Antimicrobial and Chemotactic Activity of Scorpion-Derived Peptide, ToAP2, against Mycobacterium massiliensis

 , , and
, , and 
Abstract
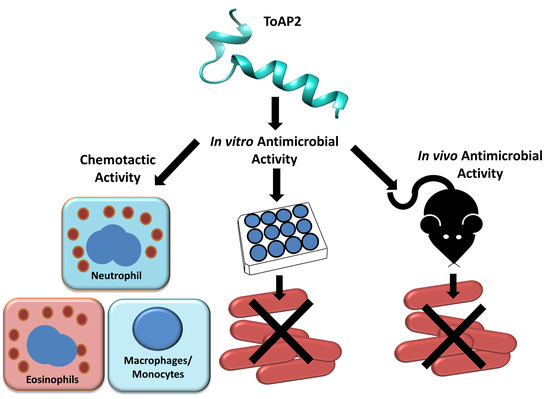
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
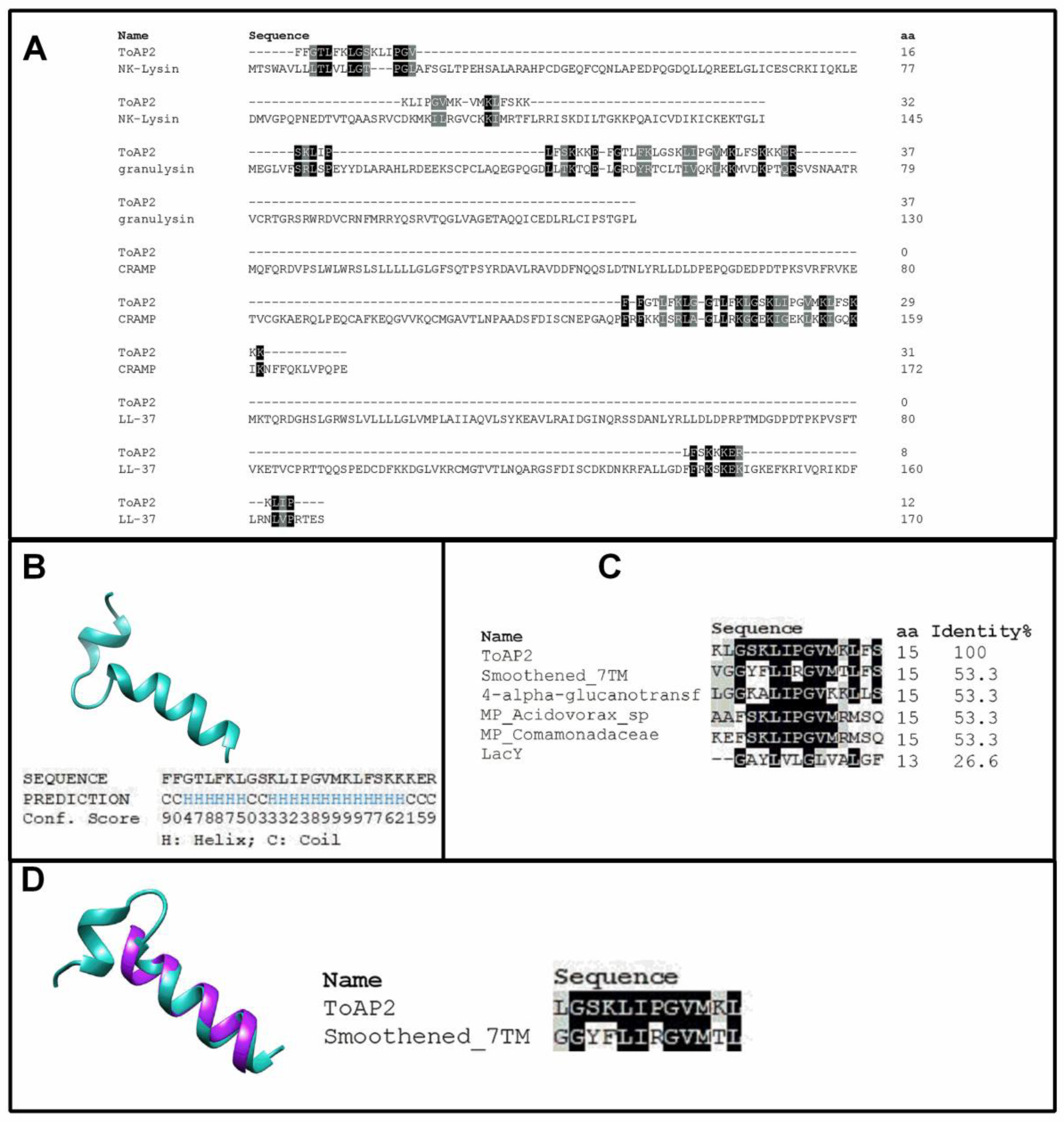
2.1. ToAP2 Secondary Structure and Similarity with Transmembrane Proteins
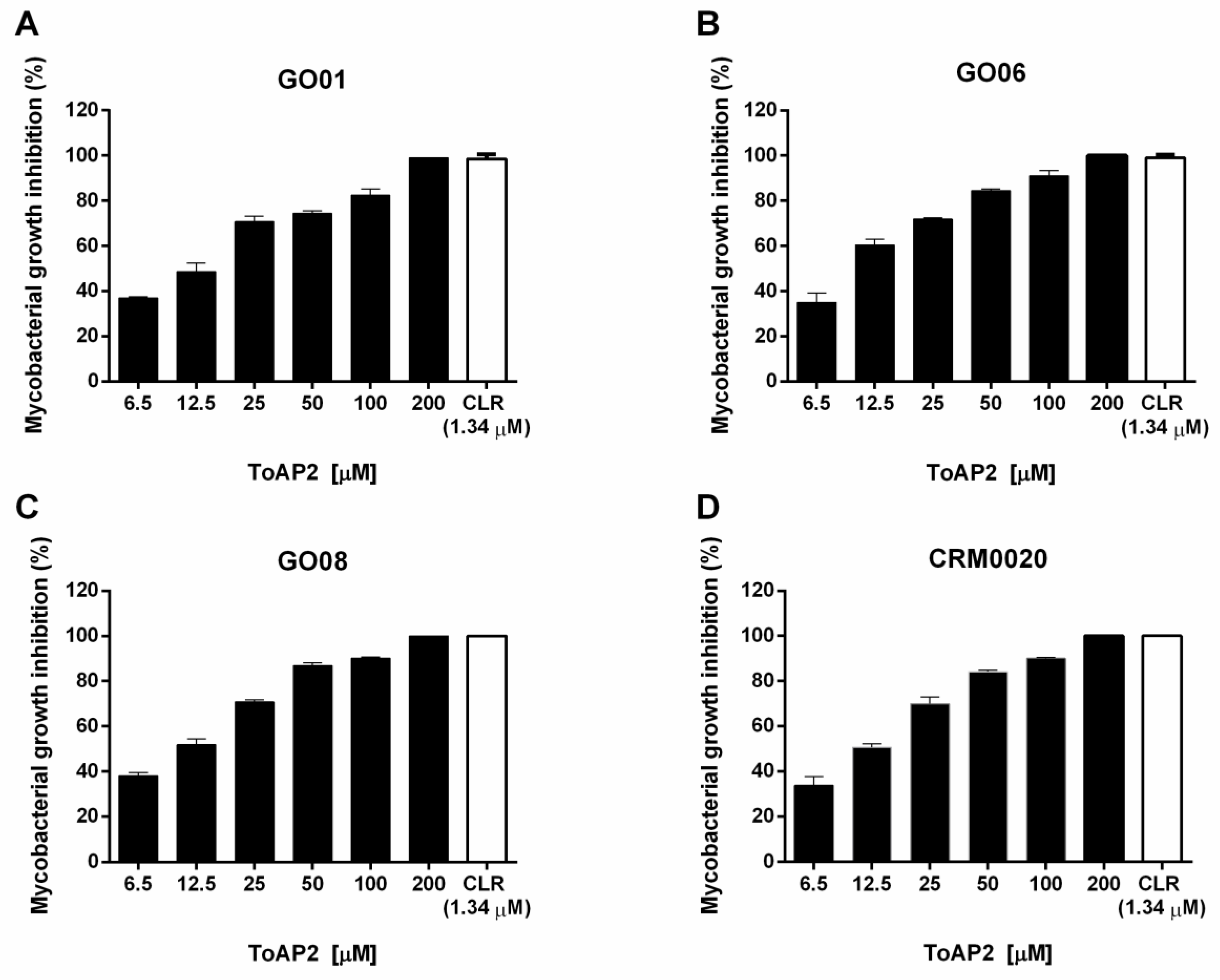
2.2. ToAP2 Antimicrobial Effect on Mycmas
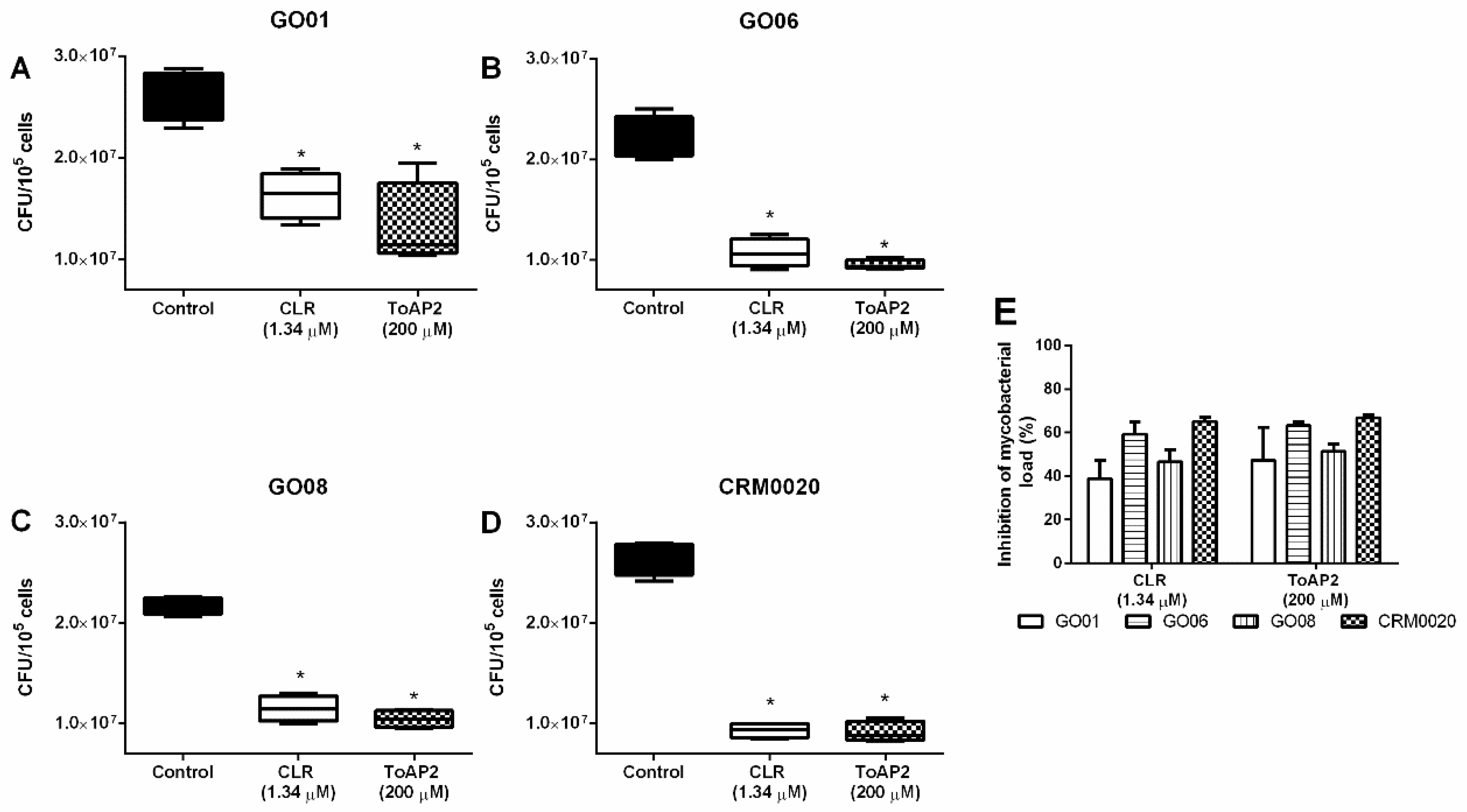
2.3. ToAP2 Antimicrobicidal Activity on Macrophages Infected with Mycmas
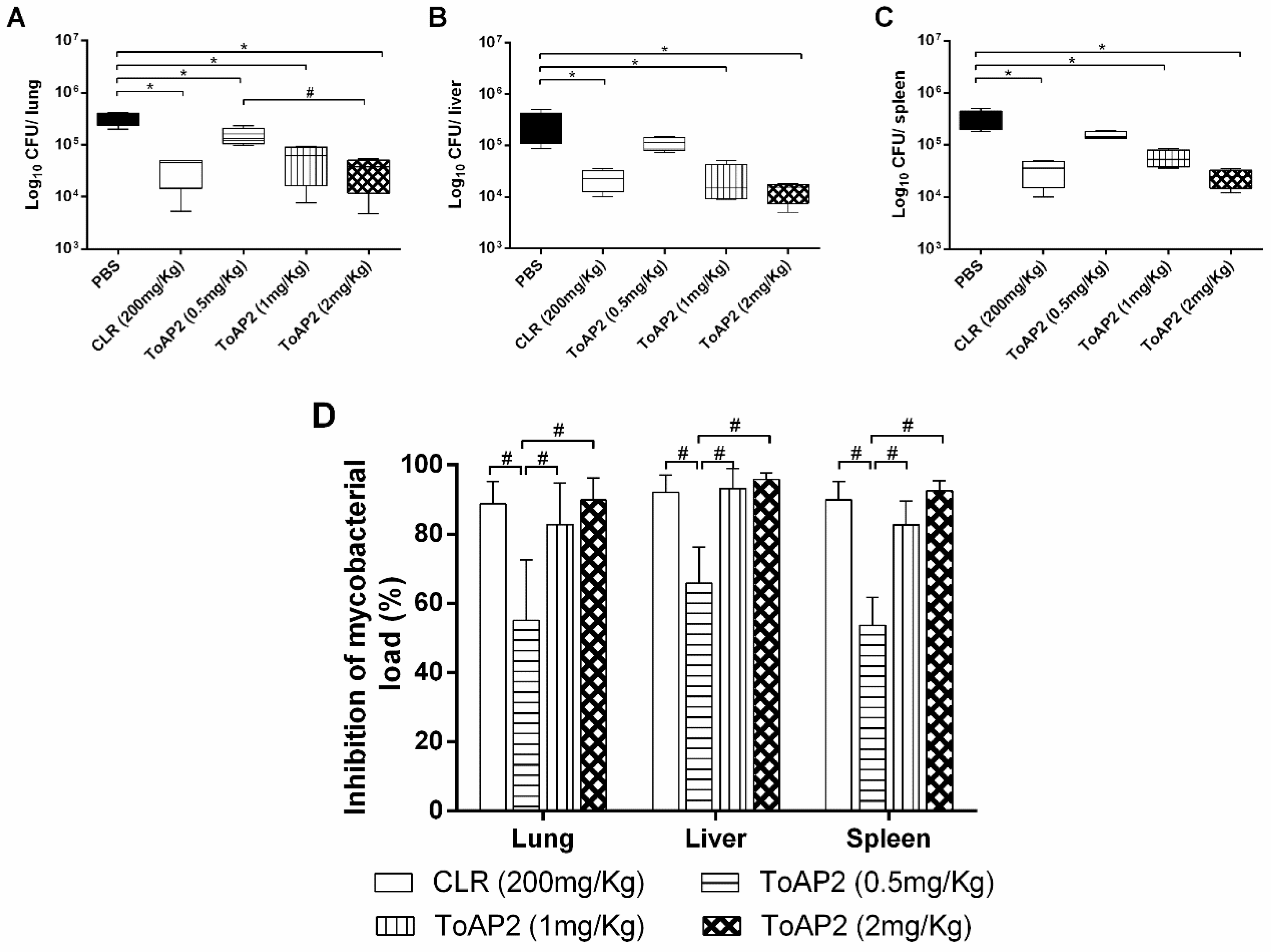
2.4. ToAP2 Reduction of Bacillary Load in Spleen, Liver, and Lung of Mice Infected with Mycmas
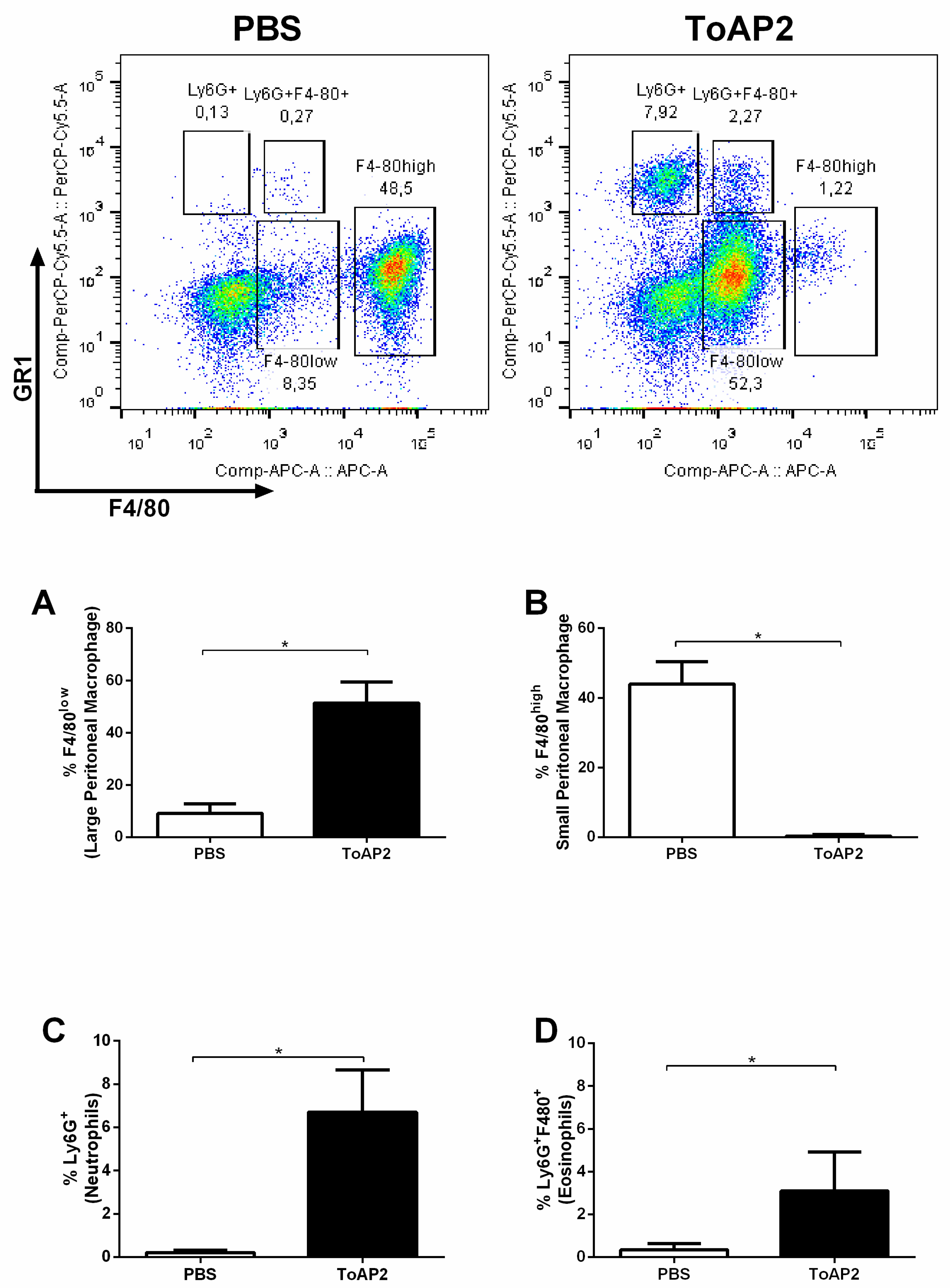
2.5. In Vivo Chemotactic Activity of ToAP2 on Monocytes
3. Discussion
4. Material and Methods
4.1. Bioinformatics Analysis
4.2. ToAP2 Synthesis and Purity
4.3. Bacterial Strain Preparation
4.4. Animals
4.5. Minimal Bactericidal Concentration (MBC) Evaluation
4.6. Antimicrobial Activity of ToAP2 in Bone Marrow Derived Macrophage Infected with Mycobacterium massiliense
4.7. In Vivo Antimicrobial Activity of ToAP2 against Mycobacterium massiliense
4.8. In Vivo Chemotactic Activity of ToAP2
4.9. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Adekambi, T.; Sassi, M.; van Ingen, J.; Drancourt, M. Reinstating Mycobacterium massiliense and Mycobacterium Bolletii as Species of the Mycobacterium Abscessus Complex. Int. J. Syst. Evol. Microbiol. 2017, 67, 2726–2730. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, A.M.; Martins de Sousa, E.; Viana-Niero, C.; Bonfim de Bortoli, F.; Pereira das Neves, Z.C.; Leao, S.C.; Junqueira-Kipnis, A.P.; Kipnis, A. Emergence of Nosocomial Mycobacterium massiliense Infection in Goias, Brazil. Microbes Infect. 2008, 10, 1552–1557. [Google Scholar] [CrossRef] [PubMed]
- Duarte, R.S.; Lourenco, M.C.; Lde, S.F.; Leao, S.C.; Ede, L.A.; Rocha, I.L.; Coelho, F.S.; Viana-Niero, C.; Gomes, K.M.; da Silva, M.G.; et al. Epidemic of Postsurgical Infections Caused by Mycobacterium Massiliense. J. Clin. Microbiol. 2009, 47, 2149–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sfeir, M.; Walsh, M.; Rosa, R.; Aragon, L.; Liu, S.Y.; Cleary, T.; Worley, M.; Frederick, C.; Abbo, L.M. Mycobacterium Abscessus Complex Infections: A Retrospective Cohort Study. Open Forum Infect. Dis. 2018, 5, ofy022. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.R.; Sheng, W.H.; Hung, C.C.; Yu, C.J.; Lee, L.N.; Hsueh, P.R. Mycobacterium Abscessus Complex Infections in Humans. Emerg. Infect. Dis. 2015, 21, 1638–1646. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.P.; Chen, Y.H.; Ming, X.; Kong, Y. Design and Synthesis of a Novel Cationic Peptide with Potent and Broad-Spectrum Antimicrobial Activity. Biomed. Res. Int. 2015, 2015, 578764. [Google Scholar] [CrossRef] [PubMed]
- Carriel-Gomes, M.C.; Kratz, J.M.; Barracco, M.A.; Bachere, E.; Barardi, C.R.; Simoes, C.M. In Vitro Antiviral Activity of Antimicrobial Peptides against Herpes Simplex Virus 1, Adenovirus, and Rotavirus. Mem. Inst. Oswaldo Cruz 2007, 102, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.; Yang, Y.; Lyu, X.; Dong, N.; Shan, A. Antimicrobial Activity, Improved Cell Selectivity and Mode of Action of Short Pmap-36-Derived Peptides against Bacteria and Candida. Sci. Rep. 2016, 6, 27258. [Google Scholar] [CrossRef] [PubMed]
- Zanger, P.; Holzer, J.; Schleucher, R.; Scherbaum, H.; Schittek, B.; Gabrysch, S. Severity of Staphylococcus Aureus Infection of the Skin Is Associated with Inducibility of Human Beta-Defensin 3 but Not Human Beta-Defensin 2. Infect. Immun. 2010, 78, 3112–3117. [Google Scholar] [CrossRef] [PubMed]
- Falcao, C.B.; de La Torre, B.G.; Perez-Peinado, C.; Barron, A.E.; Andreu, D.; Radis-Baptista, G. Vipericidins: A Novel Family of Cathelicidin-Related Peptides from the Venom Gland of South American Pit Vipers. Amino Acids 2014, 46, 2561–2571. [Google Scholar] [CrossRef] [PubMed]
- Das Neves, R.C.; Trentini, M.M.; de Castro e Silva, J.; Simon, K.S.; Bocca, A.L.; Silva, L.P.; Mortari, M.R.; Kipnis, A.; Junqueira-Kipnis, A.P. Antimycobacterial Activity of a New Peptide Polydim-I Isolated from Neotropical Social Wasp Polybia Dimorpha. PLoS ONE 2016, 11, e0149729. [Google Scholar] [CrossRef] [PubMed]
- Cerovsky, V.; Hovorka, O.; Cvacka, J.; Voburka, Z.; Bednarova, L.; Borovickova, L.; Slaninova, J.; Fucik, V. Melectin: A Novel Antimicrobial Peptide from the Venom of the Cleptoparasitic Bee Melecta Albifrons. Chembiochem 2008, 9, 2815–2821. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.L.; Abdel-Rahman, M.A.; Miller, K.; Strong, P.N. Antimicrobial Peptides from Scorpion Venoms. Toxicon 2014, 88, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial Peptides: Key Components of the Innate Immune System. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Kahlenberg, J.M.; Kaplan, M.J. Little Peptide, Big Effects: The Role of Ll-37 in Inflammation and Autoimmune Disease. J. Immunol. 2013, 191, 4895–4901. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, Z.H.; Tewary, P.; Chen, Q.; de la Rosa, G.; Oppenheim, J.J. Defensin Participation in Innate and Adaptive Immunity. Curr. Pharm. Des. 2007, 13, 3131–3139. [Google Scholar] [CrossRef] [PubMed]
- Kurosaka, K.; Chen, Q.; Yarovinsky, F.; Oppenheim, J.J.; Yang, D. Mouse Cathelin-Related Antimicrobial Peptide Chemoattracts Leukocytes Using Formyl Peptide Receptor-Like 1/Mouse Formyl Peptide Receptor-Like 2 as the Receptor and Acts as an Immune Adjuvant. J. Immunol. 2005, 174, 6257–6265. [Google Scholar] [CrossRef] [PubMed]
- Nijnik, A.; Hancock, R.E. The Roles of Cathelicidin Ll-37 in Immune Defences and Novel Clinical Applications. Curr. Opin. Hematol. 2009, 16, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Guilhelmelli, F.; Vilela, N.; Smidt, K.S.; de Oliveira, M.A.; da Cunha Morales Alvares, A.; Rigonatto, M.C.; da Silva Costa, P.H.; Tavares, A.H.; de Freitas, S.M.; Nicola, A.M.; et al. Albuquerque, and I. Silva-Pereira. Activity of Scorpion Venom-Derived Antifungal Peptides against Planktonic Cells of Candida Spp. and Cryptococcus Neoformans and Candida Albicans Biofilms. Front. Microbiol. 2016, 7, 1844. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and Arginine-Rich Antimicrobial Peptides: Structures and Mechanisms of Action. Biochim. Biophys. Acta 2006, 1758, 1184–1202. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Control of Cell Selectivity of Antimicrobial Peptides. Biochim. Biophys. Acta 2009, 1788, 1687–1692. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-Tasser Suite: Protein Structure and Function Prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Tjabringa, G.S.; Ninaber, D.K.; Drijfhout, J.W.; Rabe, K.F.; Hiemstra, P.S. Human Cathelicidin LL-37 Is a Chemoattractant for Eosinophils and Neutrophils That Acts Via Formyl-Peptide Receptors. Int. Arch. Allergy Immunol. 2006, 140, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Tjabringa, G.S.; Rabe, K.F.; Hiemstra, P.S. The Human Cathelicidin Ll-37: A Multifunctional Peptide Involved in Infection and Inflammation in the Lung. Pulm. Pharmacol. Ther. 2005, 18, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.C.; Schmidt, A.P.; Anderson, G.M.; Wang, J.M.; Wooters, J.; Oppenheim, J.J.; Chertov, O. LL-37, the Neutrophil Granule- and Epithelial Cell-Derived Cathelicidin, Utilizes Formyl Peptide Receptor-Like 1 (Fprl1) as a Receptor to Chemoattract Human Peripheral Blood Neutrophils, Monocytes, and T Cells. J. Exp. Med. 2000, 192, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Trentini, M.M.; das Neves, R.C.; Santos, B.P.; DaSilva, R.A.; de Souza, A.C.; Mortari, M.R.; Schwartz, E.F.; Kipnis, A.; Junqueira-Kipnis, A.P. Non-Disulfide-Bridge Peptide 5.5 from the Scorpion Hadrurus Gertschi Inhibits the Growth of Mycobacterium Abscessus Subsp. Massiliense. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Wang, G. The Importance of Amino Acid Composition in Natural Amps: An Evolutional, Structural, and Functional Perspective. Front. Immunol. 2012, 3, 221. [Google Scholar] [CrossRef] [PubMed]
- Campagna, S.; Saint, N.; Molle, G.; Aumelas, A. Structure and Mechanism of Action of the Antimicrobial Peptide Piscidin. Biochemistry 2007, 46, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.C.; Sun, Y.; Qian, S.; Huang, H.W. Transmembrane Pores Formed by Human Antimicrobial Peptide Ll-37. Biophys. J. 2011, 100, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Arranz-Trullen, J.; Lu, L.; Pulido, D.; Bhakta, S.; Boix, E. Host Antimicrobial Peptides: The Promise of New Treatment Strategies against Tuberculosis. Front. Immunol. 2017, 8, 1499. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, B.; Santiago, C.E.R.; Castaneda-Delgado, J.E.; Leon-Contreras, J.C.; Hancock, R.E.; Hernandez-Pando, R. Activity of Ll-37, Cramp and Antimicrobial Peptide-Derived Compounds E2, E6 and Cp26 against Mycobacterium Tuberculosis. Int. J. Antimicrob. Agents 2013, 41, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Ghosn, E.E.; Cassado, A.A.; Govoni, G.R.; Fukuhara, T.; Yang, Y.; Monack, D.M.; Bortoluci, K.R.; Almeida, S.R.; Herzenberg, L.A.; Herzenberg, L.A. Two Physically, Functionally, and Developmentally Distinct Peritoneal Macrophage Subsets. Proc. Natl. Acad. Sci. USA 2010, 107, 2568–2573. [Google Scholar] [CrossRef] [PubMed]
- Cassado Ados, A.; Lima, M.R.D.; Bortoluci, K.R. Revisiting Mouse Peritoneal Macrophages: Heterogeneity, Development, and Function. Front. Immunol. 2015, 6, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.J.; Jacobsen, E.A.; Ochkur, S.I.; McGarry, M.P.; Condjella, R.M.; Doyle, A.D.; Luo, H.; Zellner, K.R.; Protheroe, C.A.; Willetts, L.; et al. Human Versus Mouse Eosinophils: That Which We Call an Eosinophil, by Any Other Name Would Stain as Red. J. Allergy Clin. Immunol. 2012, 130, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, I.; Suzuki, K.; Niyonsaba, F.; Tamura, H.; Hirata, M. Modulation of Neutrophil Apoptosis by Antimicrobial Peptides. ISRN Microbiol. 2012, 2012, 345791. [Google Scholar] [CrossRef] [PubMed]
- Alalwani, S.M.; Sierigk, J.; Herr, C.; Pinkenburg, O.; Gallo, R.; Vogelmeier, C.; Bals, R. The Antimicrobial Peptide Ll-37 Modulates the Inflammatory and Host Defense Response of Human Neutrophils. Eur. J. Immunol. 2010, 40, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Van der Does, A.M.; Beekhuizen, H.; Ravensbergen, B.; Vos, T.; Ottenhoff, T.H.; van Dissel, J.T.; Drijfhout, J.W.; Hiemstra, P.S.; Nibbering, P.H. Ll-37 Directs Macrophage Differentiation toward Macrophages with a Proinflammatory Signature. J. Immunol. 2010, 185, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.L.; Poon, G.F.; Birkenhead, D.; Pena, O.M.; Falsafi, R.; Dahlgren, C.; Karlsson, A.; Bylund, J.; Hancock, R.E.; Johnson, P. Host Defense Peptide Ll-37 Selectively Reduces Proinflammatory Macrophage Responses. J. Immunol. 2011, 186, 5497–5505. [Google Scholar] [CrossRef] [PubMed]
- Torres-Juarez, F.; Cardenas-Vargas, A.; Montoya-Rosales, A.; Gonzalez-Curiel, I.; Garcia-Hernandez, M.H.; Enciso-Moreno, J.A.; Hancock, R.E.; Rivas-Santiago, B. Ll-37 Immunomodulatory Activity During Mycobacterium Tuberculosis Infection in Macrophages. Infect. Immun. 2015, 83, 4495–4503. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. Ucsf Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques-Neto, L.M.; Trentini, M.M.; Das Neves, R.C.; Resende, D.P.; Procopio, V.O.; Da Costa, A.C.; Kipnis, A.; Mortari, M.R.; Schwartz, E.F.; Junqueira-Kipnis, A.P. Antimicrobial and Chemotactic Activity of Scorpion-Derived Peptide, ToAP2, against Mycobacterium massiliensis. Toxins 2018, 10, 219. https://doi.org/10.3390/toxins10060219
Marques-Neto LM, Trentini MM, Das Neves RC, Resende DP, Procopio VO, Da Costa AC, Kipnis A, Mortari MR, Schwartz EF, Junqueira-Kipnis AP. Antimicrobial and Chemotactic Activity of Scorpion-Derived Peptide, ToAP2, against Mycobacterium massiliensis. Toxins. 2018; 10(6):219. https://doi.org/10.3390/toxins10060219
Chicago/Turabian StyleMarques-Neto, Lázaro M., Monalisa M. Trentini, Rogério C. Das Neves, Danilo P. Resende, Victor O. Procopio, Adeliane C. Da Costa, André Kipnis, Márcia R. Mortari, Elisabeth F. Schwartz, and Ana Paula Junqueira-Kipnis. 2018. "Antimicrobial and Chemotactic Activity of Scorpion-Derived Peptide, ToAP2, against Mycobacterium massiliensis" Toxins 10, no. 6: 219. https://doi.org/10.3390/toxins10060219