Synthesis, Structure and Biological Activity of CIA and CIB, Two α-Conotoxins from the Predation-Evoked Venom of Conus catus

 , ,
, ,
Abstract
:
1. Introduction
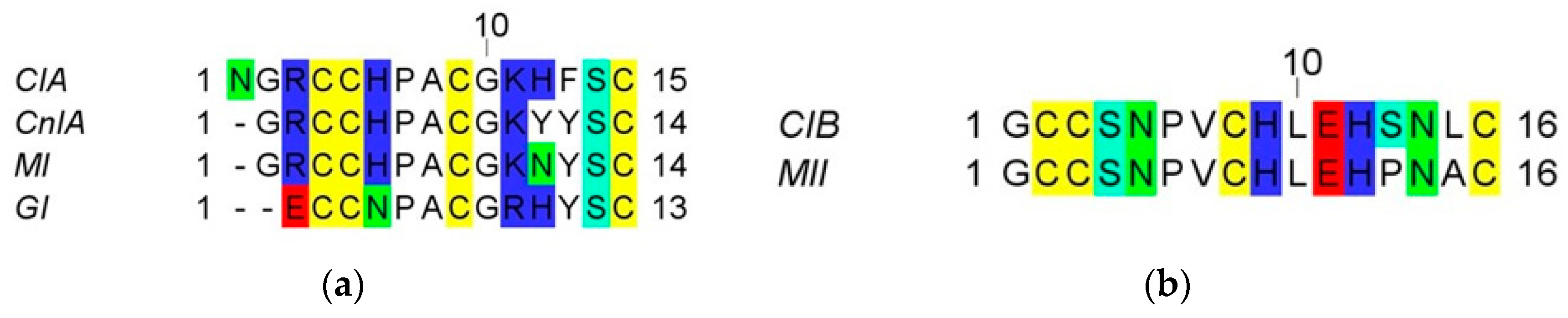
2. Results
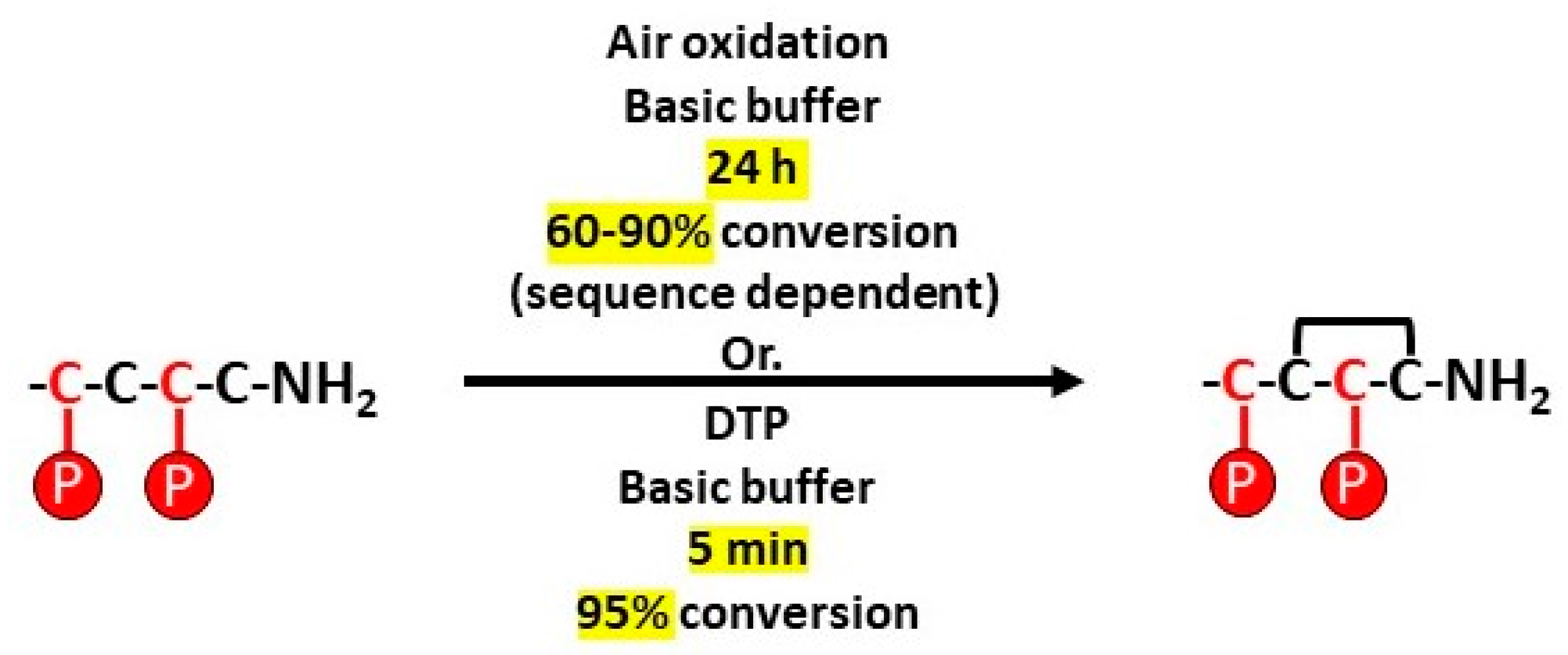
2.1. Chemical Synthesis
2.2. Electrophysiology
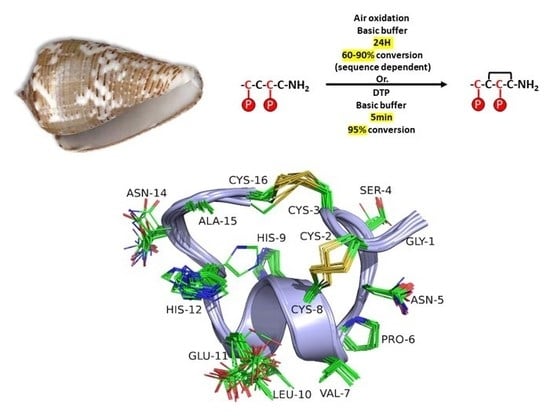
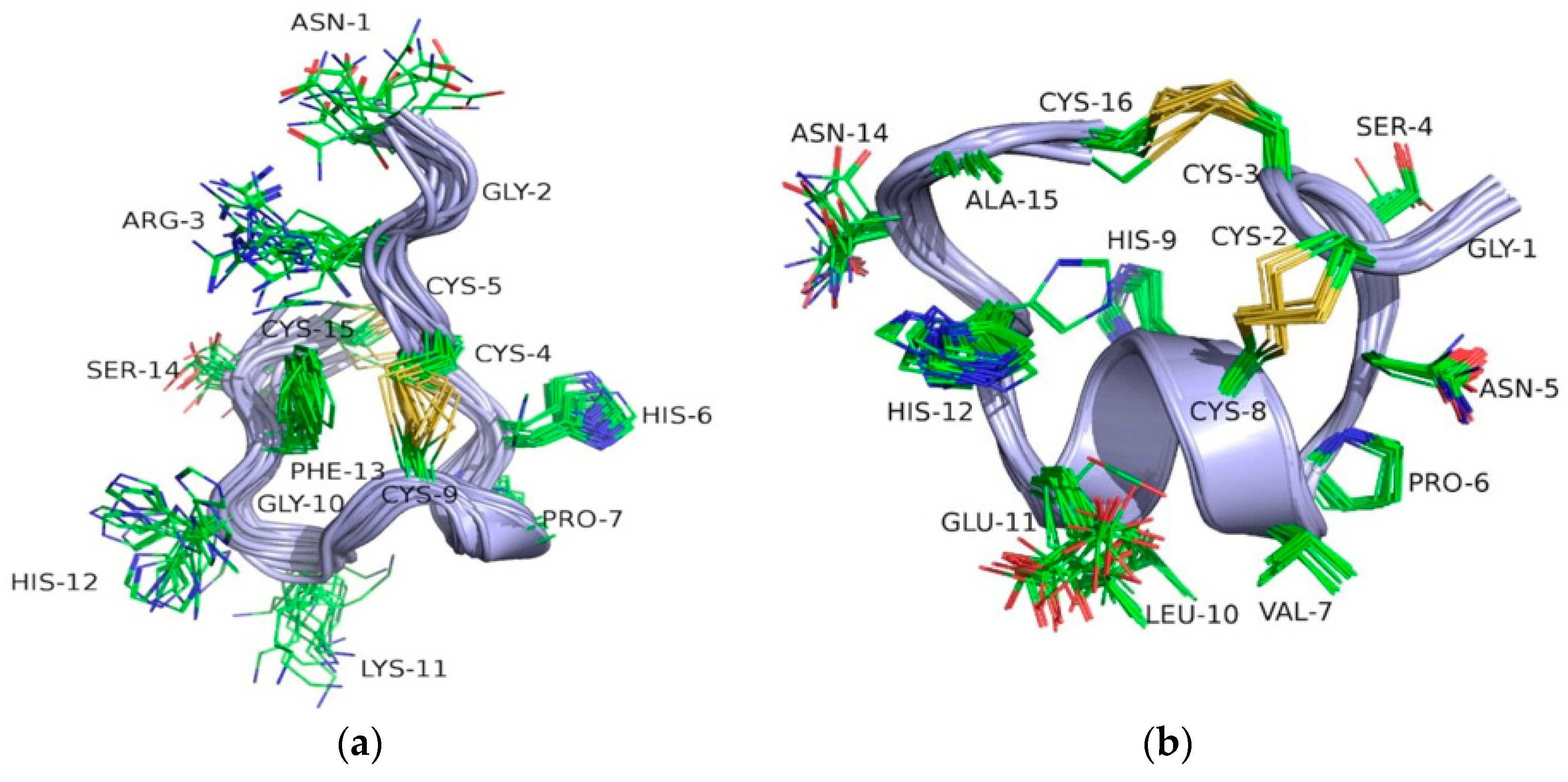
2.3. NMR Spectroscopy
2.4. In vivo Bioassays
3. Discussion
4. Materials and Methods
4.1. Abbreviations
4.2. Chemical Synthesis
4.3. Mass Spectrometry
4.4. Preparative RP-HPLC
4.5. Electrophysiology Measurements
4.6. NMR Spectroscopy
4.7. In vivo Bioassay
Author Contributions
Funding
Conflicts of Interest
References
- Puillandre, N.; Duda, T.F.; Meyer, C.; Olivera, B.M.; Bouchet, P. One, four or 100 genera? A new classification of the cone snails. J. Molluscan Stud. 2015, 81, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prashanth, J.R.; Brust, A.; Jin, A.-H.; Alewood, P.F.; Dutertre, S.; Lewis, R.J. Cone snail venomics: From novel biology to novel therapeutics. Future Med. Chem. 2014, 6, 1659–1675. [Google Scholar] [CrossRef] [PubMed]
- Sharman, J.L.; Benson, H.E.; Pawson, A.J.; Lukito, V.; Mpamhanga, C.P.; Bombail, V.; Davenport, A.P.; Peters, J.A.; Spedding, M.; Harmar, A.J. NC-IUPHAR IUPHAR-DB: Updated database content and new features. Nucleic Acids Res. 2013, 41, D1083–D1088. [Google Scholar] [CrossRef] [PubMed]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, Synthesis, and Structure–Activity Relationships of Conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Lewis, R.J. Therapeutic potential of cone snail venom peptides (conopeptides). Curr. Top. Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Terlau, H. Toxins from cone snails: Properties, applications and biotechnological production. Appl. Microbiol. Biotechnol. 2008, 79, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dutton, J.L.; Craik, D.J. Alpha-Conotoxins: Nicotinic acetylcholine receptor antagonists as pharmacological tools and potential drug leads. Curr. Med. Chem. 2001, 8, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.; Baell, J.; Norton, R. Strategies for the Development of Conotoxins as New Therapeutic Leads. Mar. Drug. 2013, 11, 2293–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide therapeutics from venom: Current status and potential. Bioorg. Med. Chem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Cooper, E.; Couturier, S.; Ballivet, M. Pentameric structure and subunit stoichiometry of a neuronal nicotinic acetylcholine receptor. Nature 1991, 350, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Twede, V.D.; Miljanich, G.; Olivera, B.M.; Bulaj, G. Neuroprotective and cardioprotective conopeptides: An emerging class of drug leads. Curr. Opin. Drug Discov. Dev. 2009, 12, 231–239. [Google Scholar]
- Exley, R.; Clements, M.A.; Hartung, H.; McIntosh, J.M.; Cragg, S.J. Alpha6-containing nicotinic acetylcholine receptors dominate the nicotine control of dopamine neurotransmission in nucleus accumbens. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2008, 33, 2158–2166. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Moretti, M.; Gaimarri, A.; Zanardi, A.; Clementi, F.; Zoli, M. Heterogeneity and complexity of native brain nicotinic receptors. Biochem. Pharmacol. 2007, 74, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Zoli, M.; Clementi, F. Brain nicotinic acetylcholine receptors: Native subtypes and their relevance. Trends Pharmacol. Sci. 2006, 27, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Garza, A.; Huang, L.Z.; Son, J.-H.; Winzer-Serhan, U.H. Expression of nicotinic acetylcholine receptors and subunit messenger RNAs in the enteric nervous system of the neonatal rat. Neuroscience 2009, 158, 1521–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdanowski, R.; Krzyżowska, M.; Ujazdowska, D.; Lewicka, A.; Lewicki, S. Role of α7 nicotinic receptor in the immune system and intracellular signaling pathways. Cent. Eur. J. Immunol. 2015, 3, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotti, C.; Clementi, F. Neuronal nicotinic receptors: From structure to pathology. Prog. Neurobiol. 2004, 74, 363–396. [Google Scholar] [CrossRef] [PubMed]
- Steinlein, O.K.; Bertrand, D. Nicotinic receptor channelopathies and epilepsy. Pflüg. Arch. Eur. J. Physiol. 2010, 460, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Rubboli, F.; Court, J.A.; Sala, C.; Morris, C.; Chini, B.; Perry, E.; Clementi, F. Distribution of Nicotinic Receptors in the Human Hippocampus and Thalamus. Eur. J. Neurosci. 1994, 6, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2017. [Google Scholar] [CrossRef] [PubMed]
- Spence, I.; Gillessen, D.; Gregson, R.P.; Quinn, R.J. Characterization of the neurotoxic constituents of Conus geographus (L) venom. Life Sci. 1977, 21, 1759–1769. [Google Scholar] [CrossRef]
- Olivera, B.M.; Cruz, L.J. Conotoxins, in retrospect. Tox. Off. J. Int. Soc. Toxinol. 2001, 39, 7–14. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.-H.; Alewood, P.F.; Lewis, R.J. Intraspecific variations in Conus geographus defence-evoked venom and estimation of the human lethal dose. Toxicon 2014, 91, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.A.; Jin, A.-H.; Dutertre, S.; Giacomotto, J.; Mohialdeen, H.; Vetter, I.; Alewood, P.F.; Lewis, R.J. Comparative Venomics Reveals the Complex Prey Capture Strategy of the Piscivorous Cone Snail Conus catus. J. Proteome Res. 2015, 14, 4372–4381. [Google Scholar] [CrossRef] [PubMed]
- Favreau, P.; Krimm, I.; Le Gall, F.; Bobenrieth, M.J.; Lamthanh, H.; Bouet, F.; Servent, D.; Molgo, J.; Ménez, A.; Letourneux, Y.; Lancelin, J.M. Biochemical characterization and nuclear magnetic resonance structure of novel alpha-conotoxins isolated from the venom of Conus consors. Biochemistry (Moscow) 1999, 38, 6317–6326. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, R.B.; DelaCruz, R.G.; Grose, J.H.; McIntosh, J.M.; Yoshikami, D.; Olivera, B.M. Critical Residues Influence the Affinity and Selectivity of α-Conotoxin MI for Nicotinic Acetylcholine Receptors. Biochemistry (Moscow) 1999, 38, 13310–13315. [Google Scholar] [CrossRef]
- Groebe, D.R.; Gray, W.R.; Abramson, S.N. Determinants Involved in the Affinity of α-Conotoxins GI and SI for the Muscle Subtype of Nicotinic Acetylcholine Receptors †. Biochemistry (Moscow) 1997, 36, 6469–6474. [Google Scholar] [CrossRef] [PubMed]
- Cartier, G.E.; Yoshikami, D.; Gray, W.R.; Luo, S.; Olivera, B.M.; McIntosh, J.M. A New—Conotoxin Which Targets 32 Nicotinic Acetylcholine Receptors. J. Biol. Chem. 1996, 271, 7522–7528. [Google Scholar] [CrossRef] [PubMed]
- Gyanda, R.; Banerjee, J.; Chang, Y.-P.; Phillips, A.M.; Toll, L.; Armishaw, C.J. Oxidative folding and preparation of α-conotoxins for use in high-throughput structure—Activity relationship studies. J. Pept. Sci. 2013, 19, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chew, G.; Hawrot, E.; Chi, C.; Wang, C.; Xu, M. Two Potent α3/5 Conotoxins from Piscivorous Conus achatinus. Acta Biochim. Biophys. Sin. 2007, 39, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, Y.; Sakakibara, S. Primary and secondary structure of conotoxin GI, a neurotoxic tridecapeptide from a marine snail. FEBS Lett. 1982, 148, 260–262. [Google Scholar] [CrossRef] [Green Version]
- Price-Carter, M.; Gray, W.R.; Goldenberg, D.P. Folding of ω-Conotoxins. 1. Efficient Disulfide-Coupled Folding of Mature Sequences in Vitro. Biochemistry (Moscow) 1996, 35, 15537–15546. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Nagasawa, H.; Suzuki, A. 2,2′-Bispyridyl disulfide rapidly induces intramolecular disulfide bonds in peptides. Peptides 1999, 20, 881–884. [Google Scholar] [CrossRef]
- Maslennikov, I.V.; Sobol, A.G.; Gladky, K.V.; Lugovskoy, A.A.; Ostrovsky, A.G.; Tsetlin, V.I.; Ivanov, V.T.; Arseniev, A.S. Two distinct structures of α-conotoxin GI in aqueous solution. Eur. J. Biochem. 1998, 254, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Gouda, H.; Yamazaki, K.; Hasegawa, J.; Kobayashi, Y.; Nishiuchi, Y.; Sakakibara, S.; Hirono, S. Solution structure of alpha-conotoxin MI determined by 1H-NMR spectroscopy and molecular dynamics simulation with the explicit solvent water. Biochim. Biophys. Acta 1997, 1343, 327–334. [Google Scholar] [CrossRef]
- Terlau, H.; Shon, K.J.; Grilley, M.; Stocker, M.; Stühmer, W.; Olivera, B.M. Strategy for rapid immobilization of prey by a fish-hunting marine snail. Nature 1996, 381, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, J.A. Intraspecific variation of venom injected by fish-hunting Conus snails. J. Exp. Biol. 2005, 208, 2873–2883. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Biass, D.; Stöcklin, R.; Favreau, P. Dramatic intraspecimen variations within the injected venom of Conus consors: An unsuspected contribution to venom diversity. Toxicon 2010, 55, 1453–1462. [Google Scholar] [CrossRef] [PubMed]
- Bren, N.; Sine, S.M. Hydrophobic Pairwise Interactions Stabilize α-Conotoxin MI in the Muscle Acetylcholine Receptor Binding Site. J. Biol. Chem. 2000, 275, 12692–12700. [Google Scholar] [CrossRef] [PubMed]
- Pucci, L.; Grazioso, G.; Dallanoce, C.; Rizzi, L.; De Micheli, C.; Clementi, F.; Bertrand, S.; Bertrand, D.; Longhi, R.; De Amici, M.; et al. Engineering of α-conotoxin MII-derived peptides with increased selectivity for native α6β2* nicotinic acetylcholine receptors. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 3775–3789. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Nicke, A.; Lewis, R.J. β2 Subunit Contribution to 4/7 α-Conotoxin Binding to the Nicotinic Acetylcholine Receptor. J. Biol. Chem. 2005, 280, 30460–30468. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.B.S.; Baker, M.R.; Kim, D.H.; LeRoy, M.; Toribo, P.; Bingham, J.-P. Cone snail milked venom dynamics—A quantitative study of Conus purpurascens. Toxicon 2012, 60, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Kohn, A.J. PISCIVOROUS GASTROPODS OF THE GENUS CONUS*. Proc. Natl. Acad. Sci. USA 1956, 42, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Kelley, W.P.; Schulz, J.R.; Jakubowski, J.A.; Gilly, W.F.; Sweedler, J.V. Two Toxins from Conus striatus that Individually Induce Tetanic Paralysis. Biochemistry (Moscow) 2006, 45, 14212–14222. [Google Scholar] [CrossRef] [PubMed]
- Violette, A.; Biass, D.; Dutertre, S.; Koua, D.; Piquemal, D.; Pierrat, F.; Stöcklin, R.; Favreau, P. Large-scale discovery of conopeptides and conoproteins in the injectable venom of a fish-hunting cone snail using a combined proteomic and transcriptomic approach. J. Proteom. 2012, 75, 5215–5225. [Google Scholar] [CrossRef] [PubMed]
- Güntert, P. Automated NMR structure calculation with CYANA. Methods Mol. Biol. 2004, 278, 353–378. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CIA | CIB | |
|---|---|---|
| Experimental restraints | ||
| Interproton distance restraints | 69 | 98 |
| Intraresidue | 30 | 30 |
| Sequential | 28 | 47 |
| Medium range (i-j < 5) | 11 | 18 |
| Long range (i-j ≥5) | 0 | 3 |
| Disulfide-bond restraints | 4 | 4 |
| Dihedral-angle restraints | 21 | 22 |
| R.m.s deviations from mean coordinate structure (Å) | ||
| Backbone atoms | 0.95 ± 0.33 | 0.48 ± 0.16 |
| Backbone atoms (res 5–11) | 0.08 ± 0.04 | 0.19 ± 0.10 |
| All heavy atoms | 1.92 ± 0.46 | 0.95 ± 0.22 |
| All heavy atoms (res 5–11) | 0.39 ± 0.41 | 0.80 ± 0.19 |
| Ramachandran Statistics | ||
| Clashscore, all atoms | 0 ± 0 | 0 ± 0 |
| % in most favoured region | 85.7 ± 0 | 91 ± 10 |
| MolProbity score | 2.13 ± 0.14 | 1.68 ± 0.45 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giribaldi, J.; Wilson, D.; Nicke, A.; El Hamdaoui, Y.; Laconde, G.; Faucherre, A.; Moha Ou Maati, H.; Daly, N.L.; Enjalbal, C.; Dutertre, S. Synthesis, Structure and Biological Activity of CIA and CIB, Two α-Conotoxins from the Predation-Evoked Venom of Conus catus. Toxins 2018, 10, 222. https://doi.org/10.3390/toxins10060222
Giribaldi J, Wilson D, Nicke A, El Hamdaoui Y, Laconde G, Faucherre A, Moha Ou Maati H, Daly NL, Enjalbal C, Dutertre S. Synthesis, Structure and Biological Activity of CIA and CIB, Two α-Conotoxins from the Predation-Evoked Venom of Conus catus. Toxins. 2018; 10(6):222. https://doi.org/10.3390/toxins10060222
Chicago/Turabian StyleGiribaldi, Julien, David Wilson, Annette Nicke, Yamina El Hamdaoui, Guillaume Laconde, Adèle Faucherre, Hamid Moha Ou Maati, Norelle L. Daly, Christine Enjalbal, and Sébastien Dutertre. 2018. "Synthesis, Structure and Biological Activity of CIA and CIB, Two α-Conotoxins from the Predation-Evoked Venom of Conus catus" Toxins 10, no. 6: 222. https://doi.org/10.3390/toxins10060222