Differential Toxicity of Cyanobacteria Isolated from Marine Sponges towards Echinoderms and Crustaceans



Abstract
:1. Introduction
2. Results
2.1. Acute Toxicity Assay Using Nauplii of Artemia Salina
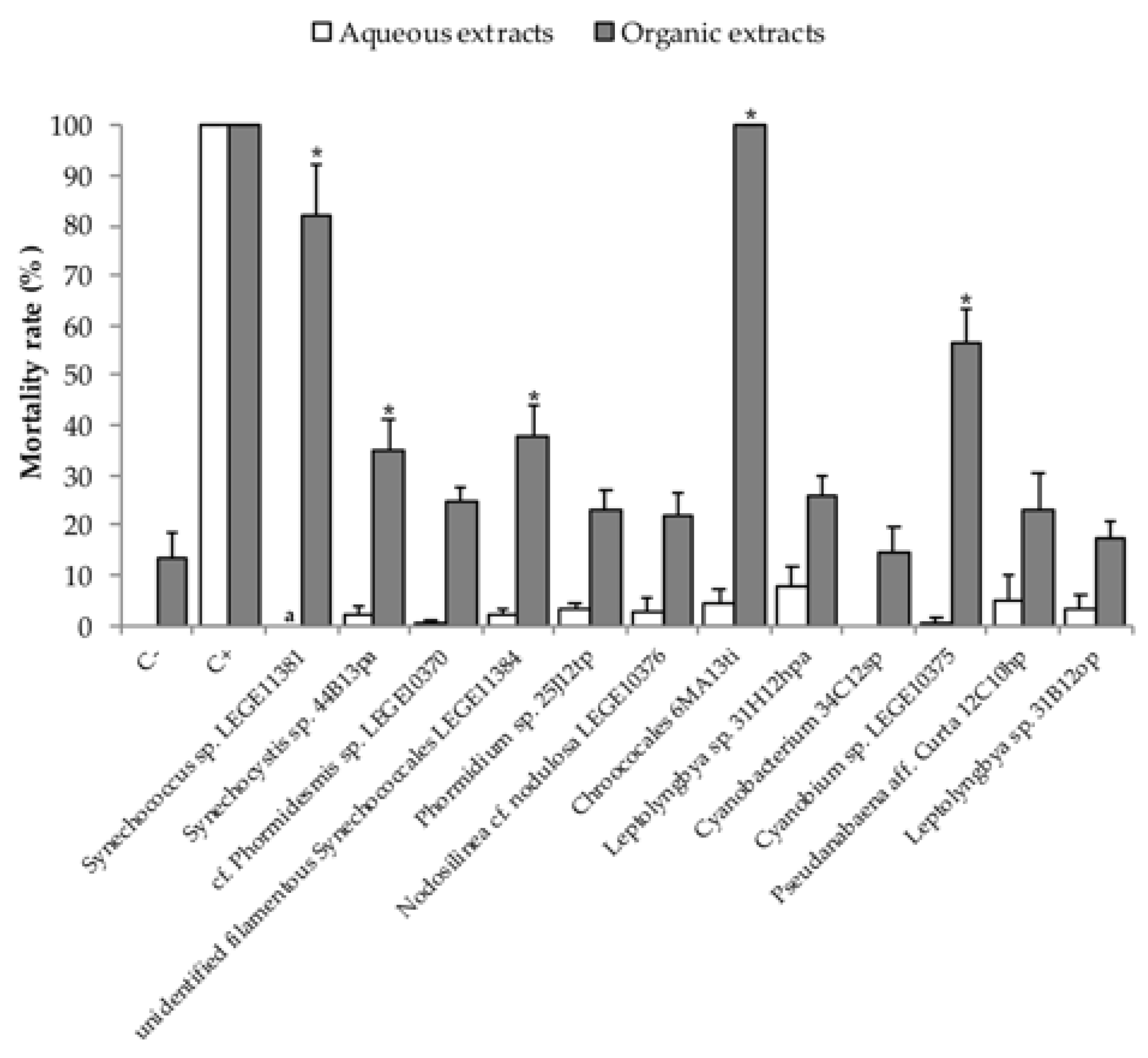
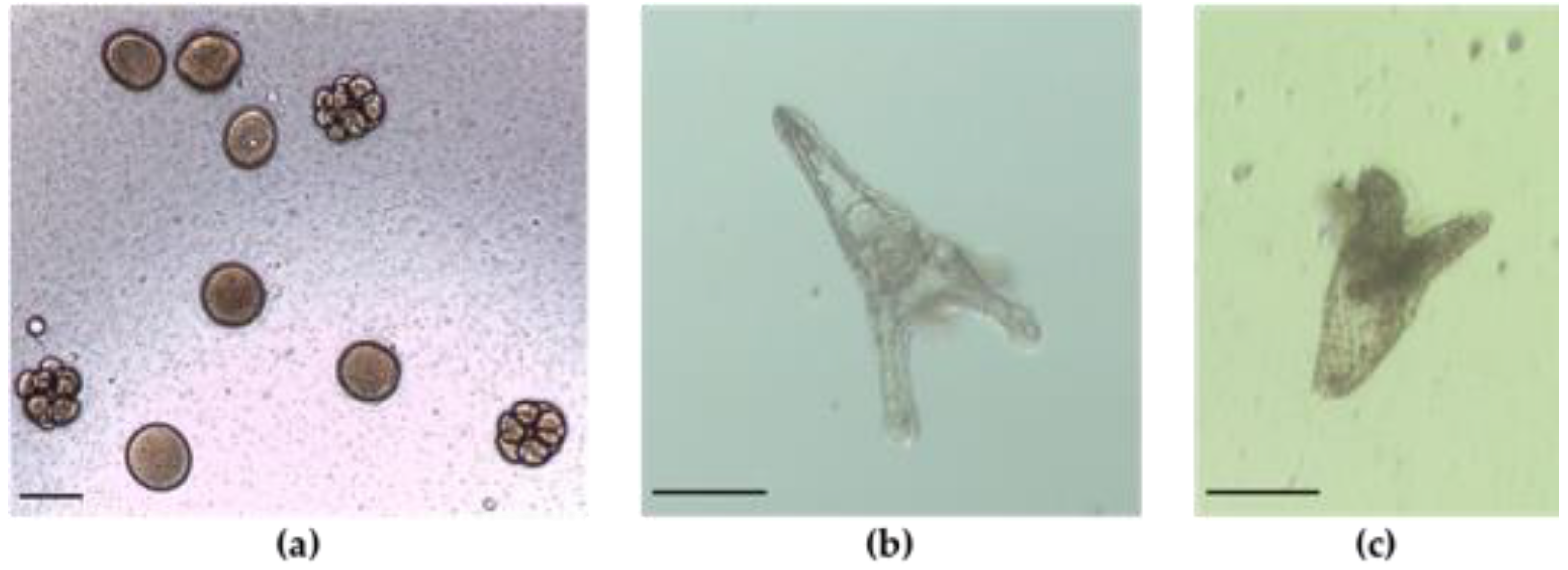
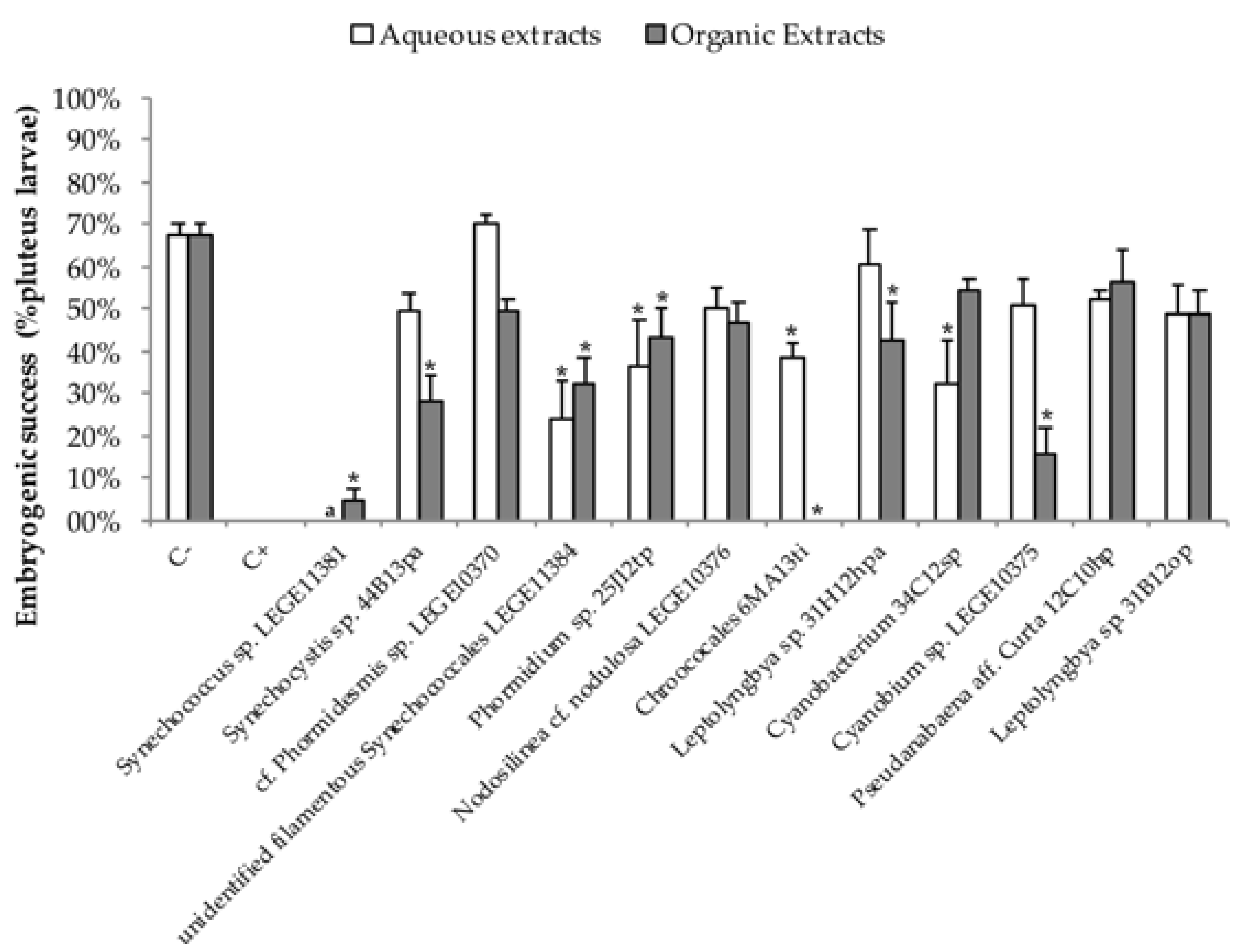
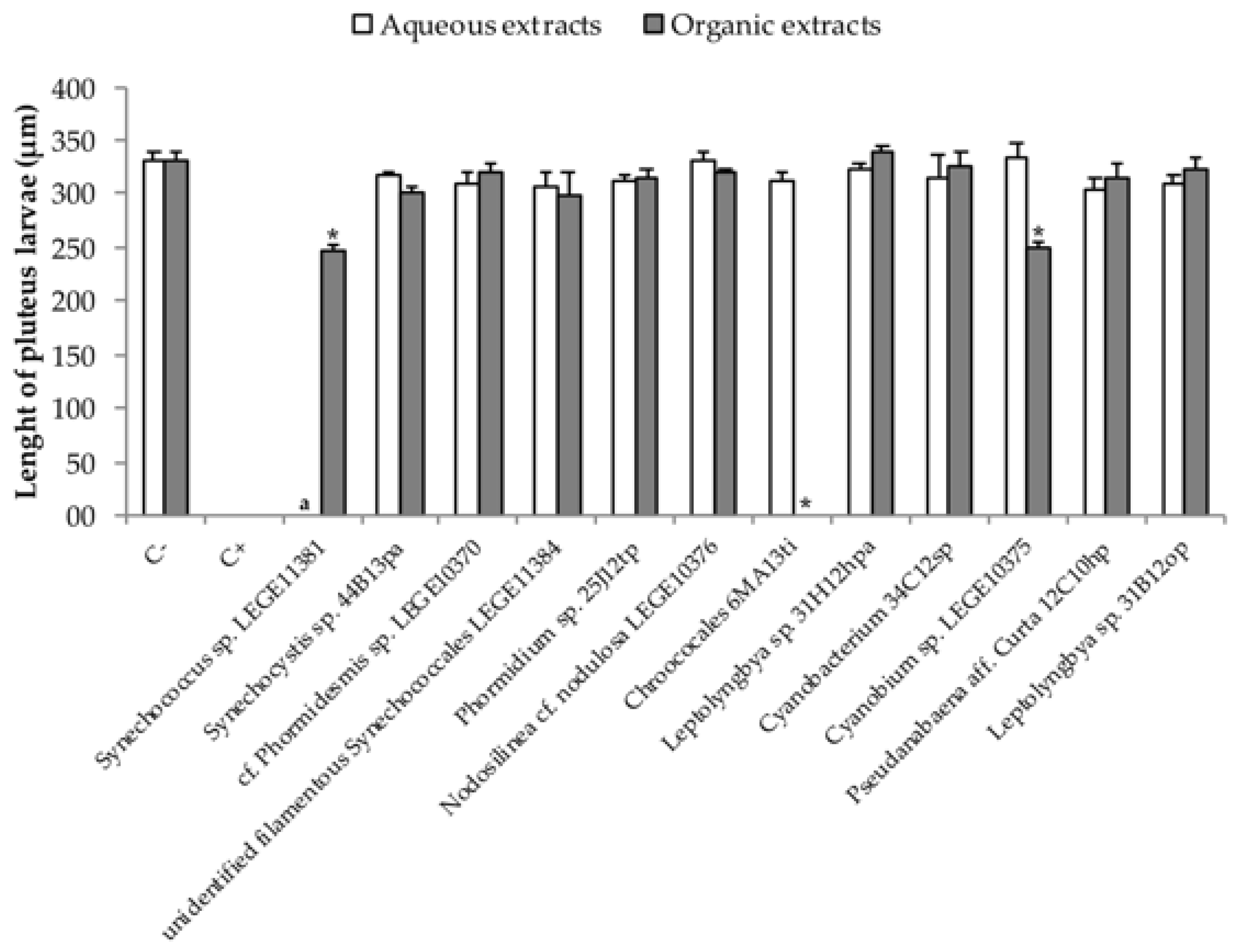
2.2. Embryo—Larval Acute Toxicity Assay with Paracentrotus Lividus
2.3. Hemolytic Assay
3. Discussion
4. Materials and Methods
4.1. Cyanobacterial Strains Selection and Biomass Production
4.2. Preparation of Cyanobacterial Extracts
4.3. Bioassays
4.3.1. Acute Toxiciyy Assay Using Nauplii of Artemia Salina
4.3.2. Embryo-Larval Acute Toxicity Assay with Paracentrotus Lividus
4.3.3. Hemolytic Assay
4.3.4. Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, D.G.; Duggan, P.S. Tansley review no. 107. Heterocyst and akinete differentiation in cyanobacteria. New Phytol. 1999, 144, 3–33. [Google Scholar] [CrossRef]
- Frazão, B.; Martins, R.; Vasconcelos, V. Are known cyanotoxins involved in the toxicity of picoplanktonic and filamentous north atlantic marine cyanobacteria? Mar. Drugs 2010, 8, 1908–1919. [Google Scholar] [CrossRef] [PubMed]
- Mi, Y.; Zhang, J.; He, S.; Yan, X. New peptides isolated from marine cyanobacteria, an overview over the past decade. Mar. Drugs 2017, 15, 132. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Brito, Â.; Ramos, V.; Mota, R.; Lima, S.; Santos, A.; Vieira, J.; Vieira, C.P.; Kaštovský, J.; Vasconcelos, V.M.; Tamagnini, P. Description of new genera and species of marine cyanobacteria from the portuguese atlantic coast. Mol. Phylogenet. Evol. 2017, 111, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Brito, Â.; Ramos, V.; Seabra, R.; Santos, A.; Santos, C.L.; Lopo, M.; Ferreira, S.; Martins, A.; Mota, R.; Frazao, B.; et al. Culture-dependent characterization of cyanobacterial diversity in the intertidal zones of the portuguese coast: A polyphasic study. Syst. Appl. Microbiol. 2012, 35, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Afonso, T.B.; Costa, M.S.; Rezende de Castro, R.; Freitas, S.; Silva, A.; Schneider, M.P.C.; Martins, R.; Leão, P.N. Bartolosides e–k from a marine coccoid cyanobacterium. J. Nat. Prod. 2016, 79, 2504–2513. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Garcia, M.; Costa-Rodrigues, J.; Costa, M.S.; Ribeiro, M.J.; Fernandes, M.H.; Barros, P.; Barreiro, A.; Vasconcelos, V.; Martins, R. Exploring bioactive properties of marine cyanobacteria isolated from the portuguese coast: High potential as a source of anticancer compounds. Mar. Drugs 2014, 12, 98–114. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.S.; Costa, M.; Ramos, V.; Leao, P.N.; Barreiro, A.; Vasconcelos, V.; Martins, R. Picocyanobacteria from a clade of marine Cyanobium revealed bioactive potential against microalgae, bacteria, and marine invertebrates. J. Toxicol. Environ. Health Part A 2015, 78, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Leão, P.N.; Ramos, V.; Gonçalves, P.B.; Viana, F.; Lage, O.M.; Gerwick, W.H.; Vasconcelos, V.M. Chemoecological screening reveals high bioactivity in diverse culturable portuguese marine cyanobacteria. Mar. Drugs 2013, 11, 1316–1335. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.; Fernandez, N.; Beiras, R.; Vasconcelos, V. Toxicity assessment of crude and partially purified extracts of marine Synechocystis and Synechococcus cyanobacterial strains in marine invertebrates. Toxicon 2007, 50, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.; Pereira, P.; Welker, M.; Fastner, J.; Vasconcelos, V.M. Toxicity of culturable cyanobacteria strains isolated from the portuguese coast. Toxicon 2005, 46, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.F.; Ramos, M.F.; Herfindal, L.; Sousa, J.A.; Skaerven, K.; Vasconcelos, V.M. Antimicrobial and cytotoxic assessment of marine cyanobacteria—Synechocystis and Synechococcus. Mar. Drugs 2008, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Brito, Â.; Gaifem, J.; Ramos, V.; Glukhov, E.; Dorrestein, P.C.; Gerwick, W.H.; Vasconcelos, V.M.; Mendes, M.V.; Tamagnini, P. Bioprospecting portuguese atlantic coast cyanobacteria for bioactive secondary metabolites reveals untapped chemodiversity. Algal Res. 2015, 9, 218–226. [Google Scholar] [CrossRef]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemloh, M.L.; Fromont, J.; Brümmer, F.; Usher, K.M. Diversity and abundance of photosynthetic sponges in temperate western australia. BMC Ecol. 2009, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Rützler, K. Associations between caribbean sponges and photosynthetic organisms. In New Perspectives in Sponge Biology; Rutzler, K., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1990; pp. 455–466. [Google Scholar]
- Regueiras, A.; Alex, A.; Pereira, S.; Costa, M.S.; Antunes, A.; Vasconcelos, V. Cyanobacterial diversity in the marine sponge Hymeniacidon perlevis from a temperate region (portuguese coast, northeast atlantic). Aquat. Microb. Ecol. 2017, 79, 259–272. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2010, 27, 165–237. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Rusch, D.; DeMaere, M.Z.; Yung, P.Y.; Lewis, M.; Halpern, A.; Heidelberg, K.B.; Egan, S.; Steinberg, P.D.; Kjelleberg, S. Functional genomic signatures of sponge bacteria reveal unique and shared features of symbiosis. ISME J. 2010, 4, 1557–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konstantinou, D.; Gerovasileiou, V.; Voultsiadou, E.; Gkelis, S. Sponges-cyanobacteria associations: Global diversity overview and new data from the eastern mediterranean. PLoS ONE 2018, 13, e0195001. [Google Scholar] [CrossRef] [PubMed]
- Alex, A.; Antunes, A. Pyrosequencing characterization of the microbiota from atlantic intertidal marine sponges reveals high microbial diversity and the lack of co-occurrence patterns. PLoS ONE 2015, 10, e0127455. [Google Scholar] [CrossRef] [PubMed]
- Alex, A.; Vasconcelos, V.; Tamagnini, P.; Santos, A.; Antunes, A. Unusual symbiotic cyanobacteria association in the genetically diverse intertidal marine sponge Hymeniacidon perlevis (demospongiae, halichondrida). PLoS ONE 2012, 7, e51834. [Google Scholar] [CrossRef] [PubMed]
- Usher, K.M.; Kuo, J.; Fromont, J.; Sutton, D.C. Vertical transmission of cyanobacterial symbionts in the marine sponge Chondrilla australiensis (demospongiae). Hydrobiologia 2001, 461, 9–13. [Google Scholar] [CrossRef]
- Burgsdorf, I.; Slaby, B.M.; Handley, K.M.; Haber, M.; Blom, J.; Marshall, C.W.; Gilbert, J.A.; Hentschel, U.; Steindler, L. Lifestyle evolution in cyanobacterial symbionts of sponges. mBio 2015, 6, e00391–e00415. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.M.; Wang, Y.; Tian, R.M.; Wong, Y.H.; Batang, Z.B.; Al-Suwailem, A.M.; Bajic, V.B.; Qian, P.Y. Symbiotic adaptation drives genome streamlining of the cyanobacterial sponge symbiont “Candidatus synechococcus spongiarum”. mBio 2014, 5, e00079–e00114. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides b and c, cancer cell toxins from a papua new guinea collection of the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Lyngbyastatin 2 and norlyngbyastatin 2, analogues of dolastatin g and nordolastatin g from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 1999, 62, 1702–1706. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L. Isolation, structure determination, and biological activity of lyngbyabellin a from the marine cyanobacterium lyngbya majuscula. J. Nat. Prod. 2000, 63, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Mundt, S.; Kreitlow, S.; Nowotny, A.; Effmert, U. Biochemical and pharmacological investigations of selected cyanobacteria. Int. J. Hyg. Environ. Health 2001, 203, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Papendorf, O.; König, G.M.; Wright, A.D. Hierridin b and 2,4-dimethoxy-6-heptadecyl-phenol, secondary metabolites from the cyanobacterium phormidium ectocarpi with antiplasmodial activity. Phytochemistry 1998, 49, 2383–2386. [Google Scholar] [CrossRef]
- Solis, P.N.; Wright, C.W.; Anderson, M.M.; Gupta, M.P.; Phillipson, J.D. A microwell cytotoxicity assay using Artemia salina. Planta Med. 1993, 59, 250–252. [Google Scholar] [CrossRef] [PubMed]
- Lopes, V.R.; Fernández, N.; Martins, R.F.; Vasconcelos, V. Primary screening of the bioactivity of brackishwater cyanobacteria: Toxicity of crude extracts to Artemia salina larvae and Paracentrotus lividus embryos. Mar. Drugs 2010, 8, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Engström-Öst, J.; Lehtiniemi, M.; Green, S.; Kozlowsky-Suzuki, B.; Viitasalo, M. Does cyanobacterial toxin accumulate in mysid shrimps and fish via copepods? J. Exp. Mar. Biol. Ecol. 2002, 276, 95–107. [Google Scholar] [CrossRef]
- Ferrão-Filho, A.d.S.; Kozlowsky-Suzuki, B.; Azevedo, S.M.F.O. Accumulation of microcystins by a tropical zooplankton community. Aquat. Toxicol. 2002, 59, 201–208. [Google Scholar] [CrossRef]
- Ridley, C.P.; Bergquist, P.R.; Harper, M.K.; Faulkner, D.J.; Hooper, J.N.A.; Haygood, M.G. Speciation and biosynthetic variation in four dictyoceratid sponges and their cyanobacterial symbiont, Oscillatoria spongeliae. Chem. Biol. 2005, 12, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.S.; Carvalho, F.D.; Guilhermino, L.M.; Van Stappen, G. Use of the genus Artemia in ecotoxicity testing. Environ. Pollut. 2006, 144, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Sakiyama, T.; Ueno, H.; Homma, H.; Numata, O.; Kuwabara, T. Purification and characterization of a hemolysin-like protein, sll1951, a nontoxic member of the rtx protein family from the cyanobacterium Synechocystis sp. Strain pcc 6803. J. Bacteriol. 2006, 188, 3535–3542. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-J.; Chien, M.-S.; Wu, F.-J.; Chou, H.-N.; Lee, S.-J. Inhibition of embryonic development by microcystin-lr in zebrafish, Danio rerio. Toxicon 2005, 45, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Pagliara, P.; Caroppo, C. Cytotoxic and antimitotic activities in aqueous extracts of eight cyanobacterial strains isolated from the marine sponge Petrosia ficiformis. Toxicon 2011, 57, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Kótai, J. Instructions for Preparation of Modified Nutrient Solution z8 for Algae; Norwegian Institute for Water Research b-11769: Oslo, Norway, 1972; p. 5. [Google Scholar]
- Rippka, R. Isolation and purification of cyanobacteria. Meth. Enzymol. 1988, 167, 3–27. [Google Scholar] [PubMed]
- Komárek, J. Süßwasserflora von Mitteleuropa, bd. 19/1: Cyanoprokaryota: Chroococcales; Springer Spektrum: Berlin/Heidelberg, Germany, 2008; p. 548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Süßwasserflora von Mitteleuropa, bd. 19/2: Cyanoprokaryota: Oscillatoriales; Elsevier/Spektrum: Berlin/Heidelberg, Germany, 2005; p. 759. [Google Scholar]
- Komárek, J. Süßwasserflora von Mitteleuropa, bd. 19/3: Cyanoprokaryota: Heterocytous Genera; Springer Spektrum: Berlin/Heidelberg, Germany, 2013; p. 1131. [Google Scholar]
- Castenholz, R.W.; Wilmotte, A.; Herdman, M.; Rippka, R.; Waterbury, J.B.; Iteman, I.; Hoffmann, L. Phylum bx. Cyanobacteria. In Bergey’s Manual® of Systematic Bacteriology: Volume One: The Archaea and the Deeply Branching and Phototrophic Bacteria; Boone, D.R., Castenholz, R.W., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2001; pp. 473–599. [Google Scholar]
- Komárek, J.; Kastovský, J.; Mares, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Fernández, N.; Beiras, R. Combined toxicity of dissolved mercury with copper, lead and cadmium on embryogenesis and early larval growth of the Paracentrotus lividus sea-urchin. Ecotoxicology 2001, 10, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.; Malpezzi, E.L.A.; Susini, S.M.M.; De Freitas, J. Hemolytic activity in extracts of the diatom Nitzschia. Toxicon 1997, 35, 305–309. [Google Scholar] [CrossRef]
- Slowing, I.I.; Wu, C.W.; Vivero-Escoto, J.L.; Lin, V.S.Y. Mesoporous silica nanoparticles for reducing hemolytic activity towards mammalian red blood cells. Small 2009, 5, 57–62. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cyanobacterial Strain | Sponge Species | Collection Site |
|---|---|---|
| Synechococcus sp. LEGE11381 | Polymastia sp. | Memória |
| Synechocystis sp. 44B13pa | Polymastia agglutinans | São Roque, Azores |
| cf. Phormidesmis sp. LEGE10370 | Hymeniacidon perlevis | Memória |
| Unidentified filamentous Synechococcales LEGE11384 | Phorbas plumosus | Memória |
| Phormidium sp. 25J12tp | Tedania pilarriosae | Memória |
| Nodosilinea cf. nodulosa LEGE10376 | Hymeniacidon perlevis | Porto Côvo |
| Chroococcales 6MA13ti | Tedania ignis | São Roque, Azores |
| Leptolyngbya sp. 31H12hpa | Halichondria panicea | Memória |
| Cyanobacterium 34C12sp | Unidentified sponge | Caniçal, Madeira |
| Cyanobium sp. LEGE10375 | Hymeniacidon perlevis | Memória |
| Pseudanabaena aff. curta 12C10hp | Hymeniacidon perlevis | Memória |
| Leptolyngbya sp. 31B12op | Ophlitaspongia papila | Memória |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regueiras, A.; Pereira, S.; Costa, M.S.; Vasconcelos, V. Differential Toxicity of Cyanobacteria Isolated from Marine Sponges towards Echinoderms and Crustaceans. Toxins 2018, 10, 297. https://doi.org/10.3390/toxins10070297
Regueiras A, Pereira S, Costa MS, Vasconcelos V. Differential Toxicity of Cyanobacteria Isolated from Marine Sponges towards Echinoderms and Crustaceans. Toxins. 2018; 10(7):297. https://doi.org/10.3390/toxins10070297
Chicago/Turabian StyleRegueiras, Ana, Sandra Pereira, Maria Sofia Costa, and Vitor Vasconcelos. 2018. "Differential Toxicity of Cyanobacteria Isolated from Marine Sponges towards Echinoderms and Crustaceans" Toxins 10, no. 7: 297. https://doi.org/10.3390/toxins10070297
APA StyleRegueiras, A., Pereira, S., Costa, M. S., & Vasconcelos, V. (2018). Differential Toxicity of Cyanobacteria Isolated from Marine Sponges towards Echinoderms and Crustaceans. Toxins, 10(7), 297. https://doi.org/10.3390/toxins10070297