impact on Symbiodinium Dinoflagellates. Mar. Drugs 2016, 14, 59. [Google Scholar] [CrossRef]
- Gessner, B.D.; Middaugh, J.P. Paralytic shellfish poisoning in alaska: A 20-year retrospective analysis. Am. J. Epidemiol. 1995, 141, 766–770. [Google Scholar] [CrossRef]
- Teitelbaum, J.S.; Zatorre, R.J.; Carpenter, S.; Gendron, D.; Evans, A.C.; Gjedde, A.; Cashman, N.R. Neurologic Sequelae of Domoic Acid Intoxication Due to the Ingestion of Contaminated Mussels. N. Engl. J. Med. 1990. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pásaro, E.; Méndez, J.; Laffon, B. Okadaic Acid: More than a diarrheic toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Chou, T.C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization (FAO). Marine biotoxins. In FAO Food and Nutrition; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; p. 80. [Google Scholar]
- Wang, J.; Wang, Y.Y.; Lin, L.; Gao, Y.; Hong, H.S.; Wang, D.Z. Quantitative proteomic analysis of okadaic acid treated mouse small intestines reveals differentially expressed proteins involved in diarrhetic shellfish poisoning. J. Proteomics 2012, 75, 2038–2052. [Google Scholar] [CrossRef]
- Mueller, P.; Rudin, D.O. Action potentials induced in biomolecular lipid membranes. Nature 1968, 217, 713–719. [Google Scholar] [CrossRef]
- Jones, R.W.; Hancock, J.G. Mechanism of Gliotoxin Action and Factors Mediating Gliotoxin Sensitivity. J. Gen. Microbiol. 1988, 134, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Riley, R.T.; Showker, J.L. The Mechanism of Patulin’s Cytotoxicity and the Antioxidant Activity of lndole Tetramic Acids. Toxicol. Appl. Pharmacol. 1991, 126, 108–126. [Google Scholar] [CrossRef]
- Fessard, V.; Alarcan, J.; Barbé, S.; Kopp, B.; Hessel-Pras, S.; Lampen, A.; Le Hégarat, L. In vitro assessment of binary mixtures effects of phycotoxins in human intestinal cells. In Proceedings of the 18th International Conference on Harmful Algae, Nantes, France, 21–26 October 2018; p. 468. [Google Scholar]
- Liu, H.; Chen, S.; Liu, W.; Liu, Y.; Huang, X.; She, Z. Polyketides with immunosuppressive activities from mangrove endophytic fungus Penicillium sp. ZJ-SY2. Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef]
- Tapia, R.; Peña, F.; Arias, C. Neurotoxic and synaptic effects of okadaic acid, an inhibitor of protein phosphatases. Neurochem. Res. 1999, 24, 1423–1430. [Google Scholar] [CrossRef]
- Arias, C.; Sharma, N.; Davies, P.; Shafit-Zagardo, B. Okadaic acid induces early changes in microtubule-associated protein 2 and tau phosphorylation prior to neurodegeneration in cultured cortical neurons. J. Neurochem. 1993, 61, 673–682. [Google Scholar] [CrossRef]
- Matias, W.G.; Traore, A.; Creppy, E.E. Variations in the distribution of okadaic acid in organs and biological fluids of mice related to diarrhoeic syndrome. Hum. Exp. Toxicol. 1999, 18, 345–350. [Google Scholar] [CrossRef]
- Lum, B.L.; Gosland, M.P. MDR expression in normal tissues. Pharmacologic implications for the clinical use of P-glycoprotein inhibitors. Hematol. Oncol. Clin. N. Am. 1995, 9, 319–336. [Google Scholar] [CrossRef]
- Tohda, H.; Yasui, A.; Yasumoto, T.; Nakayasu, M.; Shima, H.; Nagao, M.; Sugimura, T. Chinese hamster ovary cells resistant to okadaic acid express a multidrug resistant phenotype. Biochem. Biophys. Res. Commun. 1994, 203, 1210–1216. [Google Scholar] [CrossRef]
- Breuer, W.; Slotki, I.N.; Ausiello, D.A.; Cabantchik, I.Z. Induction of multidrug resistance downregulates the expression of CFTR in colon epithelial cells. Am. J. Physiol. 1993, 265, C1711–C1715. [Google Scholar] [CrossRef]
- Bates, S.E.; Shieh, C.Y.; Tsokos, M. Expression of mdr-1/P-glycoprotein in human neuroblastoma. Am. J. Pathol. 1991, 139, 305–315. [Google Scholar]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef]
- Chou, T.C. Drug combination studies and their synergy quantification using the chou-talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]
- Hamasaki, T.; Sato, Y.; Hatsuda, Y. Structure of Sydowinin A, Sydowinin B, and Sydowinol, Metabolites from Aspergillus sydowi. Agric. Biol. Chem. 1975, 39, 2341–2345. [Google Scholar] [CrossRef]
- Hamasaki, T.; Sato, Y.; Hatsuda, Y. Isolation of new metabolites from Aspergillus sydowi and structure of sydowic acid. Agric. Biol. Chem. 1975, 39, 2337–2340. [Google Scholar] [CrossRef]
- O’Brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef] [Green Version]
- Sérandour, A.L.; Ledreux, A.; Morin, B.; Derick, S.; Augier, E.; Lanceleur, R.; Hamlaoui, S.; Moukha, S.; Furger, C.; Biré, R.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part I: Screening strategy and pre-validation study with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1983–1993. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | HT-29 | 95% CI | SH-SY5Y | 95% CI |
|---|---|---|---|---|
| IC50 (µM) | IC50 (µM) | |||
| Typical A. sydowii metabolites | ||||
| Sydowinin A | 124.30 | 113.60–136.00 | 117.80 | 105.60–131.40 |
| Sydowinin B | 93.06 | 82.20–105.40 | 143.8 | 116.00–178.20 |
| Sydowinol | 2.50 | 2.21–2.82 | 5.14 | 5.06–5.23 |
| Sydowic acid | NE (283.75) 1 | - | NE (283.75) 1 | - |
| Dust storm/shellfish mycotoxins | ||||
| Sterigmatocystin | >61.67 2 | - | ~123.35 2 | - |
| Patulin | 17.46 | 10.79–28.28 | 2.23 | 2.15–2.32 |
| Alamethicin | 4.92 | 4.57–5.29 | 5.43 | 5.29–5.67 |
| Gliotoxin | 0.062 | 0.052–0.075 | 0.045 | 0.039–0.053 |
| Algal toxins | ||||
| Okadaic acid | 0.065 | 0.056–0.075 | 0.027 | 0.026–0.029 |
| Toxin Mixture | Molar Combination Ratio | |
|---|---|---|
| HT-29 | SH-SY5Y | |
| Okadaic acid:Sydowinin A | 1:1925.0 | 1:14366.2 |
| Okadaic acid:Sydowinol | 1:38.7 | 1:190.7 |
| Okadaic acid:Alamethicin | 1:76.6 | 1:201.1 |
| Okadaic acid:Patulin | 1:270.4 | 1:82.6 |
| Okadaic acid:Gliotoxin | 1:1.04 | 1:1.68 |
| Toxin Mixture | CI at | DRI at | ||||||
|---|---|---|---|---|---|---|---|---|
| IC25 | IC50 | IC75 | IC90 | IC25 | IC50 | IC75 | IC90 | |
| HT-29 | ||||||||
| Okadaic acid | 1.67 | 1.24 | 0.93 | 0.69 | - | - | 2.72 | 3.61 |
| Sydowinin A | - | - | 1.80 | 2.43 | ||||
| Okadaic acid | 1.47 | 1.34 | 1.23 | 1.13 | - | - | - | - |
| Sydowinol | - | - | - | - | ||||
| Okadaic acid | 1.06 | 0.88 | 0.78 | 0.72 | - | 2.41 | 2.18 | 1.98 |
| Alamethicin | - | 2.14 | 3.14 | 4.61 | ||||
| Okadaic acid | 1.53 | 1.01 | 0.75 | 0.53 | - | - | 1.76 | 1.95 |
| Patulin | - | - | 5.63 | 12.23 | ||||
| Okadaic acid | 2.84 | 1.42 | 0.75 | 0.41 | - | - | 4.80 | 12.45 |
| Gliotoxin | - | - | 1.85 | 3.02 | ||||
| SH-SY5Y | ||||||||
| Okadaic acid | 0.72 | 0.69 | 0.67 | 0.65 | 2.98 | 3.09 | 3.20 | 3.32 |
| Sydowinin A | 2.65 | 2.72 | 2.79 | 2.86 | ||||
| Okadaic acid | 1.34 | 1.33 | 1.34 | 1.34 | - | - | - | - |
| Sydowinol | - | - | - | - | ||||
| Okadaic acid | 1.30 | 1.33 | 1.36 | 1.41 | - | - | - | - |
| Alamethicin | - | - | - | - | ||||
| Okadaic acid | 1.29 | 1.30 | 1.32 | 1.34 | - | - | - | - |
| Patulin | - | - | - | - | ||||
| Okadaic acid | 1.30 | 1.48 | 1.68 | 1.91 | - | - | - | - |
| Gliotoxin | - | - | - | - | ||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, A.; José Dorantes-Aranda, J.; Bowman, J.P.; Hallegraeff, G. Combined Cytotoxicity of the Phycotoxin Okadaic Acid and Mycotoxins on Intestinal and Neuroblastoma Human Cell Models. Toxins 2018, 10, 526. https://doi.org/10.3390/toxins10120526
Hayashi A, José Dorantes-Aranda J, Bowman JP, Hallegraeff G. Combined Cytotoxicity of the Phycotoxin Okadaic Acid and Mycotoxins on Intestinal and Neuroblastoma Human Cell Models. Toxins. 2018; 10(12):526. https://doi.org/10.3390/toxins10120526
Chicago/Turabian StyleHayashi, Aiko, Juan José Dorantes-Aranda, John P. Bowman, and Gustaaf Hallegraeff. 2018. "Combined Cytotoxicity of the Phycotoxin Okadaic Acid and Mycotoxins on Intestinal and Neuroblastoma Human Cell Models" Toxins 10, no. 12: 526. https://doi.org/10.3390/toxins10120526