Structural Diversity, Characterization and Toxicology of Microcystins


 , , and
, , and
Abstract
1. Introduction
2. Nomenclature and General Chemical Structure of Microcystins
3. Biosynthesis of Microcystins
4. Structural Elucidation of Microcystins
4.1. Mass Spectrometry for Structural Elucidation
4.2. Preparative Isolation of Microsystins
4.3. Amino Acid Analysis
4.4. Structural Elucidation by Nuclear Magnetic Resonance Spectroscopy
4.5. 3-Dimensional Structures of Microcystins
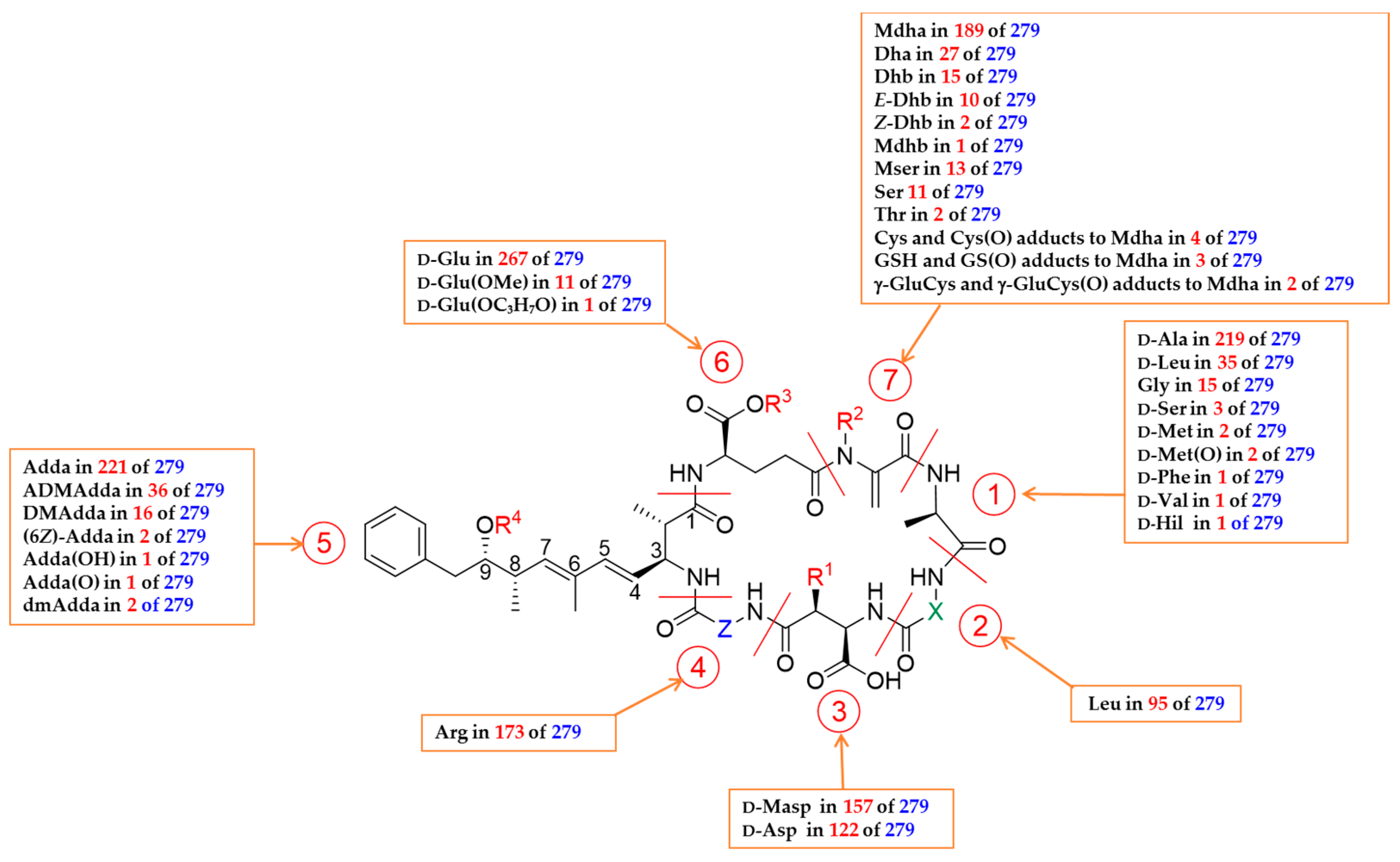
5. Diversity of Characterized Microcystin Congeners
5.1. Congeners with Variable Amino Acid at Position 1
5.2. Congeners with Variable Amino Acid at Positions 2 and 4
5.3. Congeners with Variable Amino Acid at Position 3
5.4. Congeners with Variable Amino Acid at Position 5
5.5. Congeners With Variable Amino Acid at Position 6
5.6. Congeners With Variable Amino Acid at Position 7
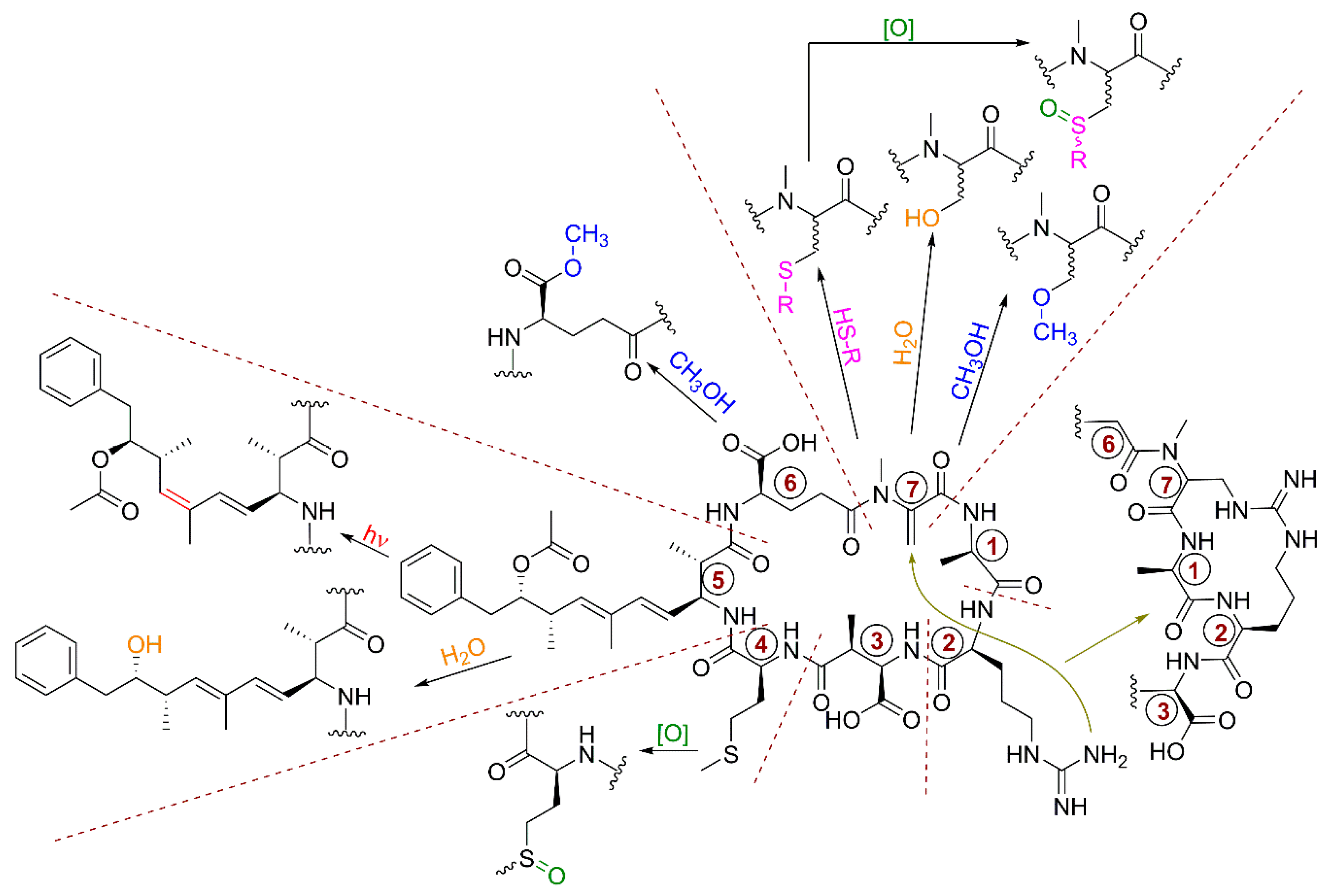
5.7. Chemical and Biochemical Transformations of Microcystins
6. Toxicity of Microcystin Congeners
6.1. Toxicity In Vivo
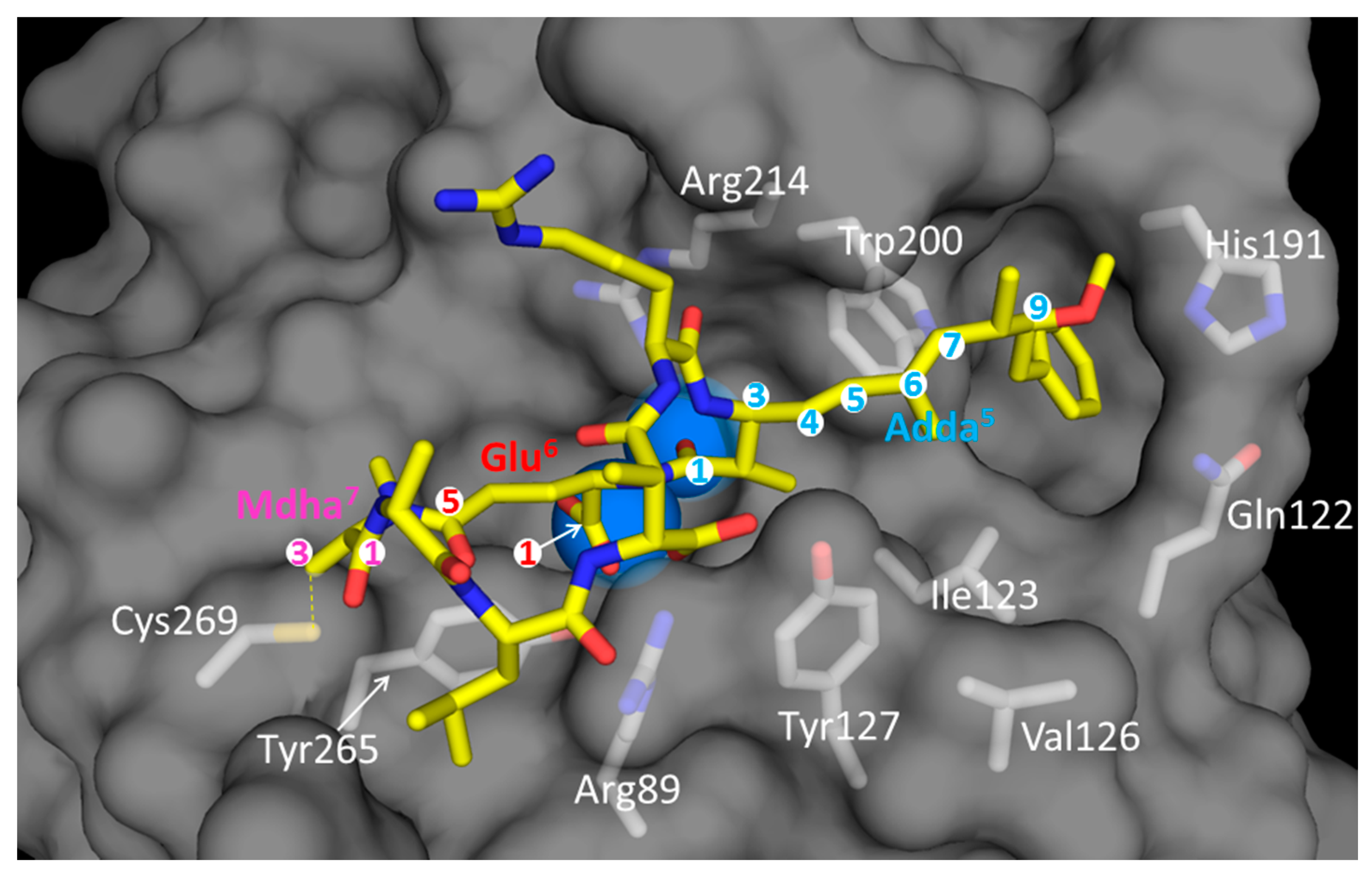
6.2. In Vitro Toxicity: Inhibition of Serine/Threonine PPs
7. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Aha | aminoheptanoic acid |
| Abu(Aba) | 2-aminobutyric acid (L-2-aminobutanoic acid |
| Apa | aminopropionic acid |
| Aib | aminoisobutyric acid (aminoisobutyrate) |
| Bu | 2-aminobutyric acid (L-butyrine) |
| Cit | citrulline |
| ADMAdda | O-acetyl-9-O-desmethylAdda |
| Dha | dehydroalanine |
| Dhb | dehydrobutyrine |
| DMAdda | 9-O-desmethylAdda |
| dmAdda | Adda demethylated at C-2, C-6, or C-8 |
| E(OMe) | glutamic acid methylester |
| H(2)Y | dihydrotyrosine |
| (H4)Y | 4,5,6,7-tetrahydrotyrosine |
| Har | homoarginine |
| Hil | homoisoleucine |
| Hph | homophenylalanine |
| Hty | homotyrosine |
| Kyn | kynurenine |
| Mdhb | N-methyldehydrobutyrine |
| M(O) | methionine-S-oxide |
| M(O2) | methionine S-sulfone |
| Mser | N-methylserine |
| Nfk | N-formylkynurenine |
| Oia | oxindolyalanine |
| Y(OMe) | methoxytyrosine |
References
- Mooij, W.M.; Hülsmann, S.; De Senerpont Domis, L.N.; Nolet, B.A.; Bodelier, P.L.E.; Boers, P.C.M.; Pires, L.M.D.; Gons, H.J.; Ibelings, B.W.; Noordhuis, R.; et al. The impact of climate change on lakes in the Netherlands: A review. Aquat. Ecol. 2005, 39, 381–400. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef]
- Botana, L.M. Toxicological perspective on climate change: Aquatic toxins. Chem. Res. Toxicol. 2016, 29, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Visser, P.M.; Verspagen, J.M.H.; Sandrini, G.; Stal, L.J.; Matthijs, H.C.P.; Davis, T.W.; Paerl, H.W.; Huisman, J. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae 2016, 54, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Russell, C. Food crops irrigated with cyanobacteria-contaminated water: An emerging public health issue in Canada. Environ. Health Rev. 2017, 60, 58–63. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; Chorus, I., Bartram, J., Eds.; E&FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Corbel, S.; Mougin, C.; Bouaïcha, N. Cyanobacterial toxins: Modes of actions, fate in aquatic and soil ecosystems, phytotoxicity and bioaccumulation in agricultural crops. Chemosphere 2014, 96, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and Analysis of Occurrence, Exposure and Toxicity of Cyanobacteria Toxins in Food; EFSA Supporting Publication EN-998; European Food Safety Authority: Parma, Italy, 2016; p. 309. [Google Scholar]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiolocal investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- Pham, T.L.; Utsumi, M. An overview of the accumulation of microcystins in aquatic ecosystems. J. Environ. Manag. 2018, 213, 520–529. [Google Scholar] [CrossRef]
- Ettoumi, A.; El Khalloufi, F.; El Ghazali, I.; Oudra, B.; Amrani, A.; Nasri, H.; Bouaïcha, N. Bioaccumulation of cyanobacterial toxins in aquatic organisms and its consequences for public health. In Zooplankton and Phytoplankton; Kattel, G., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2011; pp. 1–34. [Google Scholar]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef]
- Louw, P.G.J.; Smit, J.D. The active constituent of the poisonous algae, Microcystis toxica Stephens. With a note on experimental cases of algae poisoning in small animals. S. Afr. Ind. Chem. 1950, 4, 62–66. [Google Scholar]
- Bishop, C.T.; Anet, E.F.L.J.; Gorham, P.R. Isolation and identification of the fast death factor in Microcystis aeruginosa NRC-1. Can. J. Biochem. Physiol. 1959, 37, 453–471. [Google Scholar] [CrossRef] [PubMed]
- Botes, D.P.; Tuinman, A.A.; Wessels, P.L.; Viljoen, C.C.; Kruger, H.; Williams, D.H.; Santikarn, S.; Smith, R.J.; Hammond, S.J. The structure of cyanoginosin-LA, a cyclic heptapeptide toxin from the cyanobacterium Microcystis aeruginosa. J. Chem. Soc. Perkin Trans. 1 1984, 2311–2318. [Google Scholar] [CrossRef]
- Botes, D.P.; Wessels, P.L.; Kruger, H.; Runnegar, M.T.C.; Santikarn, S.; Smith, R.J.; Barna, J.C.J.; Williams, D.H. Structural studies on cyanoginosins-LR, -YR, -YA, and -YM, peptide toxins from Microcystis aeruginosa. J. Chem. Soc. Perkin Trans. 1 1985, 2747. [Google Scholar] [CrossRef]
- Botes, D.P.; Kruger, H.; Viljoen, C.C. Isolation and characterization of four toxins from the blue-green alga, Microcystis aeruginosa. Toxicon 1982, 20, 945–954. [Google Scholar] [CrossRef]
- Botes, D.P.; Vilioen, C.C.; Kruger, H.; Wessels, P.L.; Williams, D.H. Configuration assignments of the amino acid residues and the presence of N-methyldehydroalanine in toxins of the blue-green alga Microcystis aeruginosa. Toxicon 1982, 20, 1037–1042. [Google Scholar] [CrossRef]
- Botes, D.P. Cyanoginosins-isolation and structure. In Mycotoxins and Phycotoxins, Bioactive Molecules; Steyn, P.S., Vleggaar, R., Eds.; Elsevier: Amsterdam, The Netherlands, 1986; Volume 1, p. 167. [Google Scholar]
- Santikarn, S.; Williams, D.H.; Smith, R.J.; Hammond, S.J.; Botes, D.P.; Tuinman, A.; Wessels, P.L.; Viljoen, C.C.; Kruger, H. A partial structure for the toxin BE-4 from the blue-green algae, Microcystis aeruginosa. J. Chem. Soc. Chem. Commun. 1983, 12, 652. [Google Scholar] [CrossRef]
- Nishizawa, T.; Asayama, M.; Fujii, K.; Harada, K.-I.; Shirai, M. Genetic analysis of the peptide synthetase genes for a cyclic heptapeptide microcystin in Microcystis spp. J. Biochem. 1999, 126, 520–529. [Google Scholar] [CrossRef]
- Nishizawa, T.; Ueda, A.; Asayama, M.; Fujii, K.; Harada, K.-I.; Ochi, K.; Shirai, M. Polyketide synthase gene coupled to the peptide synthetase module involved in the biosynthesis of the cyclic heptapeptide microcystin. J. Biochem. 2000, 127, 779–789. [Google Scholar] [CrossRef]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide–polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Dittmann, E.; Börner, T. Genetic contributions to the risk assessment of microcystin in the environment. Toxicol. Appl. Pharmacol. 2005, 203, 192–200. [Google Scholar] [CrossRef]
- Song, L.; Sano, T.; Li, R.; Watanabe, M.M.; Liu, Y.; Kaya, K. Microcystin production of Microcystis viridis (cyanobacteria) under different culture conditions. Physiol. Res. 1998, 46, 19–23. [Google Scholar] [CrossRef]
- Kameyama, K.; Sugiura, N.; Inamori, Y.; Maekawa, T. Characteristics of microcystin production in the cell cycle of Microcystis viridis. Environ. Toxicol. 2004, 19, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Amé, M.V.; Wunderlin, D.A. Effects of iron, ammonium and temperature on microcystin content by a natural concentrated Microcystis aeruginosa population. Water Air Soil Pollut. 2005, 168, 235–248. [Google Scholar] [CrossRef]
- Tonk, L.; Visser, P.M.; Christiansen, G.; Dittmann, E.; Snelder, E.O.F.M.; Wiedner, C.; Mur, L.R.; Huisman, J. The microcystin composition of the cyanobacterium Planktothrix agardhii changes toward a more toxic variant with increasing light intensity. Appl. Environ. Microbiol. 2005, 71, 5177–5181. [Google Scholar] [CrossRef]
- Tonk, L.; Van De Waal, D.B.; Slot, P.; Huisman, J.; Matthijs, H.C.P.; Visser, P.M. Amino acid availability determines the ratio of microcystin variants in the cyanobacterium Planktothrix agardhii: Amino acid availability determines microcystin variants. FEMS Microbiol. Ecol. 2008, 65, 383–390. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Verspagen, J.M.H.; Lürling, M.; Van Donk, E.; Visser, P.M.; Huisman, J. The ecological stoichiometry of toxins produced by harmful cyanobacteria: An experimental test of the carbon-nutrient balance hypothesis: Ecological stoichiometry of toxin production. Ecol. Lett. 2009, 12, 1326–1335. [Google Scholar] [CrossRef]
- Dziallas, C.; Grossart, H.-P. Increasing oxygen radicals and water temperature select for toxic Microcystis sp. PLoS ONE 2011, 6, e25569. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Cary, S.C.; Hamilton, D.P. Modulation of microcystin congener abundance following nitrogen depletion of a Microcystis batch culture. Aquat. Ecol. 2016, 50, 235–246. [Google Scholar] [CrossRef]
- Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Laakso, K.; Wang, H.; Sivonen, K. Recurrent adenylation domain replacement in the microcystin synthetase gene cluster. BMC Evol. Biol. 2007, 7, 183. [Google Scholar] [CrossRef]
- Fewer, D.P.; Tooming-Klunderud, A.; Jokela, J.; Wahlsten, M.; Rouhiainen, L.; Kristensen, T.; Rohrlack, T.; Jakobsen, K.S.; Sivonen, K. Natural occurrence of microcystin synthetase deletion mutants capable of producing microcystins in strains of the genus Anabaena (Cyanobacteria). Microbiology 2008, 154, 1007–1014. [Google Scholar] [CrossRef]
- Fewer, D.P.; Halinen, K.; Sipari, H.; Bernardová, K.; Mänttäri, M.; Eronen, E.; Sivonen, K. Non-autonomous transposable elements associated with inactivation of microcystin gene clusters in strains of the genus Anabaena isolated from the Baltic Sea: Inactivation of mcy genes in Baltic Sea Anabaena. Environ. Microbiol. Rep. 2011, 3, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Christiansen, G.; Fastner, J.; Borner, T. Abundance of active and inactive microcystin genotypes in populations of the toxic cyanobacterium Planktothrix spp. Environ. Microbiol. 2004, 6, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Tooming-Klunderud, A.; Mikalsen, B.; Kristensen, T.; Jakobsen, K.S. The mosaic structure of the mcyABC operon in Microcystis. Microbiology 2008, 154, 1886–1899. [Google Scholar] [CrossRef] [PubMed]
- del Campo, F.F.; Ouahid, Y. Identification of microcystins from three collection strains of Microcystis aeruginosa. Environ. Pollut. 2010, 158, 2906–2914. [Google Scholar] [CrossRef]
- Niedermeyer, T. Microcystin congeners described in the literature. 2013. [Google Scholar] [CrossRef]
- Bouhaddada, R.; Nélieu, S.; Nasri, H.; Delarue, G.; Bouaïcha, N. High diversity of microcystins in a Microcystis bloom from an Algerian lake. Environ. Pollut. 2016, 216, 836–844. [Google Scholar] [CrossRef]
- Miller, T.R.; Beversdorf, L.J.; Weirich, C.A.; Bartlett, S.L. Cyanobacterial toxins of the Laurentian Great Lakes, their toxicological effects, and numerical limits in drinking water. Mar. Drugs 2017, 15, 160. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3: Tables of microcystins and nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 526–537. [Google Scholar]
- Hughes, E.O.; Gorham, P.R.; Zehnder, A. Toxicity of a unialgal culture of Microcystis aeruginosa. Can. J. Microbiol. 1958, 4, 225–236. [Google Scholar] [CrossRef]
- Konst, H.; McKercher, P.D.; Gorham, P.R.; Robertson, A.; Howell, J. Symptoms and pathology produced by Toxic Microcystis aeruginosa NRC-1 in laboratory and domestic animals. Can. J. Comp. Med. Vet. Sci. 1965, 29, 221–228. [Google Scholar]
- Murthy, J.R.; Capindale, J.B. A new isolation and structure for the endotoxin from Microcystis aeruginosa NRC-1. Can. J. Biochem. 1970, 48, 508–510. [Google Scholar] [CrossRef]
- Rabin, P.; Darbre, A. An improved extraction procedure for the endotoxin from Microcystis aeruginosa NRC-1. Biochem. Soc. Trans. 1975, 3, 428–430. [Google Scholar] [CrossRef] [PubMed]
- Painuly, P.; Perez, R.; Fukai, T.; Shimizu, Y. The structure of a cyclic peptide toxin, cyanogenosin-RR from Microcystis aeruginosa. Tetrahedron Lett. 1988, 29, 11–14. [Google Scholar] [CrossRef]
- Kusumi, T.; Ooi, T.; Watanabe, M.M.; Takahashi, H.; Kakisawa, H. Cyanoviridin RR, a toxin from the cyanobacterium (blue-green alga) Microcystis viridis. Tetrahedron Lett. 1987, 28, 4695–4698. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Eschedor, J.T.; Patterson, G.M.L.; Moore, R.E. Toxicity and partial structure of a hepatotoxic peptide produced by the cyanobacterium Nodularia spumigena Mertens emend. L575 from New Zealand. Appl. Environ. Microbiol. 1988, 54, 2257–2263. [Google Scholar] [PubMed]
- Carmichael, W.W.; Beasley, V.; Bunner, D.L.; Eloff, J.N.; Falconer, I.; Gorham, P.; Harada, K.; Krishnamurthy, T.; Min-Juan, Y.; Moore, R.E.; et al. Naming of cyclic heptapeptide toxins of cyanobacteria (blue-green algae). Toxicon 1988, 26, 971–973. [Google Scholar] [CrossRef]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef]
- Dixon, H.B.F.; Cornish-Bowden, A.; Liebecq, C.; Loening, K.L.; Moss, G.P.; Reedijk, J.; Velick, S.F.; Vliegenthart, J.F.G.; Bielka, H.; Sharon, N.; et al. International Union of pure and applied chemistry and international union of biochemistry, join commission on biochemical nomenclature. Nomenclature and symbolism for amino acids and peptides. Pure Appl. Chem. 1984, 56, 595–624. [Google Scholar]
- Miles, C.O.; Stirling, D. Toxin Mass List, Version 16. Available online: https://www.researchgate.net/publication/316605326_Toxin_mass_list_version_16 (accessed on 24 October 2019). [CrossRef]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef]
- Christiansen, G.; Fastner, J.; Erhard, M.; Borner, T.; Dittmann, E. Microcystin biosynthesis in Planktothrix: Genes, evolution, and manipulation. J. Bacteriol. 2003, 185, 564–572. [Google Scholar] [CrossRef]
- Rouhiainen, L.; Vakkilainen, T.; Siemer, B.L.; Buikema, W.; Haselkorn, R.; Sivonen, K. Genes coding for hepatotoxic heptapeptides (microcystins) in the cyanobacterium Anabaena Strain 90. Appl. Environ. Microbiol. 2004, 70, 686–692. [Google Scholar] [CrossRef]
- Dittmann, E.; Neilan, B.A.; Erhard, M.; von Döhren, H.; Börner, T. Insertional mutagenesis of a peptide synthetase gene that is responsible for hepatotoxin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Mol. Microbiol. 1997, 26, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Dittmann, E.; Mazmouz, R.; Ongley, S.E.; D’Agostino, P.M.; Neilan, B.A. The genetics, biosynthesis and regulation of toxic specialized metabolites of cyanobacteria. Harmful Algae 2016, 54, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, G.; Kurmayer, R.; Liu, Q.; Borner, T. Transposons inactivate biosynthesis of the nonribosomal peptide microcystin in naturally occurring Planktothrix spp. Appl. Environ. Microbiol. 2006, 72, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Mikalsen, B.; Boison, G.; Skulberg, O.M.; Fastner, J.; Davies, W.; Gabrielsen, T.M.; Rudi, K.; Jakobsen, K.S. Natural variation in the microcystin synthetase operon mcyABC and impact on microcystin production in Microcystis strains. J. Bacteriol. 2003, 185, 2774–2785. [Google Scholar] [CrossRef] [PubMed]
- Shishido, T.K.; Jokela, J.; Humisto, A.; Suurnäkki, S.; Wahlsten, M.; Alvarenga, D.O.; Sivonen, K.; Fewer, D.P. The biosynthesis of rare homo-amino acid containing variants of microcystin by a benthic cynabacterium. Mar. Drugs 2019, 17, 271. [Google Scholar] [CrossRef]
- Kaebernick, M.; Neilan, B.A.; Borner, T.; Dittmann, E. Light and the transcriptional response of the microcystin biosynthesis gene cluster. Appl. Environ. Microbiol. 2000, 66, 3387–3392. [Google Scholar] [CrossRef]
- Oh, H.-M.; Lee, S.J.; Jang, M.-H.; Yoon, B.-D. Microcystin production by Microcystis aeruginosa in a phosphorus-limited chemostat. Appl. Environ. Microbiol. 2000, 66, 176–179. [Google Scholar] [CrossRef]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Iron availability affects mcyD expression and microcystin-LR synthesis in Microcystis aeruginosa PCC7806: Iron starvation triggers microcystin synthesis. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar] [CrossRef]
- Jähnichen, S.; Long, B.M.; Petzoldt, T. Microcystin production by Microcystis aeruginosa: Direct regulation by multiple environmental factors. Harmful Algae 2011, 12, 95–104. [Google Scholar] [CrossRef]
- Tao, M.; Xie, P.; Chen, J.; Qin, B.; Zhang, D.; Niu, Y.; Zhang, M.; Wang, Q.; Wu, L. Use of a generalized additive model to investigate key abiotic factors affecting microcystin cellular quotas in heavy bloom areas of Lake Taihu. PLoS ONE 2012, 7, e32020. [Google Scholar] [CrossRef]
- Meissner, S.; Fastner, J.; Dittmann, E. Microcystin production revisited: Conjugate formation makes a major contribution: Production of cyanobacterial toxins strongly underestimated. Environ. Microbiol. 2013, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, J.S.M.; Giani, A. Microcystin production and regulation under nutrient stress conditions in toxic Microcystis strains. Appl. Environ. Microbiol. 2014, 80, 5836–5843. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Kong, F. UV-B Exposure affects the biosynthesis of microcystin in toxic Microcystis aeruginosa cells and its degradation in the extracellular space. Toxins 2015, 7, 4238–4252. [Google Scholar] [CrossRef] [PubMed]
- Horst, G.P.; Sarnelle, O.; White, J.D.; Hamilton, S.K.; Kaul, R.B.; Bressie, J.D. Nitrogen availability increases the toxin quota of a harmful cyanobacterium, Microcystis aeruginosa. Water Res. 2014, 54, 188–198. [Google Scholar] [CrossRef]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural product biosynthetic diversity and comparative genomics of the cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Smith, V.H.; Declerck, S.A.J.; Stam, E.C.M.; Elser, J.J. Stoichiometric regulation of phytoplankton toxins. Ecol. Lett. 2014, 17, 736–742. [Google Scholar] [CrossRef]
- Monchamp, M.E.; Pick, F.R.; Beisner, B.E.; Maranger, R. Nitrogen forms influence microcystin concentration and composition via changes in cyanobacterial community structure. PLoS ONE 2014, 9, e85573. [Google Scholar] [CrossRef]
- Walsh, C.T.; Chen, H.; Keating, T.A.; Hubbard, B.K.; Losey, H.C.; Luo, L.; Marshall, C.G.; Miller, D.A.; Patel, H.M. Tailoring enzymes that modify nonribosomal peptides during and after chain elongation on NRPS assembly lines. Curr. Opin. Chem. Biol. 2001, 5, 525–534. [Google Scholar] [CrossRef]
- Shishido, T.; Kaasalainen, U.; Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Fiore, M.; Yunes, J.; Rikkinen, J.; Sivonen, K. Convergent evolution of [D-Leucine1]microcystin-LR in taxonomically disparate cyanobacteria. BMC Evol. Biol. 2013, 13, 1–15. [Google Scholar] [CrossRef]
- Namikoshi, M.; Rinehart, K.L.; Sakai, R.; Stotts, R.R.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W.; Evans, W.R. Identification of 12 hepatotoxins from a Homer lake bloom of the cyanobacteria Microcystis aeruginosa, Microcystis viridis, Microcystis wesenbergii; nine new microcystins. J. Org. Chem. 1992, 57, 866–872. [Google Scholar] [CrossRef]
- Miles, C.O.; Sandvik, M.; Nonga, H.E.; Rundberget, T.; Wilkins, A.L.; Rise, F.; Ballot, A. Identification of microcystins in a Lake Victoria cyanobacterial bloom using LC–MS with thiol derivatization. Toxicon 2013, 70, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Rinehart, K.L.; Sakai, R.; Sivonen, K.; Carmichael, W.W. Structures of three new cyclic heptapeptide hepatotoxins produced by the cyanobacterium (blue-green alga) Nostoc sp. strain 152. J. Org. Chem. 1990, 55, 6135–6139. [Google Scholar] [CrossRef]
- Sivonen, K.; Carmichael, W.W.; Namikoshi, M.; Rinehart, K.L.; Dahlem, A.M. Isolation and characterization of hepatotoxic microcystin homologs from the filamentous freshwater cyanobacterium Nostoc sp. strain 152. Appl. Environ. Microbiol. 1990, 56, 2650–2657. [Google Scholar]
- Namikoshi, M.; Sun, F.; Choi, B.W.; Rinehart, K.L.; Carmichael, W.W.; Evans, W.R.; Beasley, V.R. Seven more microcystins from Homer lake cells: Application of the general method for structure assignment of peptides containing dehydroamino acid unit(s). J. Org. Chem. 1995, 60, 3671–3679. [Google Scholar] [CrossRef]
- Foss, A.J.; Miles, C.O.; Samdal, I.A.; Løvberg, K.E.; Wilkins, A.L.; Rise, F.; Jaabæk, J.A.H.; McGowan, P.C.; Aubel, M.T. Analysis of free and metabolized microcystins in samples following a bird mortality event. Harmful Algae 2018, 80, 117–129. [Google Scholar] [CrossRef]
- Yilmaz, M.; Foss, A.J.; Miles, C.O.; Özen, M.; Demir, N.; Balci, M.; Beach, D.G. Comprehensive multi-technique approach reveals the high diversity of microcystins in field collections and an associated isolate of Microcystis aeruginosa from a Turkish lake. Toxicon 2019, 167, 87–100. [Google Scholar] [CrossRef]
- Harada, K.; Ogawa, K.; Matsuura, K.; Nagai, H.; Murata, H.; Suzuki, M.; Itezono, Y.; Nakayama, N.; Shirai, M.; Nakano, M. Isolation of two toxic heptapeptide microcystins from an axenic strain of Microcystis aeruginosa, K-139. Toxicon 1991, 29, 479–489. [Google Scholar] [CrossRef]
- Namikoshi, M.; Sivonen, K.; Evans, W.R.; Carmichael, W.W.; Sun, F.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Two new L-serine variants of microcystins-LR and -RR from Anabaena sp. strains 202 A1 and 202 A2. Toxicon 1992, 30, 1457–1464. [Google Scholar] [CrossRef]
- Sivonen, K.; Skulberg, O.M.; Namikoshi, M.; Evans, W.R.; Carmichael, W.W.; Rinehart, K.L. Two methyl ester derivatives of microcystins, cyclic heptapeptide hepatotoxins, isolated from Anabaena flos-aquae strain CYA 83/1. Toxicon 1992, 30, 1465–1471. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Fardig, M.; Carmichael, W.W.; Rinehart, K.L. Three new microcystins, cyclic heptapeptide hepatotoxins, from Nostoc sp. strain 152. Chem. Res. Toxicol. 1992, 5, 464–469. [Google Scholar] [CrossRef]
- Lee, T.H.; Chou, H.N. Isolation and identification of seven microcystins from a cultured M.TN-2 strain of Microcystis aeruginosa. Bot. Bull. Acad. Sin. 2000, 41, 197–202. [Google Scholar]
- Welker, M.; Christiansen, G.; von Döhren, H. Diversity of coexisting Planktothrix (Cyanobacteria) chemotypes deduced by mass spectral analysis of microcystins and other oligopeptides. Arch. Microbiol. 2004, 182, 288–298. [Google Scholar] [CrossRef]
- Vasas, G.; Szydlowska, D.; Gaspar, A.; Welker, M.; Trojanowicz, M.; Borbély, G. Determination of microcystins in environmental samples using capillary electrophoresis. J. Biochem. Biophys. Methods 2006, 66, 87–97. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Miles, C.O.; Rise, F.; Cary, S.C.; Hamilton, D.P.; Wilkins, A.L. Structural characterization of new microcystins containing tryptophan and oxidized tryptophan residues. Mar. Drugs 2013, 11, 3025–3045. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Cary, S.C.; Hamilton, D.P.; Holland, P.T. Further characterization of glycine-containing microcystins from the McMurdo dry valleys of Antarctica. Toxins 2015, 7, 493–515. [Google Scholar] [CrossRef]
- Wood, S.A.; Mountfort, D.; Selwood, A.I.; Holland, P.T.; Puddick, J.; Cary, S.C. Widespread distribution and identification of eight novel microcystins in Antarctic cyanobacterial mats. Appl. Environ. Microbiol. 2008, 74, 7243–7251. [Google Scholar] [CrossRef]
- Miles, C.O.; Melanson, J.E.; Ballot, A. Sulfide oxidations for LC-MS Analysis of methionine-containing microcystins in Dolichospermum flos-aquae NIVA-CYA 656. Environ. Sci. Technol. 2014, 48, 13307–13315. [Google Scholar] [CrossRef]
- Diehnelt, C.W.; Dugan, N.R.; Peterman, S.M.; Budde, W.L. Identification of microcystin toxins from a strain of Microcystis aeruginosa by liquid chromatography introduction into a hybrid linear ion trap-fourier transform ion cyclotron resonance mass spectrometer. Anal. Chem. 2006, 78, 501–512. [Google Scholar] [CrossRef]
- Niedermeyer, T.H.J.; Daily, A.; Swiatecka-Hagenbruch, M.; Moscow, J.A. Selectivity and potency of microcystin congeners against OATP1B1 and OATP1B3 expressing cancer cells. PLoS ONE 2014, 9, e91476. [Google Scholar] [CrossRef]
- Teta, R.; Della Sala, G.; Glukhov, E.; Gerwick, L.; Gerwick, W.H.; Mangoni, A.; Costantino, V. Combined LC-MS/MS and molecular networking approach reveals new cyanotoxins from the 2014 cyanobacterial bloom in Green Lake, Seattle (WA, USA). Environ. Sci. Technol. 2015, 49, 14301–14310. [Google Scholar] [CrossRef]
- Ortiz, X.; Korenkova, E.; Jobst, K.J.; MacPherson, K.A.; Reiner, E.J. A high throughput targeted and non-targeted method for the analysis of microcystins and anatoxin-a using on-line solid phase extraction coupled to liquid chromatography-quadrupole time-of-flight high resolution mass spectrometry. Anal. Bioanal. Chem. 2017, 409, 4959–4969. [Google Scholar] [CrossRef]
- Meriluoto, J. Chromatography of microcystins. Anal. Chim. Acta 1997, 352, 277–298. [Google Scholar] [CrossRef]
- Mallia, V.; Uhlig, S.; Rafuse, C.; Meija, J.; Miles Christopher, O. Novel microcystins from Planktothrix prolifica NIVA-CYA 544 identified by LC-MS/MS, functional group derivatization and 15N-labeling. Mar. Drugs 2019, 17, 643. [Google Scholar] [CrossRef]
- Kondo, F.; Ikai, Y.; Oka, H.; Okumura, M.; Ishikawa, N.; Harada, K.; Matsuura, K.; Murata, H.; Suzuki, M. Formation, characterization, and toxicity of the glutathione and cysteine conjugates of toxic heptapeptide microcystins. Chem. Res. Toxicol. 1992, 5, 591–596. [Google Scholar] [CrossRef]
- Miles, C.O.; Sandvik, M.; Nonga, H.E.; Ballot, A.; Wilkins, A.L.; Rise, F.; Jaabaek, A.H.; Loader, J.I. Conjugation of microcystins with thiols is reversible: Base-catalyzed deconjugation for chemical analysis. Chem. Res. Toxicol. 2016, 29, 860–870. [Google Scholar] [CrossRef]
- Mayumi, T.; Kato, H.; Imanishi, S.; Kawasaki, Y.; Hasegawa, M.; Harada, K. Structural characterization of microcystins by LC/MS/MS under ion trap conditions. J. Antibiot. 2006, 59, 710–719. [Google Scholar] [CrossRef]
- Stewart, A.K.; Strangman, W.K.; Percy, A.; Wright, J.L.C. The biosynthesis of 15N-labeled microcystins and the comparative MS/MS fragmentation of natural abundance and their 15N-labeled congeners using LC-MS/MS. Toxicon 2018, 144, 91–102. [Google Scholar] [CrossRef]
- Ferranti, P.; Fabbrocino, S.; Nasi, A.; Caira, S.; Bruno, M.; Serpe, L.; Gallo, P. Liquid chromatography coupled to quadrupole time of flight tandem mass spectrometry for microcystin analysis in freshwaters: Method performances and characterisation of a novel variant of microcystin-RR. Rapid Commun. Mass Spectrom. 2009, 23, 1328–1336. [Google Scholar] [CrossRef]
- Dörr, F.A.; Oliveira-Silva, D.; Lopes, N.P.; Iglesias, J.; Volmer, D.A.; Pinto, E. Dissociation of deprotonated microcystin variants by collision-induced dissociation following electrospray ionization. Rapid Commun. Mass Spectrom. 2011, 25, 1981–1992. [Google Scholar] [CrossRef]
- Flores, C.; Caixach, J. An integrated strategy for rapid and accurate determination of free and cell-bound microcystins and related peptides in natural blooms by liquid chromatography–electrospray-high resolution mass spectrometry and matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry using both positive and negative ionization modes. J. Chromatogr. A 2015, 1407, 76–89. [Google Scholar]
- Ngoka, L.C.M.; Gross, M.L. Multistep tandem mass spectrometry for sequencing cyclic peptides in an ion-trap mass spectrometer. J. Am. Soc. Mass Spectrom. 1999, 10, 732–746. [Google Scholar] [CrossRef]
- Miles, C.O.; Sandvik, M.; Nonga, H.E.; Rundberget, T.; Wilkins, A.L.; Rise, F.; Ballot, A. Thiol derivatization for LC-MS identification of microcystins in complex matrices. Environ. Sci. Technol. 2012, 46, 8937–8944. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.O.; Sandvik, M.; Haande, S.; Nonaga, H.; Ballot, A. LC-MS analysis with thiol derivatization to differentiate [Dhb7] from [Mdha7]microcystins: Analysis of cyanobacterial blooms, Planktothrix cultures and European crayfish from Lake Steinsfjorden, Norway. Environ. Sci. Technol. 2013, 47, 4080–4087. [Google Scholar] [CrossRef] [PubMed]
- Zemskov, I.; Kropp, H.M.; Wittmann, V. Regioselective cleavage of thioether linkages in microcystin conjugates. Chem. Eur. J. 2016, 22, 10990–10997. [Google Scholar] [CrossRef]
- Miles, C.O. Rapid and convenient oxidative release of thiol-conjugated forms of microcystins for chemical analysis. Chem. Res. Toxicol. 2017, 30, 1599–1608. [Google Scholar] [CrossRef]
- Harada, K.; Murata, H.; Qiang, Z.; Suzuki, M.; Kondo, F. Mass spectrometric screening method for microcystins in cyanobacteria. Toxicon 1996, 34, 701–710. [Google Scholar] [CrossRef]
- Davidson, I. Hydrolysis of samples for amino acid analysis. In Protein Sequencing Protocols. Methods in Molecular Biology; Smith, B.J., Ed.; Humana Press: Totowa, NJ, USA, 1997; Volume 64. [Google Scholar]
- Luukkainen, R.; Sivonen, K.; Namikoshi, M.; Fardig, M.; Rinehart, K.L.; Niemelä, S.I. Isolation and identification of eight microcystins from thirteen Oscillatoria agardhii strains and structure of a new microcystin. Appl. Environ. Microbiol. 1993, 59, 2204–2209. [Google Scholar]
- Harada, K.; Ogawa, K.; Matsuura, K.; Murata, H.; Suzuki, M.; Watanabe, M.F.; Itezono, Y.; Nakayama, N. Structural determination of geometrical isomers of microcystins LR and RR from cyanobacteria by two-dimensional NMR spectroscopic techniques. Chem. Res. Toxicol. 1990, 3, 473–481. [Google Scholar] [CrossRef]
- Burton, I.W.; Quilliam, M.A.; Walter, J.A. Quantitative 1H NMR with external standards: Use in preparation of calibration solutions for algal toxins and other natural products. Anal. Chem. 2005, 77, 3123–3131. [Google Scholar] [CrossRef]
- National Research Council Canada List of CRM Products. Available online: www.nrc-cnrc.gc.ca/eng/solutions/advisory/crm/list_product.html (accessed on 25 September 2019).
- Goldberg, J.; Huang, H.; Kwon, Y.; Greengard, P.; Nairn, A.C.; Kuriyan, J. Three-dimensional structure of the catalytic subunit of protein serine/threonine phosphatase-1. Nature 1995, 376, 745–753. [Google Scholar] [CrossRef]
- Xing, Y.; Xu, Y.; Chen, Y.; Jeffrey, P.D.; Chao, Y.; Lin, Z.; Li, Z.; Strack, S.; Stock, J.B.; Shi, Y. Structure of protein phosphatase 2A core enzyme bound to tumor-inducing toxins. Cell 2006, 127, 341–353. [Google Scholar] [CrossRef]
- Maynes, J.T.; Luu, H.A.; Cherney, M.M.; Andersen, R.J.; Williams, D.; Holmes, C.F.B.; James, M.N.G. Crystal structures of protein phosphatase-1 bound to motuporin and dihydromicrocystin-LA: Elucidation of the mechanism of enzyme inhibition by cyanobacterial toxins. J. Mol. Biol. 2006, 356, 111–120. [Google Scholar] [CrossRef]
- Cho, U.S.; Xu, W. Crystal structure of a protein phosphatase 2A heterotrimeric holoenzyme. Nature 2007, 445, 53–57. [Google Scholar] [CrossRef]
- Xu, Y.; Chen, Y.; Zhang, P.; Jeffrey, P.D.; Shi, Y. Structure of a protein phosphatase 2A holoenzyme: Insights into B55-mediated tau dephosphorylation. Mol. Cell 2008, 31, 873–885. [Google Scholar] [CrossRef]
- Xu, Z.; Cetin, B.; Anger, M.; Cho, U.S.; Helmhart, W.; Nasmyth, K.; Xu, W. Structure and Function of the PP2A-Shugoshin Interaction. Mol. Cell 2009, 35, 426–441. [Google Scholar] [CrossRef]
- Wlodarchak, N.; Guo, F.; Satyshur, K.A.; Jiang, L.; Jeffrey, P.D.; Sun, T.; Stanevich, V.; Mumby, M.C.; Xing, Y. Structure of the Ca2+-dependent PP2A heterotrimer and insights into Cdc6 dephosphorylation. Cell Res. 2013, 23, 931. [Google Scholar] [CrossRef]
- Chen, E.; Choy, M.S.; Petrényi, K.; Kónya, Z.; Erdődi, F.; Dombrádi, V.; Peti, W.; Page, R. Molecular insights into the fungus-specific serine/threonine protein phosphatase Z1 in Candida albicans. mBio 2016, 7, e00872-16. [Google Scholar] [CrossRef]
- Kumar, G.S.; Choy, M.S.; Koveal, D.M.; Lorinsky, M.K.; Lyons, S.P.; Kettenbach, A.N.; Page, R.; Peti, W. Identification of the substrate recruitment mechanism of the muscle glycogen protein phosphatase 1 holoenzyme. Sci. Adv. 2018, 4, eaau6044. [Google Scholar] [CrossRef]
- Rudolph-Böhner, S.; Mierke, D.F.; Moroder, L. Molecular structure of the cyanobacterial tumor-promoting microcystins. FEBS Lett. 1994, 349, 319–323. [Google Scholar] [CrossRef]
- Bagu, J.R.; Sönnichsen, F.D.; Williams, D.; Andersen, R.J.; Sykes, B.D.; Holmes, C.F.B. Comparison of the solution structures of microcystin-LR and motuporin. Nat. Struct. Mol. Biol. 1995, 2, 114–116. [Google Scholar] [CrossRef]
- Mierke, D.F.; Rudolph-Böhner, S.; Müller, G.; Moroder, L. Structure of two microcystins: Refinement with nuclear Overhauser effects and ensemble calculations. Biopolymers 1995, 36, 811–828. [Google Scholar] [CrossRef]
- Trogen, G.B.; Edlund, U.; Larsson, G.; Sethson, I. The solution NMR structure of a blue-green algae hepatotoxin, microcystin-RR. A comparison with the structure of microcystin-LR. Eur. J. Biochem. 1998, 258, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Sandvik, M.; Rundberget, T.; Botha, C.J.; Miles, C.O. Diversity of cyanobacteria and cyanotoxins in Hartbeespoort Dam, South Africa. Mar. Freshw. Res. 2014, 65, 175–189. [Google Scholar] [CrossRef]
- Mahakhant, A.; Sano, T.; Ratanachot, P.; Tong-a-ram, T.; Srivastava, V.C.; Watanabe, M.M.; Kaya, K. Detection of microcystins from cyanobacterial water blooms in Thailand fresh water. Phycol. Res. 1998, 42, 25–29. [Google Scholar] [CrossRef]
- Craig, M.; McCready, T.L.; Luu, H.A.; Smillie, M.A.; Dubord, P.; Holmes, C.F.B. Identification and characterization of hydrophobic microcystins in Canadian freshwater cyanobacteria. Toxicon 1993, 31, 1541–1549. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Cary, S.C.; Hamilton, D.P.; Wilkins, A.L. Isolation and structure determination of two new hydrophobic microcystins from Microcystis sp. (CAWBG11). Phytochem. Lett. 2013, 6, 575–581. [Google Scholar] [CrossRef]
- Lee, T.H.; Chen, Y.M.; Chou, H.N. First report of microcystins in Taiwan. Toxicon 1998, 36, 247–255. [Google Scholar] [CrossRef]
- Prakash, S.; Lawton, L.A.; Edwards, C. Stability of toxigenic Microcystis blooms. Harmful Algae 2009, 8, 377–384. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Jokela, J.; Fewer, D.P.; Sivonen, K.; Rikkinen, J. Microcystin production in the tripartite cyanolichen Peltigera leucophlebia. Mol. Plant Microbe Interact. 2009, 22, 695–702. [Google Scholar] [CrossRef]
- Namikoshi, M.; Yuan, M.; Sivonen, K.; Carmichael, W.W.; Rinehart, K.L.; Rouhiainen, L.; Sun, F.; Brittain, S.; Otsuki, A. Seven new microcystins possessing two L-glutamic acid units, isolated from Anabaena sp. Strain 186. Chem. Res. Toxicol. 1998, 11, 143–149. [Google Scholar] [CrossRef]
- Robillot, C.; Vinh, J.; Puiseux-Dao, S.; Hennion, M.-C. Hepatotoxin production kinetics of the cyanobacterium Microcystis aeruginosa PCC 7820, as determined by HPLC−mass spectrometry and protein phosphatase bioassay. Environ. Sci. Technol. 2000, 34, 3372–3378. [Google Scholar] [CrossRef]
- Bateman, K.P.; Thibault, P.; Douglas, D.J.; White, R.L. Mass spectral analyses of microcystins from toxic cyanobacteria using on-line chromatographic and electrophoretic separations. J. Chromatogr. A 1995, 712, 253–268. [Google Scholar] [CrossRef]
- Krishnamurthy, T.; Szafraniec, L.; Hunt, D.F.; Shabanowitz, J.; Yates, J.R.; Hauer, C.R.; Carmichael, W.W.; Skulberg, O.; Codd, G.A.; Missler, S. Structural characterization of toxic cyclic peptides from blue-green algae by tandem mass spectrometry. Proc. Nat. Acad. Sci. USA 1989, 86, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Kaya, K. Two new (E)-2-amino-2-butenoic acid (Dhb)-containing microcystins isolated from Oscillatoria agardhii. Tetrahedron 1998, 54, 463–470. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Browarczyk-Matusiak, G.; Forycka, K.; Kobos, J.; Plinski, M. Morphological, genetic, chemical and ecophysiological characterisation of two Microcystis aeruginosa isolates from the Vistula Lagoon, southern Baltic. Oceanologia 2010, 52, 127–146. [Google Scholar] [CrossRef]
- Sano, T.; Takagi, H.; Kaya, K.A. Dhb-microcystin from the filamentous cyanobacterium Planktothrix rubescens. Phytochemistry 2004, 65, 2159–2162. [Google Scholar] [CrossRef]
- Sano, T.; Takagi, H.; Nishikawa, M.; Kaya, K. NIES certified reference material for microcystins, hepatotoxic cyclic peptide toxins from cyanobacterial blooms in eutrophic water bodies. Anal. Bioanal. Chem. 2008, 391, 2005–2010. [Google Scholar] [CrossRef]
- Krishnamurthy, T.; Carmichael, W.W.; Sarver, E.W. Toxic peptides from freshwater cyanobacteria (blue-green algae). I. Isolation, purification and characterization of peptides from Microcystis aeruginosa and Anabaena flos-aquae. Toxicon 1986, 24, 865–873. [Google Scholar] [CrossRef]
- Harada, K.; Matsuura, K.; Suzuki, M.; Watanabe, M.F.; Oishi, S.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W. Isolation and characterization of the minor components associated with microcystins LR and RR in the cyanobacterium (blue-green algae). Toxicon 1990, 28, 55–64. [Google Scholar] [CrossRef]
- Rudolph-Böhner, S.; Wu, J.T.; Moroder, L. Identification and characterization of microcystin-LY from Microcystis aeruginosa (strain 298). Biol. Chem. 1993, 374, 635–640. [Google Scholar]
- Oksanen, I.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Rikkinen, J.; Sivonen, K. Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. strain IO-102-I. Appl. Environ. Microbiol. 2004, 70, 5756–5763. [Google Scholar] [CrossRef] [PubMed]
- Beattie, K.A.; Kaya, K.; Sano, T.; Codd, G.A. Three dehydrobutyrine (Dhb)-containing microcystins from the cyanobacterium Nostoc sp. Phytochemistry 1998, 47, 1289–1292. [Google Scholar] [CrossRef]
- Brittain, S.; Mohamed, Z.A.; Wang, J.; Lehmann, V.K.B.; Carmichael, W.W.; Rinehart, K.L. Isolation and characterization of microcystins from a River Nile strain of Oscillatoria tenuis Agardh ex Gomont. Toxicon 2000, 38, 1759–1771. [Google Scholar] [CrossRef]
- Qi, Y.; Rosso, L.; Sedan, D.; Giannuzzi, L.; Andrinolo, D.; Volmer, D.A. Seven new microcystin variants discovered from a native Microcystis aeruginosa strain—Unambiguous assignment of product ions by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2015, 29, 1–5. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar] [CrossRef]
- Choi, B.W.; Namikoshi, M.; Sun, F.; Rinehart, K.L.; Carmichael, W.W.; Kaup, A.M.; Evans, W.R.; Beasley, V.R. Isolation of linear peptides related to the hepatotoxins nodularin and microcystins. Tetrahedron Lett. 1993, 34, 7881–7884. [Google Scholar] [CrossRef]
- Luukkainen, R.; Namikoshi, M.; Sivonen, K.; Rinehart, K.L.; Niemela, S.I. Isolation and identification of 12 microcystins from four strains and two bloom samples of Microcystis spp.: Structure of a new hepatotoxin. Toxicon 1994, 32, 133–139. [Google Scholar] [CrossRef]
- Meriluoto, J.A.O.; Sandström, A.; Eriksson, J.E.; Remaud, G.; Grey Graig, A.; Chattopadhyaya, J. Structure and toxicity of a peptide hepatotoxin from the cyanobacterium Oscillatoria agardhii. Toxicon 1989, 27, 1021–1034. [Google Scholar] [CrossRef]
- Kiviranta, J.; Namikoshi, M.; Sivonen, K.; Evans, W.R.; Carmichael, W.W.; Rinehart, K.L. Structure determination and toxicity of a new microcystin from Microcystis aeruginosa strain 205. Toxicon 1992, 30, 1093–1098. [Google Scholar] [CrossRef]
- Sano, T.; Kaya, K. A 2-amino-2-butenoic acid (Dhb)-containing microcystin isolated from Oscillatoria agardhii. Tetrahedron Lett. 1995, 36, 8603–8606. [Google Scholar] [CrossRef]
- Kleinteich, J.; Puddick, J.; Wood, S.; Hildebrand, F.; Laughinghouse IV, H.; Pearce, D.; Dietrich, D.; Wilmotte, A. Toxic cyanobacteria in Svalbard: Chemical diversity of microcystins detected using a liquid chromatography mass spectrometry precursor ion screening method. Toxins 2018, 10, 147. [Google Scholar] [CrossRef] [PubMed]
- Gathercole, P.S.; Thiel, P.G. Liquid chromatographic determination of the cyanoginosins, toxins produced by the cyanobacterium Microcystis aeruginosa. J. Chromatogr. 1987, 408, 435–440. [Google Scholar] [CrossRef]
- Namikoshi, M.; Sivonen, K.; Evans, W.R.; Carmichael, W.W.; Rouhiainen, L.; Luukkainen, R.; Rinehart, K.L. Structures of three new homotyrosine-containing microcystins and a new homophenylalanine variant from Anabaena sp. strain 66. Chem. Res. Toxicol. 1992, 5, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Gromov, B.V.; Carmichael, W.W.; Rinehart, K.L. Isolation and structures of five microcystins from a Russian Microcystis aeruginosa strain CALU 972. Toxicon 1992, 30, 1481–1485. [Google Scholar] [CrossRef]
- Okello, W.; Ostermaier, V.; Portmann, C.; Gademann, K.; Kurmayer, R. Spatial isolation favours the divergence in microcystin net production by Microcystis in Ugandan freshwater lakes. Water Res. 2010, 44, 2803–2814. [Google Scholar] [CrossRef]
- Okello, W.; Portmann, C.; Erhard, M.; Gademann, K.; Kurmayer, R. Occurrence of microcystin-producing cyanobacteria in Ugandan fresh waters. Environ. Toxicol. 2010, 25, 367–380. [Google Scholar] [CrossRef]
- Namikoshi, M.; Sivonen, K.; Evans, W.R.; Sun, F.; Carmichael, W.W.; Rinehart, K.L. Isolation and structures of microcystins from a cyanobacterial water bloom (Finland). Toxicon 1992, 30, 1473–1479. [Google Scholar] [CrossRef]
- Elleman, T.; Falconer, I.; Jackson, A.; Runnegar, M. Isolation, characterization and pathology of the toxin from a Microcystis aeruginosa (= Anacystis cyanea) bloom. Aust. J. Biol. Sci. 1978, 31, 209. [Google Scholar] [CrossRef]
- Matthiensen, A.; Beattie, K.A.; Yunes, J.S.; Kaya, K.; Codd, G.A. [D-Leu1]Microcystin-LR, from the cyanobacterium Microcystis RST 9501 and from a Microcystis bloom in the Patos Lagoon estuary, Brazil. Phytochemistry 2000, 55, 383–387. [Google Scholar] [CrossRef]
- Park, H.; Namikoshi, M.; Brittain, S.; Carmichael, W.; Murphy, T. [D-Leu1]Microcystin-LR, a new microcystin isolated from waterbloom in a Canadian prairie lake. Toxicon 2001, 39, 855–862. [Google Scholar] [CrossRef]
- Grach-Pogrebinsky, O.; Sedmak, B.; Carmeli, S. Seco[D-Asp3]microcystin-RR and [D-Asp3,D-Glu(OMe)6]microcystin-RR, two new microcystins from a toxic water bloom of the cyanobacterium Planktothrix rubescens. J. Nat. Prod. 2004, 67, 337–342. [Google Scholar] [CrossRef]
- Roy-Lachapelle, A.; Solliec, M.; Sauvé, S.; Gagnon, C. A data-independent methodology for the structural characterization of microcystins and anabaenopeptins leading to the identification of four new congeners. Toxins 2019, 11, 619. [Google Scholar] [CrossRef] [PubMed]
- Welker, M.; Erhard, M. Consistency between chemotyping of single filaments of Planktothrix rubescens (cyanobacteria) by MALDI-TOF and the peptide patterns of strains determined by HPLC-MS. J. Mass Spectrom. 2007, 42, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Hollingdale, C.; Thomas, K.; Lewis, N.; Bekri, K.; McCarron, P.; Quillam, M.A. Feasibility study on production of a matrix reference material for cyanobacterial toxins. Anal. Bioanal. Chem. 2015, 407, 5353–5363. [Google Scholar] [CrossRef]
- Harada, K.; Ogawa, K.; Kimura, Y.; Murata, H.; Suzuki, M.; Thorn, P.M.; Evans, W.R.; Carmichael, W.W. Microcystins from Anabaena flos-aquae NRC 525-17. Chem. Res. Toxicol. 1991, 4, 535–540. [Google Scholar] [CrossRef]
- Birbeck, J.A.; Peraino, N.J.; O’Neill, G.M.; Coady, J.; Westrick, J.A. Dhb microcystins discovered in USA using an online concentration LC–MS/MS platform. Toxins 2019, 11, 653. [Google Scholar] [CrossRef]
- Christiansen, G.; Yoshida, W.Y.; Blom, J.F.; Portmann, C.; Gademann, K.; Hemscheidt, T.; Kurmayer, R. Isolation and structure determination of two microcystins and sequence comparison of the McyABC adenylation domains in Planktothrix species. J. Nat. Prod. 2008, 71, 1881–1886. [Google Scholar] [CrossRef]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Kupper, F.C.; Dietrich, D. Potent toxins in Arctic environments—Presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystem. Chem. Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef]
- Frias, H.V.; Mendes, M.A.; Cardozo, K.H.; Carvalho, V.M.; Tomazela, D.; Colepicolo, P.; Pinto, E. Use of electrospray tandem mass spectrometry for identification of microcystins during a cyanobacterial bloom event. Biochem. Biophys. Res. Commun. 2006, 344, 741–746. [Google Scholar] [CrossRef]
- Laub, J.; Henriksen, P.; Brittain, S.M.; Wang, J.; Carmichael, W.W.; Rinehart, K.L.; Moestrup, Ø. [ADMAdda5]-microcystins in Planktothrix agardhii strain PH-123 (cyanobacteria)? Importance for monitoring of microcystins in the environment. Environ. Toxicol. 2002, 17, 351–357. [Google Scholar] [CrossRef]
- Rapala, J.; Sivonen, K.; Lyra, C.; Niemela, S.I. Variation of microcystins, cyanobacterial hepatotoxins, in Anabaena spp. as a function of growth stimuli. Appl. Environ. Microbiol. 1997, 63, 2206–2212. [Google Scholar] [PubMed]
- Ndlela, L.L.; Oberholster, P.J.; Van Wyk, J.H.; Cheng, P.H. An overview of cyanobacterial bloom occurrences and research in Africa over the last decade. Harmful Algae 2016, 60, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Barrow, K.D.; Neilan, B.A. Characterization of the 2-hydroxy-acid dehydrogenase McyI, encoded within the microcystin biosynthesis gene cluster of Microcystis aeruginosa PCC7806. J. Biol. Chem. 2007, 282, 4681–4692. [Google Scholar] [CrossRef] [PubMed]
- Sielaff, H.; Dittmann, E.; Tandeau De Marsac, N.; Bouchier, C.; von Döhren, H.; Börner, T.; Schwecke, T. The mcyF gene of the microcystin biosynthetic gene cluster from Microcystis aeruginosa encodes an aspartate racemase. Biochem. J. 2003, 373, 909–916. [Google Scholar] [CrossRef]
- Fewer, D.P.; Wahlsten, M.; Österholm, J.; Jokela, J.; Rouhiainen, L.; Kaasalainen, U.; Rikkinen, J.; Sivonen, K. The genetic basis for O-acetylation of the microcystin toxin in cyanobacteria. Chem. Biol. 2013, 20, 861–869. [Google Scholar] [CrossRef]
- Sieber, S.A.; Marahiel, M.A. Molecular mechanisms underlying nonribosomal peptide synthesis: Approaches to new antibiotics. Chem. Rev. 2005, 105, 715–738. [Google Scholar] [CrossRef]
- Kurmayer, R.; Christiansen, G.; Gumpenberger, M.; Fastner, J. Genetic identification of microcystin ecotypes in toxic cyanobacteria of the genus Planktothrix. Microbiology 2005, 151, 1525–1533. [Google Scholar] [CrossRef]
- Marahiel, M.A.; Stachelhaus, T.; Mootz, H.D. Modular peptide synthetases involved in nonribosomal peptide synthesis. Chem. Rev. 1997, 97, 2651–2674. [Google Scholar] [CrossRef]
- Moffitt, M.C.; Neilan, B.A. Characterization of the nodularin synthetase gene cluster and proposed theory of the evolution of cyanobacterial hepatotoxins. Appl. Environ. Microbiol. 2004, 70, 6353–6362. [Google Scholar] [CrossRef]
- Namikoshi, M.; Choi, B.W.; Sakai, R.; Sun, F.; Rinehart, K.L.; Carmichael, W.W.; Evans, W.R.; Cruz, P.; Munro, M.H.G.; Blunt, J.W. New nodularins: A general method for structure assignment. J. Org. Chem. 1994, 59, 2349–2357. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Namikoshi, M.; Choi, B.W. Structure and biosynthesis of toxins from blue-green algae (cyanobacteria). J. Appl. Phycol. 1994, 6, 159–176. [Google Scholar] [CrossRef]
- Tsuji, K.; Watanuki, T.; Kondo, F.; Watanabe, M.F.; Suzuki, S.; Nakazawa, H.; Suzuki, M.; Uchida, H.; Harada, K.I. Stability of microcystins from cyanobacteria-II. Effect of UV light on decomposition and isomerization. Toxicon 1995, 33, 1619–1631. [Google Scholar] [CrossRef]
- Kaya, K.; Sano, T.A. Photodetoxification mechanism of the cyanobacterial hepatotoxin microcystin-lr by ultraviolet irradiation. Chem. Res. Toxicol. 1998, 11, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Bardowell, S.; O’Shea, K.E. Mechanistic study and the influence of oxygen on the photosensitized transformations of microcystins (cyanotoxins). Environ. Sci. Technol. 2007, 41, 5336–5341. [Google Scholar] [CrossRef]
- Fastner, J.; Erhard, M.; von Döhren, H. Determination of oligopeptide diversity within a natural population of Microcystis spp. (cyanobacteria) by typing single colonies by matrix-assisted laser desorption ionization-time of flight mass spectrometry. Appl. Environ. Microbiol. 2001, 67, 5069–5076. [Google Scholar] [CrossRef]
- Buratti, F.M.; Scardala, S.; Funari, E.; Testai, E. The conjugation of microcystin-RR by human recombinant GSTs and hepatic cytosol. Toxicol. Lett. 2013, 219, 231–238. [Google Scholar] [CrossRef]
- Dai, M.; Xie, P.; Liang, G.; Chen, J.; Lei, H. Simultaneous determination of microcystin-LR and its glutathione conjugate in fish tissues by liquid chromatography–tandem mass spectrometry. J. Chromatogr. B 2008, 862, 43–50. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Puerto, M.; Gutiérrez-Praena, D.; Llana-Ruiz-Cabello, M.; Jos, A.; Cameán, A.M. Microcystin-RR: Occurrence, content in water and food and toxicological studies. A review. Environ. Res. 2019, 168, 467–489. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Zechmann, B.; Tomašić, A.; Horvat, L.; Fulgosi, H. Subcellular distribution of glutathione and cysteine in cyanobacteria. Protoplasma 2010, 246, 65–72. [Google Scholar] [CrossRef]
- Meister, A. Glutathione metabolism and its selective modification. J. Biol. Chem. 1988, 263, 17205–17208. [Google Scholar] [PubMed]
- Li, J.; Li, R.; Li, J. Current research scenario for microcystins biodegradation—A review on fundamental knowledge, application prospects and challenges. Sci. Total Environ. 2017, 595, 615–632. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, C.T.; Mason, M.F. Observations on the pathological changes produced by a toxic substance present in blue-green algae (Microcystis aeruginosa). Am. J. Pathol. 1946, XXII, 369–383. [Google Scholar]
- McBarron, E.J.; May, V. Poisoning of sheep in New South Wales by the blue-green alga Anacystis cyanea (Kuetz.) Dr. And dail. Aust. Vet. J. 1966, 42, 449–453. [Google Scholar] [CrossRef]
- Fawell, J.K.; James, C.P.; James, H.A. Toxins from Blue-Green Algae: Toxicological Assessment of Microcystin-LR and a Method for Its Determination in Water; Report No: FR 0359/2/DoE 3358/2; Foundation for Water Research: Denver, CO, USA, 1994. [Google Scholar]
- Kuiper-Goodman, T.; Falconer, I.R.; Fitzgerald, J. Human health aspects. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E & FN Spon: New York, NY, USA, 1999; pp. 113–153. [Google Scholar]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; E & FN Spon: New York, NY, USA, 1999. [Google Scholar]
- Stoner, R.D.; Adams, W.H.; Slatkin, D.N.; Siegelman, H.W. The effects of single L-amino acid substitutions on the lethal potencies of the microcystins. Toxicon 1989, 27, 825–828. [Google Scholar] [CrossRef]
- Creasia, D.A. Acute inhalation toxicity of microcystin-LR with mice. Toxicon 1990, 28, 605. [Google Scholar] [CrossRef]
- Miura, G.A.; Robinson, N.A.; Lawrence, W.B.; Pace, J.G. Hepatotoxicity of microcystin-LR in fed and fasted rats. Toxicon 1991, 29, 337–346. [Google Scholar] [CrossRef]
- Ito, E.; Kondo, F.; Harada, K.-I. Intratracheal administration of microcystin-LR, and its distribution. Toxicon 2001, 39, 265–271. [Google Scholar] [CrossRef]
- Fawell, J.K.; Mitchell, R.E.; Everett, D.J.; Hill, R.E. The toxicity of cyanobacterial toxins in the mouse: I Microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef]
- Smith, J.L.; Schulz, K.L.; Zimba, P.V.; Boyer, G.L. Possible mechanism for the foodweb transfer of covalently bound microcystins. Ecotoxicol. Environ. Saf. 2010, 73, 757–761. [Google Scholar] [CrossRef]
- Freitas, M.; Azevedo, J.; Carvalho, A.P.; Campos, A.; Vasconcelos, V. Effects of storage, processing and proteolytic digestion on microcystin-LR concentration in edible clams. Food Chem. Toxicol. 2014, 66, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Gulledge, B.M.; Aggen, J.B.; Eng, H.; Sweimeh, K.; Chamberlin, A.R. Microcystin analogues comprised only of Adda and a single additional amino acid retain moderate activity as PP1/PP2A inhibitors. Bioorg. Med. Chem. Lett. 2003, 13, 2907–2911. [Google Scholar] [CrossRef]
- Gulledge, B.M.; Aggen, J.B.; Chamberlin, A.R. Linearized and truncated microcystin analogues as inhibitors of protein phosphatases 1 and 2A. Bioorg. Med. Chem. Lett. 2003, 13, 2903–2906. [Google Scholar] [CrossRef]
- Blom, J.F.; Jüttner, F. High crustacean toxicity of microcystin congeners does not correlate with high protein phosphatase inhibitory activity. Toxicon 2005, 46, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-M.; Lee, T.-H.; Lee, S.-J.; Huang, H.-B.; Huang, R.; Chou, H.-N. Comparison of protein phosphatase inhibition activities and mouse toxicities of microcystins. Toxicon 2006, 47, 742–746. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- Matsushima, R.; Yoshizawa, S.; Watanabe, M.F.; Harada, K.; Furusawa, M.; Carmichael, W.W.; Fujiki, H. In vitro and in vivo effects of protein phosphatase inhibitors, microcystins and nodularin, on mouse skin and fibroblasts. Biochem. Bioph. Res. Commun. 1990, 171, 867–874. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystis and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. 1990, 116, 609–614. [Google Scholar] [CrossRef]
- Honkanen, R.E.; Zwiller, J.; Daily, S.; Khatra, B.S.; Dukelow, M.; Boynton, A.L. Identification, purification, and characterization of novel serine/threonine protein phosphatase from bovine brain. J. Biol. Chem. 1991, 266, 6614–6619. [Google Scholar]
- Runnegar, M.; Berndt, N.; Kong, S.M.; Lee, E.Y.C.; Zhang, L.F. In vivo and in vitro binding of microcystin to protein phosphatase 1 and 2A. Biochem. Biophys. Res. Commun. 1995, 216, 162–169. [Google Scholar] [CrossRef]
- Dawson, R. The toxicology of microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef]
- Guzman, R.E.; Solter, P.F.; Runnegar, M.T. Inhibition of nuclear protein phosphatase activity in mouse hepatocytes by the cyanobacterial toxin microcystin-LR. Toxicon 2003, 41, 773–781. [Google Scholar] [CrossRef]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Monks, N.R.; Liu, S.; Xu, Y.; Yu, H.; Bendelow, A.S.; Moscow, J.A. Potent cytotoxicity of the phosphatase inhibitor microcystin-LR and microcystin analogues in OATP1B1- and OATP1B3-expressing HeLa cells. Mol. Cancer Ther. 2007, 6, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Kaya, K.; Watanabe, M.M. Microcystin composition of an axenic clonal strain of Microcystis viridis and Microcystis viridis-containing waterblooms in Japanese freshwaters. J. Appl. Phycol. 1990, 2, 173–178. [Google Scholar] [CrossRef]
- Zurawell, R.W.; Chen, H.; Burke, J.M.; Prepas, E.E. Hepatotoxic cyanobacteria: A review of the biological importance of microcystins in freshwater environments. J. Toxicol. Environ. Health Part B 2005, 8, 1–37. [Google Scholar] [CrossRef]
- MacKintosh, R.W.; Dalby, K.N.; Campbell, D.G.; Cohen, P.T.W.; Cohen, P.; MacKintosh, C. The cyanobacterial toxin microcystin binds covalently to cysteine-273 on protein phosphatase 1. FEBS Lett. 1995, 371, 236–240. [Google Scholar] [CrossRef]
- Craig, M.; Luu, H.A.; McCready, T.L.; Holmes, C.F.B.; Williams, D.; Andersen, R.J. Molecular mechanisms underlying the interaction of motuporin and microcystins with type-1 and type-2A protein phosphatases. Biochem. Cell Biol. 1996, 74, 569–578. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; Long, F.; Lee, E.Y.C. Tyrosine-272 is involved in the inhibition of protein phosphatase-1 by multiple toxins. Biochemistry 1996, 35, 1606–1611. [Google Scholar] [CrossRef]
- Dawson, J.F.; Holmes, C.F.B. Molecular mechanisms underlying inhibition of protein phosphatases by marine toxins. Front. Biosci. 1999, 4, 646–658. [Google Scholar] [CrossRef]
- Chen, M.X.; McPartlin, A.E.; Brown, L.; Chen, Y.H.; Barker, H.M.; Cohen, P.T. A novel human protein serine/threonine phosphatase, which possesses four tetratricopeptide repeat motifs and localizes to the nucleus. EMBO J. 1994, 13, 4278–4290. [Google Scholar] [CrossRef] [PubMed]
- Hastie, C.J.; Cohen, P.T.W. Purification of protein phosphatase 4 catalytic subunit: Inhibition by the antitumour drug fostriecin and other tumour suppressors and promoters. FEBS Lett. 1998, 431, 357–361. [Google Scholar] [CrossRef]
- Prickett, T.D.; Brautigan, D.L. The α4 Regulatory subunit exerts opposing allosteric effects on protein phosphatases PP6 and PP2A. J. Biol. Chem. 2006, 281, 30503–30511. [Google Scholar] [CrossRef] [PubMed]
- Swingle, M.; Ni, L.; Honkanen, R.E. Small-molecule inhibitors of Ser/Thr protein phosphatases: Specificity, use and common forms of abuse. In Methods in Molecular Biology: Protein Phosphatase Protocols; Ludlow, J.W., Ed.; Humana Press: Totowa, NJ, USA, 2006; Volume 365, pp. 23–38. [Google Scholar]
- Huang, X.; Honkanen, R.E. Molecular cloning, expression, and characterization of a novel human Serine/Threonine protein phosphatase, PP7, that is homologous to Drosophila retinal degeneration C gene product (rdgC). J. Biol. Chem. 1998, 273, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, R.E.; Codispoti, B.A.; Tse, K.; Boynton, A.L. Characterization of natural toxins with inhibitory activity against serine/threonine protein phosphatases. Toxicon 1994, 32, 339–350. [Google Scholar] [CrossRef]
- Robillot, C.; Hennion, M.-C. Issues arising when interpreting the results of the protein phosphatase 2A inhibition assay for the monitoring of microcystins. Anal. Chim. Acta 2004, 512, 339–346. [Google Scholar] [CrossRef]
- Ikehara, T.; Imamura, S.; Sano, T.; Nakashima, J.; Kuniyoshi, K.; Oshiro, N.; Yoshimoto, M.; Yasumoto, T. The effect of structural variation in 21 microcystins on their inhibition of PP2A and the effect of replacing Cys269 with glycine. Toxicon 2009, 54, 539–544. [Google Scholar] [CrossRef]
- An, J.; Carmichael, W.W. Use of a colorimetric protein phosphatase inhibition assay and enzyme-linked immunosorbent assay for the study of microcystins and nodularins. Toxicon 1994, 32, 1495–1507. [Google Scholar] [CrossRef]
- Ufelmann, H.; Krüger, T.; Luckas, B.; Schrenk, D. Human and rat hepatocyte toxicity and protein phosphatase 1 and 2A inhibitory activity of naturally occurring desmethyl-microcystins and nodularins. Toxicology 2012, 293, 59–67. [Google Scholar] [CrossRef]
- Suganuma, M.; Fujiki, H.; Okabe, S.; Nishiwaki, S.; Brautigan, D.; Ingebritsen, T.S.; Rosner, M.R. Structurally different members of the okadaic acid class selectively inhibit protein serine/threonine but not tyrosine phosphatase activity. Toxicon 1992, 30, 873–878. [Google Scholar] [CrossRef]
- Holmes, C.F.B.; Boland, M.P. Inhibitors of protein phosphatase-1 and -2A; two of the major serine/threonine protein phosphatases involved in cellular regulation. Curr. Opin. Struct. Biol. 1993, 3, 934–943. [Google Scholar] [CrossRef]
- Wirsing, B.; Flury, T.; Wiedner, C.; Neumann, U.; Weckesser, J. Estimation of the microcystin content in cyanobacterial field samples from German lakes using the colorimetric protein-phosphatase inhibition assay and RP-HPLC. Environ. Toxicol. 1999, 14, 23–29. [Google Scholar] [CrossRef]
- Catanante, G.; Espin, L.; Marty, J.-L. Sensitive biosensor based on recombinant PP1α for microcystin detection. Biosens. Bioelectron. 2015, 67, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.E.; Toivola, D.; Meriluoto, J.A.O.; Karaki, H.; Han, Y.-G.; Hartshorne, D. Hepatocyte deformation induced by cyanobacterial toxins reflects inhibition of protein phosphatases. Biochem. Biophys. Res. Commun. 1990, 173, 1347–1353. [Google Scholar] [CrossRef]
- Xu, L.; Lam, P.K.S.; Chen, J.; Zhang, Y.; Harada, K. Comparative study on in vitro inhibition of grass carp (Ctenopharyngodon idellus) and mouse protein phosphatases by microcystins. Environ. Toxicol. 2000, 15, 71–75. [Google Scholar] [CrossRef]
- Heresztyn, T.; Nicholson, B.C. Determination of cyanobacterial hepatotoxins directly in water using a protein phosphatase inhibition assay. Water Res. 2001, 35, 3049–3056. [Google Scholar] [CrossRef]
- Rivasseau, C.; Racaud, P.; Deguin, A.; Hennion, M.-C. Development of a bioanalytical phosphatase inhibition test for the monitoring of microcystins in environmental water samples. Anal. Chim. Acta 1999, 394, 243–257. [Google Scholar] [CrossRef]
- Mountfort, D.O.; Holland, P.; Sprosen, J. Method for detecting classes of microcystins by combination of protein phosphatase inhibition assay and ELISA: Comparison with LC-MS. Toxicon 2005, 45, 199–206. [Google Scholar] [CrossRef]
- Hastie, C.; Borthwick, E.; Morrison, L.; Codd, G.; Cohen, P. Inhibition of several protein phosphatases by a non-covalently interacting microcystin and a novel cyanobacterial peptide, nostocyclin. BBA Gen. Subj. 2005, 1726, 187–193. [Google Scholar] [CrossRef]
- Nishiwaki-Matsushima, R.; Nishiwaki, S.; Ohta, T.; Yoshizawa, S.; Suganuma, M.; Harada, K.-I.; Watanabe, M.F.; Fujiki, H. Structure-function relationships of microcystins, liver tumor promoters, in interaction with protein phosphatase. Jpn. J. Cancer Res. 1991, 82, 993–996. [Google Scholar] [CrossRef]
- Wong, B.S.F.; Lam, P.K.S.; Xu, L.; Zhang, Y.; Richardson, B.J. A colorimetric assay for screening microcystin class compounds in aquatic systems. Chemosphere 1999, 38, 1113–1122. [Google Scholar] [CrossRef]
- Hoeger, S.J.; Schmid, D.; Blom, J.F.; Ernst, B.; Dietrich, D.R. Analytical and functional characterization of microcystins [Asp3]MC-RR and [Asp3,Dhb7]MC-RR: Consequences for risk assessment? Environ. Sci. Technol. 2007, 41, 2609–2616. [Google Scholar] [CrossRef] [PubMed]
- Bagu, J.R.; Sykes, B.D.; Craig, M.M.; Holmes, C.F.B. A molecular basis for different interactions of marine toxins with protein phosphatase-1: Molecular models for bound motuporin, microcystins, okadaic acid, and calyculin A. J. Biol. Chem. 1997, 272, 5087–5097. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.R.; Vasconcelos, V.M.; Antunes, A. The phosphoprotein phosphatase family of Ser/Thr phosphatases as principal targets of naturally occurring toxins. Crit. Rev. Toxicol. 2011, 41, 83–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xing, Y.; Chen, Y.; Chao, Y.; Lin, Z.; Fan, E.; Yu, J.W.; Strack, S.; Jeffrey, P.D.; Shi1, Y. Structure of the protein phosphatase 2A holoenzyme. Cell 2006, 127, 1239–1251. [Google Scholar] [CrossRef]
- Kelker, M.S.; Page, R.; Peti, W. Crystal structures of protein phosphatase-1 bound to nodularin-R and tautomycin: A novel scaffold for structure-based drug design of serine/threonine phosphatase inhibitors. J. Mol. Biol. 2009, 385, 11–21. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Entry | Microcystin | Molecular Formula | Exact Mass a | Characterization | Reference |
|---|---|---|---|---|---|
| 1 | [D-Asp³,DMAdda5]MC-LA | C44H63N7O12 | 881.4535 | LC-MS/MS | [33] |
| 2 | [D-Asp³]MC-VA | C44H63N7O12 | 881.4535 | LC-MS/MS | [33] |
| 3 | [D-Asp3]MC-LA | C45H65N7O12 | 895.4691 | LC-HRMS/MS | [94] |
| 4 | [Dha7]MC-LA | C45H65N7O12 | 895.4691 | LC-MS/MS | [33] |
| 5 | [DMAdda5]MC-LA | C45H65N7O12 | 895.4691 | LC–MS/MS, thiol | [131] |
| 6 | MC-VA | C45H65N7O12 | 895.4691 | LC-MS/MS | [33] |
| 7 | MC-LA | C46H67N7O12 | 909.4848 | MS, NMR, AA, LC-HRMS/MS | [15,94] |
| 8 | MC-LAbu | C47H69N7O12 | 923.5004 | HRFABMS, AA, NMR | [132] |
| 9 | [D-Asp3,D-Glu(OMe)6]MC-LAbu b | C47H69N7O12 | 923.5004 | LC-HRMS/MS | [38] |
| 10 | [D-Asp³]MC-LV | C47H69N7O12 | 923.5004 | LC-MS/MS | [33] |
| 11 | [D-Asp³]MC-FA | C48H63N7O12 | 929.4535 | LC-MS/MS, thiol | [12] |
| 12 | [D-Asp³,Dha7]MC-YA | C47H61N7O13 | 931.4327 | MALDI-TOF MS | [89] |
| 13 | [D-Asp3]MC-LL | C48H71N7O12 | 937.5161 | LC-HRMS/MS | [94] |
| 14 | MC-LV | C48H71N7O12 | 937.5161 | AA, LC-MS/MS | [33,133] |
| 15 | [D-Asp³]MC-RA | C45H66N10O12 | 938.4862 | LC-MS/MS, thiol | [12,33] |
| 16 | MC-FA | C49H65N7O12 | 943.4691 | NMR, LC-MS/MS, AA, thiol | [134] |
| 17 | MC-LL | C49H73N7O12 | 951.5317 | LC-HRMS/MS, AA | [94,133] |
| 18 | MC-AR | C46H68N10O12 | 952.5018 | AA, HRMS, NMR | [76] |
| 19 | MC-RA | C46H68N10O12 | 952.5018 | LC-MS/MS, NMR, thiol | [77,135] |
| 20 | [D-Asp³]MC-RAbu | C46H68N10O12 | 952.5018 | LC-MS/MS, thiol | [12] |
| 21 | MC-FAbu | C50H67N7O12 | 957.4848 | LC-MS/MS, thiol | [12] |
| 22 | MC-YA | C49H65N7O13 | 959.4640 | AA, NMR, MS, LC-MS/MS, thiol | [16,77] |
| 23 | MC-AHar | C47H70N10O12 | 966.5175 | LC-MS/MS | [136] |
| 24 | [Gly1,D-Asp3]MC-LR | C47H70N10O12 | 966.5175 | LC-MS/MS | [92] |
| 25 | [D-Asp3,Dha7]MC-LR | C47H70N10O12 | 966.5175 | AA, FABMS/MS | [84] |
| 26 | [D-Asp³,DMAdda5]MC-LR | C47H70N10O12 | 966.5175 | LC-MS/MS | [137] |
| 27 | [D-Asp³,DMAdda5,Dhb7]MC-LR | C47H70N10O12 | 966.5175 | LC-MS/MS, thiol | [109] |
| 28 | [Gly¹,D-Asp3,Dhb7]MC-LR | C47H70N10O12 | 966.5175 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 29 | MC-RAbu | C47H70N10O12 | 966.5175 | LC-MS/MS, thiol | [77] |
| 30 | [D-Asp3]MC-HarAbu | C47H70N10O12 | 966.5175 | LC-MS/MS | [40] |
| 31 | [D-Asp³]MC-WA | C50H64N8O12 | 968.4644 | LC-MS/MS, thiol | [12] |
| 32 | [D-Asp3,Dha7]MC-EE(OMe) b | C46H63N7O16 | 969.4331 | HRMS, MS/MS | [138] |
| 33 | [D-Asp³]MC-EE | C46H63N7O16 | 969.4331 | LC-HRMS/MS, thiol, esterification, 15N-label | [99] |
| 34 | MC-LM | C48H71N7O12S | 969.4881 | AA, MS | [133] |
| 35 | [D-Asp3]MC-LF | C51H69N7O12 | 971.5004 | LC-MS, MS/MS, LC-MS/MS, thiol | [108,139] |
| 36 | MC-VF | C51H69N7O12 | 971.5004 | LC-MS/MS, 15N-enrichment | [140] |
| 37 | [D-Asp3,Dha7]MC-LY | C50H67N7O13 | 973.4797 | LC-MS/MS, thiol | [108] |
| 38 | MC-YAbu | C50H67N7O13 | 973.4797 | LC-MS/MS, thiol | [77] |
| 39 | [D-Asp3]MC-LR | C48H72N10O12 | 980.5331 | MS/MS, HRMS, AA | [141] |
| 40 | [D-Asp3,(E)-Dhb7]MC-LR | C48H72N10O12 | 980.5331 | NMR, AA, HRMS | [142] |
| 41 | [D-Asp3,(Z)-Dhb7]MC-LR | C48H72N10O12 | 980.5331 | NMR, AA, HRMS | [142] |
| 42 | [Dha7]MC-LR | C48H72N10O12 | 980.5331 | AA, FABMS/MS | [84] |
| 43 | [DMAdda5]MC-LR | C48H72N10O12 | 980.5331 | AA, HRMS, NMR | [76] |
| 44 | [Gly1,D-Asp3,Dhb7]MC-LHar | C48H72N10O12 | 980.5331 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 45 | MC-RApa | C48H72N10O12 | 980.5331 | LC-MS/MS, thiol | [77] |
| 46 | MC-VR | C48H72N10O12 | 980.5331 | LC-MS/MS | [143] |
| 47 | MC-WA | C51H66N8O12 | 982.4800 | NMR, LC-MS/MS, AA, thiol | [134] |
| 48 | [D-Ser1,D-Asp³,Dha7]MC-LR | C47H70N10O13 | 982.5124 | LC-MS/MS | [33] |
| 49 | [Dha7]MC-EE(OMe) b | C47H65N7O16 | 983.4488 | HRMS, MS/MS | [138] |
| 50 | [D-Asp3,Dha7]MCE(OMe)E(OMe) b | C47H65N7O16 | 983.4488 | HRMS, MS/MS | [138] |
| 51 | MC-FL | C52H71N7O12 | 985.5161 | LC-MS/MS, thiol | [12] |
| 52 | MC-LF | C52H71N7O12 | 985.5161 | AA, MS | [133] |
| 53 | MC-KynA b | C50H66N8O13 | 986.4749 | LC-MS/MS, HRMS, thiol, semisynthesis | [90] |
| 54 | [D-Asp³]MC-LY | C51H69N7O13 | 987.4953 | LC-MS/MS, thiol | [108] |
| 55 | [D-Asp3,(E)-Dhb7]MC-LY | C51H69N7O13 | 987.4953 | NMR, LC-HRMS/MS, thiol | [109] |
| 56 | [Gly1,D-Asp3,ADMAdda5]MC-LR | C48H70N10O13 | 994.5124 | LC-MS/MS | [92] |
| 57 | [Gly1,D-Asp³,ADMAdda5,Dhb7]MC-LR | C48H70N10O13 | 994.5124 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 58 | [D-Asp³,ADMAdda5]MC-VR | C48H70N10O13 | 994.5124 | LC-MS/MS | [33] |
| 59 | [D-Asp³,Dhb7]MC-AhaR | C49H74N10O12 | 994.5488 | LC-MS/MS, thiol | [109] |
| 60 | [D-Asp³]MC-Hil/HleR | C49H74N10O12 | 994.5488 | LC-MS/MS | [33] |
| 61 | [D-Asp³,(E)-Dhb7]MC-HilR | C49H74N10O12 | 994.5488 | NMR, HRMS, AA | [144] |
| 62 | [Dha7]MC-HilR | C49H74N10O12 | 994.5488 | HRMS, NMR, AA | [145] |
| 63 | [DMAdda5]MC-HilR | C49H74N10O12 | 994.5488 | LC-MS/MS | [33] |
| 64 | [DMAdda5]MC-LHar | C49H74N10O12 | 994.5488 | LC-MS/MS | [33] |
| 65 | [D-Asp3,D-Glu(OMe)6]MC-LR b | C49H74N10O12 | 994.5488 | HRMS, MS/MS, AA | [85] |
| 66 | MC-LR | C49H74N10O12 | 994.5488 | AA, NMR, HRMS, LC-MS/MS | [16,141,146] |
| 67 | [D-Asp3]MC-ER | C47H68N10O14 | 996.4916 | LC-HRMS/MS, thiol, esterification, 15N-label | [99] |
| 68 | [(6Z)-Adda5]MC-LR b | C49H74N10O12 | 994.5488 | NMR, AA, MS | [115,147] |
| 69 | MC-RL | C49H74N10O12 | 994.5488 | LC-MS/MS, thiol | [77] |
| 70 | MC-WAbu | C52H68N8O12 | 996.4957 | LC-MS/MS, thiol | [90] |
| 71 | [Dha7]MC-E(OMe)E(OMe) b | C48H67N7O16 | 997.4644 | HRMS, MS/MS | [138] |
| 72 | MC-OiaA b | C51H66N8O13 | 998.4749 | LC-MS/MS, HRMS, thiol, semisynthesis | [90] |
| 73 | [D-Asp³]MC-MR | C47H70N10O12S | 998.4895 | LC-MS/MS, thiol, S-oxidation | [93] |
| 74 | [seco-4/5][D-Asp³]MC-LR b | C48H74N10O13 | 998.5437 | LC-MS/MS, thiol | [93] |
| 75 | [D-Asp³,Mser7]MC-LR | C48H74N10O13 | 998.5437 | LC-MS/MS, MS/MS, thiol | [88,93] |
| 76 | [Ser7]MC-LR | C48H74N10O13 | 998.5437 | AA, HRMS, MS/MS | [84] |
| 77 | MC-LHph | C53H73N7O12 | 999.5317 | LC-MS/MS | [75] |
| 78 | MC-KynAbu b | C51H68N8O13 | 1000.4906 | LC-MS/MS, thiol | [90] |
| 79 | [D-Asp³,Dha7]MC-FR | C50H68N10O12 | 1000.5018 | LC-MS/MS | [33] |
| 80 | [Ser7]MC-EE(OMe) b | C47H67N7O17 | 1001.4593 | HRMS, MS/MS | [138] |
| 81 | [D-Asp3,Ser7]MC-E(OMe)E(OMe) b | C47H67N7O17 | 1001.4593 | HRMS, MS/MS | [138] |
| 82 | [D-Asp³]MC-HilY | C52H71N7O13 | 1001.5110 | LC-MS/MS, thiol | [108] |
| 83 | MC-LY | C52H71N7O13 | 1001.5110 | LC-MS/MS, NMR | [38,148] |
| 84 | MC-YL | C52H71N7O13 | 1001.5110 | LC-MS/MS | [38] |
| 85 | [D-Asp³,Mser7]MC-LY | C51H71N7O14 | 1005.5059 | LC-MS/MS, thiol | [108] |
| 86 | [D-Asp³,ADMAdda5,Dha7]MC-HilR | C49H72N10O13 | 1008.5280 | LC-MS/MS | [33] |
| 87 | [Gly1,D-Asp3,ADMAdda5,Dhb7]MC-LHar | C49H72N10O13 | 1008.5280 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 88 | [Gly1,D-Asp3,ADMAdda5]MC-LHar | C49H72N10O13 | 1008.5280 | LC-MS/MS | [92] |
| 89 | [D-Asp3,ADMAdda5]MC-LR | C49H72N10O13 | 1008.5280 | HRMS, NMR, AA, MS/MS, LC-MS/MS | [78,79,149] |
| 90 | [ADMAdda5,Dha7]MC-LR | C49H72N10O13 | 1008.5280 | LC-MS/MS | [33] |
| 91 | [D-Asp³,ADMAdda5,Dhb7]MC-LR | C49H72N10O13 | 1008.5280 | NMR, HRMS, AA | [150] |
| 92 | MC-HilR | C50H76N10O12 | 1008.5644 | MS/MS, HRMS, NMR, AA | [80] |
| 93 | MC-LHar | C50H76N10O12 | 1008.5644 | AA, MS/MS, HRMS, NMR | [151] |
| 94 | [D-Glu(OMe)6]MC-LR b | C50H76N10O12 | 1008.5644 | HRMS, MS/MS, AA, HRMS/MS | [85,152] |
| 95 | [Mdhb7]MC-LR | C50H76N10O12 | 1008.5644 | AA, MS | [79] |
| 96 | [D-Leu1,D-Asp³,DMAdda5]MC-LR | C50H76N10O12 | 1008.5644 | LC-MS/MS | [153] |
| 97 | [D-Asp3,Dha7]MC-RR | C47H71N13O12 | 1009.5345 | AA, NMR, HRMS | [76] |
| 98 | [D-Asp³,DMAdda5]MC-RR | C47H71N13O12 | 1009.5345 | MS, MS/MS | [55] |
| 99 | [Gly1,D-Asp3]MC-RR | C47H71N13O12 | 1009.5345 | LC-MS/MS | [92] |
| 100 | [Gly1,D-Asp3,Dhb7]MC-RR | C47H71N13O12 | 1009.5345 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 101 | [D-Asp3]MC-LW | C53H70N8O12 | 1010.5113 | LC-MS, MS/MS | [139] |
| 102 | [D-Asp3,(E)-Dhb7]MC-LW | C53H70N8O12 | 1010.5113 | NMR, LC-HRMS/MS | [95] |
| 103 | MC-OiaAbu b | C52H68N8O13 | 1012.4906 | LC-MS/MS, thiol | [90] |
| 104 | MC-MR | C48H72N10O12S | 1012.5052 | LC-MS/MS, thiol, S-oxidation | [93] |
| 105 | [Mser7]MC-LR | C49H76N10O13 | 1012.5593 | LC-HRMS | [80,94] |
| 106 | [seco-4/5]MC-LR b | C49H76N10O13 | 1012.5593 | LC-MS/MS, HRMS, thiol, NMR | [93,154] |
| 107 | [seco-1/2]MC-LR b | C49H76N10O13 | 1012.5593 | MS/MS, HRMS, NMR | [154] |
| 108 | MC-NfkA b | C51H66N8O14 | 1014.4698 | NMR, LC-MS/MS, HRMS, thiol, semisynthesis | [90] |
| 109 | [D-Asp³]MC-M(O)R b | C47H70N10O13S | 1014.4845 | LC-MS/MS, thiol, S-oxidation | [93] |
| 110 | [D-Asp³,Dha7]MC-HphR | C51H70N10O12 | 1014.5175 | LC-MS/MS | [33] |
| 111 | [D-Asp3]MC-FR | C51H70N10O12 | 1014.5175 | AA, MS, NMR | [87] |
| 112 | [Dha7]MC-FR | C51H70N10O12 | 1014.5175 | AA, HRMS, MS/MS | [155] |
| 113 | [DMAdda5]MC-FR | C51H70N10O12 | 1014.5175 | LC-MS/MS | [137] |
| 114 | [D-Asp³]MC-RF | C51H70N10O12 | 1014.5175 | LC-MS/MS, thiol | [108] |
| 115 | [Ser7]MC-E(OMe)E(OMe) b | C48H69N7O17 | 1015.4750 | HRMS, MS/MS | [138] |
| 116 | MC-LHty | C53H73N7O13 | 1015.5266 | LC-MS/MS | [75] |
| 117 | [D-Asp³,Dha7]MC-RY | C50H68N10O13 | 1016.4967 | LC-MS/MS, thiol | [108] |
| 118 | [D-Asp³,DMAdda5]MC-RY | C50H68N10O13 | 1016.4967 | LC-MS/MS, thiol | [109] |
| 119 | MC-YM | C51H69N7O13S | 1019.4674 | AA, NMR, MS | [16] |
| 120 | [D-Asp3,ADMAdda5]MC-LHar | C50H74N10O13 | 1022.5437 | HRMS, MS/MS, AA | [86] |
| 121 | [ADMAdda5]MC-LR | C50H74N10O13 | 1022.5437 | HRMSNMR, AA, MS/MS | [78,79] |
| 122 | [D-Leu1,DMAdda5]MC-LR | C51H78N10O12 | 1022.5801 | LC-HRMS/MS, thiol | [81] |
| 123 | [D-Leu1,dmAdda5]MC-LR (isomer 1) c | C51H78N10O12 | 1022.5801 | LC-HRMS/MS, thiol | [81] |
| 124 | [D-Leu1,dmAdda5]MC-LR (isomer 2) c | C51H78N10O12 | 1022.5801 | LC-HRMS/MS, thiol | [81] |
| 125 | [D-Leu1,D-Asp3]MC-LR | C51H78N10O12 | 1022.5801 | LC-MS/MS, HRMS/MS | [152,153] |
| 126 | [D-Leu1,Dha7]MC-LR | C51H78N10O12 | 1022.5801 | LC-MS/MS | [75] |
| 127 | [D-Val1]MC-LR | C51H78N10O12 | 1022.5801 | LC-MS/MS | [75] |
| 128 | [Gly1,D-Asp³,Dhb7]MC-RHar | C48H73N13O12 | 1023.5502 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 129 | [D-Asp3]MC-RR | C48H73N13O12 | 1023.5502 | AA, HRMS, NMR | [76,156] |
| 130 | [Dha7]MC-RR | C48H73N13O12 | 1023.5502 | AA, HRMS, MS/MS, NMR | [157] |
| 131 | [D-Asp3,(E)-Dhb7]MC-RR | C48H73N13O12 | 1023.5502 | NMR, HRMS | [158] |
| 132 | [Gly1,D-Asp3]MC-RHar | C48H73N13O12 | 1023.5502 | LC-MS/MS | [92] |
| 133 | [DMAdda5]MC-RR | C48H73N13O12 | 1023.5502 | LC-HRMS/MS, thiol | [82] |
| 134 | MC-WL | C54H72N8O12 | 1024.5270 | LC-MS/MS, thiol | [90] |
| 135 | MC-LW | C54H72N8O12 | 1024.5270 | LC-MS/MS, 15N-enrichment | [140] |
| 136 | [D-Asp3]MC-RCit | C48H72N12O13 | 1024.5342 | LC-HRMS/MS, thiol, 15N-label | [99] |
| 137 | [D-Asp3,ADMAdda5,Thr7]MC-LR | C49H74N10O14 | 1026.5386 | LC-MS/MS, thiol | [159] |
| 138 | [Seco-1/2]MC-HilR b | C50H78N10O13 | 1026.5750 | MS/MS, HRMS | [80] |
| 139 | [D-Asp3,Ser7]MC-RR | C47H73N13O13 | 1027.5451 | LC-MS/MS, thiol | [159] |
| 140 | MC-NfkAbu b | C52H68N8O14 | 1028.4855 | LC-MS/MS, thiol | [90] |
| 141 | MC-M(O)R b | C48H72N10O13S | 1028.5001 | AA, HRMS, NMR | [76] |
| 142 | MC-FR | C52H72N10O12 | 1028.5331 | AA, HRMS, NMR | [76,160] |
| 143 | MC-RF | C52H72N10O12 | 1028.5331 | LC-MS/MS, thiol | [77] |
| 144 | [D-Asp³]MC-HphR | C52H72N10O12 | 1028.5331 | MS/MS | [88] |
| 145 | [Dha7]MC-HphR | C52H72N10O12 | 1028.5331 | AA, HRMS, MS/MS, 1H-NMR | [161] |
| 146 | [D-Asp³]MC-M(O2)R b | C47H70N10O14S | 1030.4794 | LC-MS/MS, thiol, S-oxidation | [93] |
| 147 | [D-Asp3,Dha7]MC-HtyR | C51H70N10O13 | 1030.5124 | AA, HRMS, MS/MS, NMR | [161] |
| 148 | [D-Asp³,DMAdda5]MC-HtyR | C51H70N10O13 | 1030.5124 | MS/MS | [88] |
| 149 | [Dha7]MC-YR | C51H70N10O13 | 1030.5124 | HRMS, MS/MS, AA | [162] |
| 150 | [D-Asp3]MC-RY | C51H70N10O13 | 1030.5124 | HRMS, LC-MS/MS, thiol | [108,163,164] |
| 151 | [Dha7]MC-RY | C51H70N10O13 | 1030.5124 | LC-MS/MS, thiol | [77] |
| 152 | [D-Asp³,Dhb7]MC-RY | C51H70N10O13 | 1030.5124 | LC-MS/MS, thiol | [109] |
| 153 | [D-Asp3]MC-YR | C51H70N10O13 | 1030.5124 | AA, HRMS, MS/MS | [165] |
| 154 | [D-Asp3,(E)-Dhb7]MC-YR | C51H70N10O13 | 1030.5124 | NMR, LC-HRMS/MS | [95] |
| 155 | [DMAdda5]MC-YR | C51H70N10O13 | 1030.5124 | LC–MS/MS, thiol | [131] |
| 156 | MC-LY(OMe) | C53H73N7O14 | 1031.5216 | LC-MS/MS, thiol | [108] |
| 157 | [D-Asp³]MC-(H4)YR | C51H74N10O13 | 1034.5437 | LC–MS/MS, thiol | [131] |
| 158 | [Dha7]MC-(H4)YR | C51H74N10O13 | 1034.5437 | HRMS, NMR, AA | [145] |
| 159 | [DMAdda5]MC-(H4)YR | C51H74N10O13 | 1034.5437 | LC-MS/MS | [137] |
| 160 | MC-YM(O) b | C51H69N7O14S | 1035.4623 | AA, NMR, MS | [166] |
| 161 | [ADMAdda5]MC-HilR | C51H76N10O13 | 1036.5593 | LC-MS/MS | [33] |
| 162 | [ADMAdda5]MC-LHar | C51H76N10O13 | 1036.5593 | HRMS, NMR, AA, MS/MS | [78,79] |
| 163 | MC-AnaR | C52H80N10O12 | 1036.5957 | LC–MS/MS, thiol | [131] |
| 164 | [D-Leu1]MC-LR | C52H80N10O12 | 1036.5957 | NMR, HRMS, MS/MS, AA | [167,168] |
| 165 | [D-Asp³]MC-YY | C54H67N7O14 | 1037.4746 | LC-MS/MS, thiol | [108] |
| 166 | [Gly1,D-Asp3,ADMAdda5,Dhb7]MC-RR | C48H71N13O13 | 1037.5294 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 167 | [Gly1,D-Asp3,ADMAdda5]MC-RR | C49H71N13O13 | 1037.5294 | LC-MS/MS | [92] |
| 168 | MC-RR | C49H75N13O12 | 1037.5658 | NMR, AA, MS | [48] |
| 169 | [(6Z)-Adda5]MC-RR b | C49H75N13O12 | 1037.5658 | NMR, AA, MS | [115,147] |
| 170 | [D-Asp3,D-Glu(OMe)6]MC-RR b | C49H75N13O12 | 1037.5658 | NMR, AA, MS/MS | [169] |
| 171 | [D-Asp3]MC-RHar | C49H75N13O12 | 1037.5658 | LC-HRMS/MS | [170] |
| 172 | [D-Ser1,ADMAdda5]MC-LR | C50H74N10O14 | 1038.5386 | HRMS, MS/MS, AA | [86] |
| 173 | [D-Met1,D-Asp3]MC-LR | C50H76N10O12S | 1040.5365 | LC-MS/MS | [75] |
| 174 | [ADMAdda5,Mser7]MC-LR | C50H76N10O14 | 1040.5542 | HRMS, MS/MS, AA | [86] |
| 175 | [Ser7]MC-RR | C48H75N13O13 | 1041.5607 | AA, HRMS, MS/MS | [84] |
| 176 | [D-Asp³,Mser7]MC-RR | C48H75N13O13 | 1041.5607 | AA, HRMS, MS/MS | [114] |
| 177 | [D-Asp³,Thr7]MC-RR | C48H75N13O13 | 1041.5607 | MS/MS | [171] |
| 178 | [seco-1/6][D-Asp3]MC-RR b | C48H75N13O13 | 1041.5607 | NMR, AA, MS/MS | [169] |
| 179 | MC-HphR | C53H74N10O12 | 1042.5488 | LC-MS/MS, thiol | [34,93] |
| 180 | [D-Glu(OMe)6]MC-FR b | C53H74N10O12 | 1042.5488 | LC-MS/MS | [40] |
| 181 | [D-Leu1]MC-LY | C55H77N7O13 | 1043.5579 | LC-HRMS/MS, LC-MS/MS | [97,172] |
| 182 | MC-M(O2)R b | C48H72N10O14S | 1044.4950 | LC-MS/MS, thiol, S-oxidation | [93] |
| 183 | [D-Asp3]MC-HtyR | C52H72N10O13 | 1044.5280 | AA, MS, NMR | [173] |
| 184 | [Dha7]MC-HtyR | C52H72N10O13 | 1044.5280 | AA, HRMS, MS/MS, 1H-NMR | [161] |
| 185 | [D-Asp3,(E)-Dhb7]MC-HtyR | C52H72N10O13 | 1044.5280 | NMR, AA, HRMS | [142] |
| 186 | [D-Asp3,(Z)-Dhb7]MC-HtyR | C52H72N10O13 | 1044.5280 | NMR, AA, HRMS | [142] |
| 187 | MC-RY | C52H72N10O13 | 1044.5280 | LC-MS/MS, NMR, thiol | [77,163] |
| 188 | MC-YR | C52H72N10O13 | 1044.5280 | AA, NMR, MS | [16] |
| 189 | [Dhb7]MC-YR | C52H72N10O13 | 1044.5280 | LC-HRMS/MS, thiol | [174] |
| 190 | [seco-1/2]MC-FR b | C52H74N10O13 | 1046.5437 | HRMS, MS/MS | [80,154] |
| 191 | MC-(H2)YR | C52H74N10O13 | 1046.5437 | LC-HRMS/MS, thiol | [82] |
| 192 | MC-HphHph | C57H73N7O12 | 1047.5317 | LC-MS/MS | [75] |
| 193 | [D-Asp³,Ser7]MC-HtyR | C51H72N10O14 | 1048.5229 | LC-MS/MS | [33] |
| 194 | [D-Asp³,Mser7]MC-RY | C51H72N10O14 | 1048.5229 | LC-MS/MS, thiol | [108] |
| 195 | [Ser7]MC-YR | C51H72N10O14 | 1048.5229 | LC–MS/MS | [102] |
| 196 | MC-(H4)YR | C52H76N10O13 | 1048.5593 | HRMS, MS/MS, NMR | [80] |
| 197 | [ADMAdda5]MC-HilHar | C52H78N10O13 | 1050.5750 | LC-MS/MS | [33] |
| 198 | [D-Leu1, D-Asp³,ADMAdda5]MC-LR | C52H78N10O13 | 1050.5750 | LC-MS/MS | [153] |
| 199 | [D-Leu1,Adda(O)5]MC-LR b | C52H78N10O13 | 1050.5750 | LC-MS/MS, thiol | [81] |
| 200 | [D-Leu1]MC-HilR | C53H82N10O12 | 1050.6114 | LC-MS/MS | [75] |
| 201 | [D-Leu1]MC-LHar | C53H82N10O12 | 1050.6114 | LC-MS/MS | [75] |
| 202 | [D-Leu1,Glu(OMe)6]MC-LR b | C53H82N10O12 | 1050.6114 | HRMS/MS | [152] |
| 203 | [Hil1]MC-LR | C53H82N10O12 | 1050.6114 | LC-MS/MS, thiol | [81] |
| 204 | [D-Asp³,(E)-Dhb7]MC-HtyY | C55H69N7O14 | 1051.4902 | AA, NMR, HRMS | [175] |
| 205 | MC-YY | C55H69N7O14 | 1051.4902 | NMR, LC-HRMS/MS | [95] |
| 206 | [Gly1,D-Asp3,ADMAdda5]MC-RHar | C49H73N13O13 | 1051.5451 | LC-MS/MS | [92] |
| 207 | [Gly1,D-Asp3,ADMAdda5,Dhb7]MC-RHar | C49H73N13O13 | 1051.5451 | LC-MS/MS, HRMS, AA, thiol | [91] |
| 208 | [D-Asp³,ADMAdda5]MC-RR | C49H73N13O13 | 1051.5451 | LC-MS/MS | [153] |
| 209 | [D-Asp3,ADMAdda5,Dhb7]MC-RR | C49H73N13O13 | 1051.5451 | NMR, HRMS, AA, LC-MS/MS, thiol | [150,176] |
| 210 | [D-Glu(OC3H6O)6]MC-LR b | C52H80N10O13 | 1052.5906 | AA, HRMS, NMR | [76] |
| 211 | [D-Leu1,Adda(OH)5]MC-LR b | C52H80N10O13 | 1052.5906 | LC-MS/MS, thiol | [81] |
| 212 | [D-Asp3]MC-WR | C53H71N11O12 | 1053.5284 | AA, NMR, MS | [87] |
| 213 | [Dha7]MC-WR | C53H71N11O12 | 1053.5284 | LC–MS/MS, thiol | [131] |
| 214 | [DMAdda5]MC-WR | C53H71N11O12 | 1053.5284 | LC–MS/MS, thiol | [131] |
| 215 | [D-Asp3]MC-RW | C53H71N11O12 | 1053.5284 | LC-HRMS/MS, thiol, 15N-label | [99] |
| 216 | [D-Met1]MC-LR | C51H78N10O12S | 1054.5521 | LC-MS/MS | [75] |
| 217 | [D-Leu1]MC-MR | C51H78N10O12S | 1054.5521 | LC-HRMS/MS, thiol, S-oxidation | [81] |
| 218 | [D-Leu1,Mser7]MC-LR | C52H82N10O13 | 1054.6063 | HRMS/MS | [152] |
| 219 | [Mser7]MC-RR | C49H77N13O13 | 1055.5764 | LC-HRMS/MS, thiol | [82] |
| 220 | [ADMAdda5]MC-FR | C53H72N10O13 | 1056.5280 | LC-MS/MS | [137] |
| 221 | [D-Asp³,ADMAdda5]MC-HphR | C53H72N10O13 | 1056.5280 | LC-MS/MS | [153] |
| 222 | MC-HtyR | C53H74N10O13 | 1058.5437 | AA, MS, NMR | [173] |
| 223 | [D-Asp³,D-Glu(OMe)6]MC-HtyR b | C53H74N10O13 | 1058.5437 | MS/MS | [88] |
| 224 | [D-Glu(OMe)6]MC-YR b | C53H74N10O13 | 1058.5437 | NMR, LC-HRMS/MS | [95] |
| 225 | [D-Ser1,D-Asp³]MC-HtyR | C52H72N10O14 | 1060.5229 | LC-MS/MS, thiol | [93] |
| 226 | [D-Asp³]MC-Y(OMe)R | C52H72N10O14 | 1060.5229 | LC–MS/MS, thiol | [131] |
| 227 | [DMAdda5]MC-Y(OMe)R | C52H72N10O14 | 1060.5229 | LC–MS/MS, thiol | [131] |
| 228 | [seco-4/5][D-Asp³]MC-HtyR b | C52H74N10O14 | 1062.5386 | LC-MS/MS, thiol | [93] |
| 229 | [Ser7]MC-HtyR | C52H74N10O14 | 1062.5386 | AA, HRMS, MS/MS, NMR | [161] |
| 230 | [D-Asp³,Mser7]MC-HtyR | C52H74N10O14 | 1062.5386 | MALDI-TOF MS | [88] |
| 231 | [D-Asp³,ADMAdda5]MC-(H4)YR | C52H74N10O14 | 1062.5386 | LC-MS/MS | [153] |
| 232 | [Mser7]MC-RY | C52H74N10O14 | 1062.5386 | LC-MS/MS, thiol | [77] |
| 233 | [D-Asp³,Mser7]MC-YHar | C52H74N10O14 | 1062.5386 | NMR, LC-HRMS/MS | [95] |
| 234 | [Mser7]MC-YR | C52H74N10O14 | 1062.5386 | LC–MS/MS, NMR, thiol, HRMS | [131] |
| 235 | MC-HphHty | C57H73N7O13 | 1063.5266 | LC-MS/MS | [75] |
| 236 | [D-Leu1,ADMAdda5]MC-LR | C53H80N10O13 | 1064.5906 | LC –MS/MS | [153] |
| 237 | [D-Leu1,Glu(OMe)6]MC-HilR b | C54H84N10O12 | 1064.6270 | HRMS/MS | [152] |
| 238 | [D-Asp³,(E)-Dhb7]MC-HtyHty | C56H71N7O14 | 1065.5059 | AA, NMR, HRMS | [175] |
| 239 | [ADMAdda5]MC-RR | C50H75N13O13 | 1065.5607 | LC-MS/MS, LC-HRMS/MS | [104,153] |
| 240 | MC-HarHar | C51H79N13O12 | 1065.5971 | LC–MS/MS | [177] |
| 241 | [D-Leu1,D-Asp3]MC-RR | C51H79N13O12 | 1065.5971 | LC-MS/MS | [75] |
| 242 | MC-WR | C54H73N11O12 | 1067.5440 | AA, HRMS, NMR | [76] |
| 243 | [D-Leu1,D-Glu(OMe)6,Mser7]MC-LR b | C53H84N10O13 | 1068.6219 | HRMS/MS | [152] |
| 244 | [D-Met(O)1]MC-LR b | C51H78N10O13S | 1070.5471 | HRMS/MS | [152] |
| 245 | [D-Leu1]MC-M(O)R b | C51H78N10O13S | 1070.5471 | LC-HRMS/MS, thiol, S-oxidation | [81,97] |
| 246 | [D-Leu1,D-Asp³]MC-HphR | C55H78N10O12 | 1070.5801 | LC-MS/MS | [153] |
| 247 | [D-Phe1]MC-LR | C55H78N10O12 | 1070.5801 | LC-MS/MS | [75] |
| 248 | [D-Leu1]MC-FR | C55H78N10O12 | 1070.5801 | LC-MS/MS, thiol | [81] |
| 249 | MC-KynR b | C53H73N11O13 | 1071.5389 | LC-MS/MS, thiol | [90] |
| 250 | [D-Asp3,ADMAdda5]MC-HtyR | C53H72N10O14 | 1072.5229 | AA, HRMS, MS/MS | [178] |
| 251 | [D-Asp3,ADMAdda5,Dhb7]MC-HtyR | C53H72N10O14 | 1072.5229 | NMR, HRMS, AA | [150] |
| 252 | [ADMAdda5]MC-YR | C53H72N10O14 | 1072.5229 | LC-MS/MS | [33] |
| 253 | MC-MhtyR | C54H76N10O13 | 1072.5593 | LC-HRMS/MS, thiol, esterification | [96] |
| 254 | [D-Asp3,(E)-Dhb7]MC-HtyW | C57H70N8O13 | 1074.5062 | NMR, LC-HRMS/MS | [95] |
| 255 | MC-RY(OMe) | C53H74N10O14 | 1074.5386 | LC-MS/MS, thiol | [77] |
| 256 | [D-Asp³]MC-Hty(OMe)R | C53H74N10O14 | 1074.5386 | LC-MS/MS, thiol | [93] |
| 257 | MC-Y(OMe)R | C53H74N10O14 | 1074.5386 | LC–MS/MS, thiol | [131] |
| 258 | [seco-4/5]MC-HtyR b | C53H76N10O14 | 1076.5542 | LC-MS/MS, thiol | [93] |
| 259 | [ADMAdda5]MC-(H4)YR | C53H76N10O14 | 1076.5542 | LC-MS/MS | [137] |
| 260 | [Mser7]MC-HtyR | C53H76N10O14 | 1076.5542 | LC-MS/MS, thiol | [93] |
| 261 | [D-Leu1,ADMAdda5]MC-LHar | C54H82N10O13 | 1078.6063 | LC-MS/MS | [153] |
| 262 | [ADMAdda5]MC-RHar | C51H77N13O13 | 1079.5764 | LC-MS/MS | [153] |
| 263 | [D-Leu1]MC-RR | C52H81N13O12 | 1079.6128 | LC-MS/MS | [75] |
| 264 | MC-OiaR b | C54H73N11O13 | 1083.5389 | LC-MS/MS, thiol | [90] |
| 265 | [D-Met(O)1,Glu(OMe)6]MC-LR b | C52H80N10O13S | 1084.5627 | HRMS/MS | [152] |
| 266 | [D-Leu1]MC-HphR | C56H80N10O12 | 1084.5957 | LC-MS/MS | [75] |
| 267 | [D-Leu1]MC-M(O2)R b | C51H78N10O14S | 1086.5420 | LC-HRMS/MS, thiol, S-oxidation | [81] |
| 268 | [ADMAdda5]MC-(H4)YHar | C54H78N10O14 | 1090.5699 | LC-MS/MS | [137] |
| 269 | MC-NfkR b | C54H73N11O14 | 1099.5338 | LC-MS/MS, thiol | [90] |
| 270 | [D-Leu1]MC-HtyR | C56H80N10O13 | 1100.5906 | LC-MS/MS | [153] |
| 271 | [D-Leu1,Ser7]MC-HtyR | C55H80N10O14 | 1104.5855 | LC-HRMS/MS | [170] |
| 272 | MC-LR Cys conjugate b | C52H81N11O14S | 1115.5685 | HRMS, MS/MS, semisynthesis | [80] |
| 273 | [D-Leu1]MC-LR Cys conjugate b | C55H87N11O14S | 1157.6155 | LC-HRMS/MS, semisynthesis | [81] |
| 274 | [D-Leu1]MC-LR Cys sulfoxide conjugate b | C55H87N11O15S | 1173.6104 | LC-HRMS/MS, semisynthesis, S-oxidation | [81] |
| 275 | [D-Leu1]MC-LR γ-GluCys conjugate b | C60H94N12O17S | 1286.6581 | LC-HRMS/MS, semisynthesis | [81] |
| 276 | [D-Leu1]MC-LR γ-GluCys sulfoxide conjugate b | C60H94N12O18S | 1302.6530 | LC-HRMS/MS, semisynthesis, S-oxidation | [81] |
| 277 | [D-Asp³]MC-RRGSH conjugate b | C58H90N16O18S | 1330.6340 | LC-HRMS/MS, thiol, 15N-label, semisynthesis | [99] |
| 278 | [D-Leu1]MC-LR GSH conjugate b | C62H97N13O18S | 1343.6795 | LC-HRMS/MS, semisynthesis | [81] |
| 279 | [D-Leu1]MC-LR GSH sulfoxide conjugate b | C62H97N13O19S | 1359.6744 | LC-HRMS/MS, semisynthesis, S-oxidation | [81] |
| Entry a | Microcystin | LD50 | Reference |
|---|---|---|---|
| 7 | MC-LA | 50 | [226] |
| 18 | MC-AR | 250 | [76] |
| 22 | MC-YA | 60–70 | [227] |
| 25 | [D-Asp3,Dha7]MC-LR | 160–300 | [83] |
| 39 | [D-Asp3]MC-LR | 200–500 | [139,215] |
| 40 | [D-Asp3,(E)-Dhb7]MC-LR | 70 | [142] |
| 42 | [Dha7]MC-LR | 250 | [83] |
| 43 | [DMAdda5]MC-LR | 90–100 | [76,149] |
| 66 | MC-LR | 50 | [205] |
| 68 | [(6Z)-Adda5]MC-LR | >1200 | [115,147] |
| 83 | MC-LY | 90 | [206] |
| 89 | [D-Asp3,ADMAdda5]MC-LR | 160 | [78,79] |
| 92 | MC-HilR | 100 | [80] |
| 94 | [D-Glu(OMe)6]MC-LR | >1000 | [85,140,189] |
| 105 | [Mser7]MC-LR | 150 | [80] |
| 111 | [D-Asp3]MC-FR | 90 ± 10 | [216] |
| 121 | [ADMAdda5]MC-LR | 60 | [78,79] |
| 129 | [D-Asp3]MC-RR | 350 ± 10 | [216] |
| 130 | [Dha7]MC-RR | 180 | [157] |
| 131 | [D-Asp3,(E)-Dhb7]MC-RR | 250 | [142] |
| 141 | MC-M(O)R | 700–800 | [76] |
| 142 | MC-FR | 250 | [76] |
| 160 | MC-YM(O) | 56 | [166] |
| 162 | [ADMAdda5]MC-LHar | 60 | [78,79] |
| 164 | [D-Leu1]MC-LR | 100 | [167,168] |
| 168 | MC-RR | 500–800 | [48,76] |
| 169 | [(6Z)-Adda5]MC-RR | >1200 | [115,147] |
| 183 | [D-Asp3]MC-HtyR | 80–100 | [147] |
| 185 | [D-Asp3,(E)-Dhb7]MC-HtyR | 70 | [142] |
| 188 | MC-YR | 70 | [16] |
| 209 | [D-Asp3,ADMAdda5,Dhb7]MC-RR | 200 | [150] |
| 210 | [D-Glu(OC3H6O)6]MC-LR | >1000 | [76] |
| 212 | [D-Asp3]MC-WR | 95 ± 10 | [216] |
| 222 | MC-HtyR | 160–300 | [147] |
| 242 | MC-WR | 150–200 | [76] |
| 251 | [D-Asp3, ADMAdda5,Dhb7]MC-HtyR | 100 | [150] |
| 272 | MC-LR Cys conjugate | 1000 | [80] |
| Entry a | Microcystin | Type of PP | Origin of PP | IC50 | Reference |
|---|---|---|---|---|---|
| 7 | MC-LA | PP1 | Rabbit muscle | 2.3 | [237] |
| PP2A | Human hepatocytes | 0.56 | [238] | ||
| PP2A | Rabbit muscle | 0.05 | [237] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.161 ± 0.002 | [239] | ||
| 14 | MC-LV | PP1 | Rabbit skeletal muscle | 0.06–0.45 | [133] |
| 17 | MC-LL | PP1 | Rabbit skeletal muscle | 0.06–0.45 | [133] |
| 25 | [D-Asp3,Dha7] MC-LR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.254 ± 0.004 | [239] |
| 34 | MC-LM | PP1 | Rabbit skeletal muscle | 0.06–0.45 | [133] |
| 39 | [D-Asp3]MC-LR | PP2A | Rabbit skeletal muscle | 0.09 | [215] |
| 40 | [D-Asp3,(E)-Dhb7]MC-LR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.201 ± 0.003 | [239] |
| 41 | [D-Asp3,(Z)-Dhb7]MC-LR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.16 ± 0.01 | [239] |
| 42 | [Dha7]MC-LR | PP1 | Rabbit skeletal muscle | 0.54–5 | [240,241] |
| PP2A | Bovine kidney | 0.11 ± 0.04 | [241] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.167 ± 0.003 | [239] | ||
| 43 | [DMAdda5]MC-LR | rPP1c | Recombinant rabbit skeletal muscle PP1 | 1.5 | [240] |
| 52 | MC-LF | rPP1c | Recombinant rabbit skeletal muscle PP1 | 1.8 | [224] |
| PP1 | Rabbit skeletal muscle | 0.06–0.45 | [133] | ||
| PP2A | Human hepatocytes | 0.57 | [238] | ||
| PP2A | Human red blood cells | 1.1 | [224] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.10 ± 0.02 | [239] | ||
| 66 | MC-LR | PP1 | Rabbit muscle | 0.1–1.9 | [215,217,237,240,241,242,243,244,245] |
| PP1 | Chicken gizzard myosin B | 6 | [246] | ||
| PP1 | Liver of grass carp | 0.90 | [247] | ||
| rPP1c | Recombinant rabbit skeletal muscle PP1 | 1.2 | [224] | ||
| PP2A | Rabbit skeletal muscle | 0.04–0.5 | [215,217,237,243,248] | ||
| PP2A | Human hepatocytes | 0.46 | [238] | ||
| PP2A | Human erythrocytes | 0.03–2.2 | [241,242,249,250] | ||
| PP2A | Bovine heart | 0.05-2 | [241,246,251] | ||
| PP2A | Bovine kidney | 0.2 ± 0.1 | [241] | ||
| PP2A | Mouse brain | 0.28–3.15 | [218,247,252,253] | ||
| PP2A | Liver of grass carp | 0.28 | [247] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.032 ± 0.004 | [239] | ||
| 68 | [(6Z)-Adda5]MC-LR | PP2A | Mouse brain | 80 | [252] |
| rPP1c | Recombinant rabbit skeletal muscle PP1 | >100 | [240] | ||
| 83 | MC-LY | PP2A | Human hepatocytes | 0.34 | [238] |
| 89 | [D-Asp3,ADMAdda5]MC-LR | PP2Ab | 4 | [178] | |
| 94 | [D-Glu(OMe)6]MC-LR | rPP1c | Recombinant rabbit skeletal muscle PP1 | >100 | [240] |
| 97 | [D-Asp3,Dha7]MC-RR | rPP1c | Recombinant rabbit skeletal muscle PP1 | 0.22 ± 0.01 | [239] |
| 129 | [D-Asp3]MC-RR | PP2A rPP2Ac | Human red blood cells Recombinant human PP2A catalytic subunit | 0.45–11.5 0.30 ± 0.01 | [254] [239] |
| 130 | [Dha7]MC-RR | PP1 | Rabbit muscle | 8.3 ± 0.8 | [241] |
| PP2A | Bovine kidney | 4 ± 1 | [241] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.29 ± 0.01 | [239] | ||
| PP1 | Rabbit skeletal muscle | 2.6–5.7 | [254] | ||
| 131 | [D-Asp3,(E)-Dhb7]MC-RR | PP1 | Rabbit skeletal muscle | 1.8–56.4 | [215,254] |
| PP2A | Rabbit skeletal muscle | 2.4 | [215] | ||
| PP2A | Human red blood cells | 17.9–49.4 | [254] | ||
| 135 | MC-LW | rPP1c | Recombinant rabbit skeletal muscle PP1 | 1.9 | [224] |
| PP2A | Human hepatocytes | 0.29 | [238] | ||
| PP2A | Human red blood cells | 1.1 | [224] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.114 ± 0.003 | [239] | ||
| 142 | MC-FR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.069 ± 0.003 | [239] |
| 149 | [Dha7]MC-YR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.379 ± 0.003 | [239] |
| 164 | [D-Leu1]MC-LR | PP1 | Recombinant rabbit skeletal muscle PP1 | 0.5–4.43 | [167,168] |
| 168 | MC-RR | PP1 | Rabbit skeletal muscle | 0.68 | [215] |
| PP1 | Chicken gizzard myosin B | 3 | [246] | ||
| PP1 | Liver of grass carp | 3.60 | [247] | ||
| rPP1c | Recombinant rabbit skeletal muscle PP1 | 1.5 | [224] | ||
| PP2A | Human red blood cells | 0.241–175 | [241,249,250] | ||
| PP2A | Human hepatocytes | 0.60 | [238] | ||
| PP2A | Mouse brain | 0.72–1.4 | [218,224,247,252] | ||
| PP2A | Bovine cardiac muscle | 1 | [246] | ||
| PP2A | Bovine Kidney | 10 ± 2 | [241] | ||
| PP2A | Rabbit skeletal muscle | 0.1 | [215] | ||
| PP2A | Liver of grass carp | 0.64 | [247] | ||
| rPP2Ac | Recombinant human PP2A catalytic subunit | 0.056 ± 0.002 | [239] | ||
| 169 | [(6Z)-Adda5]MC-RR | rPP2Ac | Recombinant human PP2A catalytic subunit | 10.1 ± 0.3 | [239] |
| PP2A | Mouse brain | 80 | [252] | ||
| 183 | [D-Asp3]MC-HtyR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.098 ± 0.006 | [239] |
| 185 | [D-Asp3,(E)-Dhb7]MC-HtyR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.122 ± 0.005 | [239] |
| 186 | [D-Asp3,(Z)-Dhb7]MC-HtyR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.110 ± 0.008 | [239] |
| 188 | MC-YR | PP1 PP1 PP2A PP2A PP2A PP2A PP2A PP2A rPP2Ac | Rabbit skeletal muscle Liver of grass carp Human red blood cells Human hepatocytes Bovine kidney Rabbit skeletal muscle Mouse brain Liver of grass carp Recombinant human PP2A catalytic subunit | 1.0 0.90 0.26–9.0 0.84 0.09 ± 0.02 0.26 0.39–1.3 0.40 0.125 ± 0.005 | [215] [247] [241,249,250] [238] [241] [215] [218,247] [247] [239] |
| 242 | MC-WR | rPP2Ac | Recombinant human PP2A catalytic subunit | 0.18 ± 0.01 | [239] |
| 251 | [D-Asp3,ADMAdda5,Dhb7]MC-HtyR | PP1 | Rabbit skeletal muscle | 0.15–0.24 | [254] |
| PP2A | Human red blood cells | 0.06 | [254] | ||
| PP2A | Bovine heart | 0.06 | [251] | ||
| PP4 | Porcine testis | 0.04 | [251] | ||
| PP5 | Recombinant human PP5 expressed in E. coli | 0.5 | [251] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins 2019, 11, 714. https://doi.org/10.3390/toxins11120714
Bouaïcha N, Miles CO, Beach DG, Labidi Z, Djabri A, Benayache NY, Nguyen-Quang T. Structural Diversity, Characterization and Toxicology of Microcystins. Toxins. 2019; 11(12):714. https://doi.org/10.3390/toxins11120714
Chicago/Turabian StyleBouaïcha, Noureddine, Christopher O. Miles, Daniel G. Beach, Zineb Labidi, Amina Djabri, Naila Yasmine Benayache, and Tri Nguyen-Quang. 2019. "Structural Diversity, Characterization and Toxicology of Microcystins" Toxins 11, no. 12: 714. https://doi.org/10.3390/toxins11120714
APA StyleBouaïcha, N., Miles, C. O., Beach, D. G., Labidi, Z., Djabri, A., Benayache, N. Y., & Nguyen-Quang, T. (2019). Structural Diversity, Characterization and Toxicology of Microcystins. Toxins, 11(12), 714. https://doi.org/10.3390/toxins11120714