Tetrodotoxins Occurrence in Non-Traditional Vectors of the North Atlantic Waters (Portuguese Maritime Territory, and Morocco Coast)

 , ,
, ,  and
and
Abstract
:1. Introduction
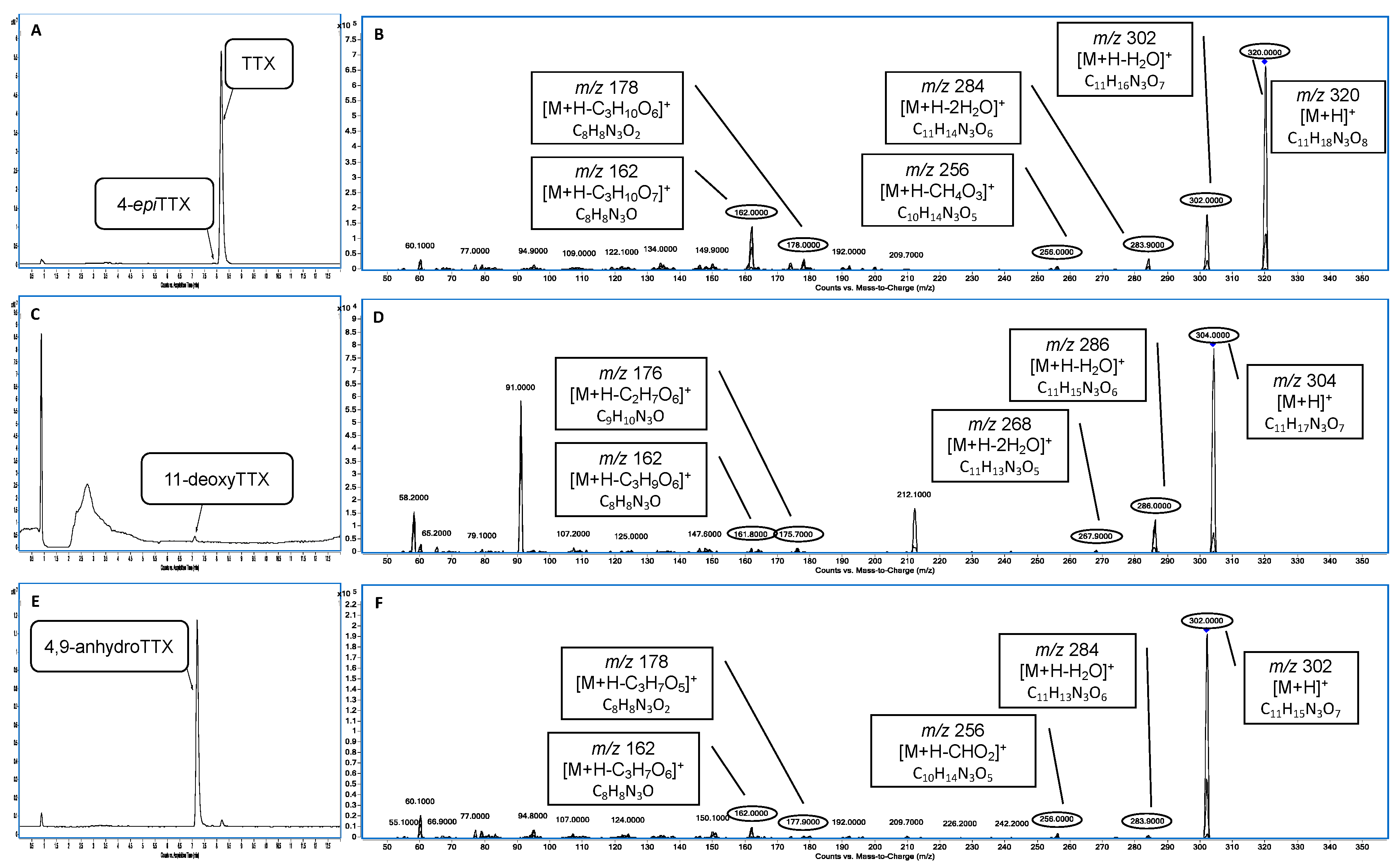
2. Results and Discussion
2.1. Portuguese Continental Coast
2.2. Madeira Island (Madeira Archipelago)
2.3. São Miguel Island (Azores Archipelago)
2.4. Moroccan Coast
2.5. Statistical Analysis
3. Conclusions
4. Materials and Methods
4.1. Sampled Points and Selected Species
4.2. Reagents
4.3. Sample Extraction
4.4. UHPLC Conditions
4.5. MS Methods
4.5.1. Source Conditions
4.5.2. UHPLC-MS/MS Analysis: MRM Mode
4.5.3. UHPLC-MS/MS Analysis: PIS Mode
4.5.4. UHPLC-HRMS
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, Toxicity, Source, Distribution and Detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef] [Green Version]
- Botana, L.M.; Alfonso, A.; Rodriguez, I.; Botana, A.M.; Louzao, M.C.; Vieytes, M.R. How safe is safe for marine toxins monitoring? Toxins 2016, 8, 208. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Pratheepa, V.K.; Botana, L.M.; Vasconcelos, V. Emergent toxins in North Atlantic temperate waters: A challenge for monitoring programs and legislation. Toxins 2015, 7, 859–885. [Google Scholar] [CrossRef] [PubMed]
- Tahara, Y.; Hirata, Y. Studies on the puffer fish toxin. J. Pharm. Soc. Jpn. 1909, 29, 587–625. [Google Scholar]
- Miyazawa, K.; Noguchi, T. Distribution and origin of tetrodotoxin. J. Toxicol. Toxin Rev. 2001, 20, 11–33. [Google Scholar] [CrossRef]
- Pratheepa, V.; Vasconcelos, V. Microbial diversity associated with tetrodotoxin production in marine organisms. Environ. Toxicol. Pharmacol. 2013, 36, 1046–1054. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-Producing Bacteria: Detection, Distribution and Migration of the Toxin in Aquatic Systems. Toxins 2017, 9, 166. [Google Scholar] [CrossRef]
- Rodriguez, I.; Alfonso, A.; Alonso, E.; Rubiolo, J.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.; Margassery, L.; Dobson, A.; et al. The association of bacterial C 9-based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci. Rep. 2017, 7, 40880. [Google Scholar] [CrossRef]
- Choudhary, G.; Yotsu-Yamashita, M.; Shang, L.; Yasumoto, T.; Dudley, S.C., Jr. Interactions of the C-11 hydroxyl of tetrodotoxin with the sodium channel outer vestibule. Biophys. J. 2003, 84, 287–294. [Google Scholar] [CrossRef]
- Scheib, H.; McLay, I.; Guex, N.; Clare, J.; Blaney, F.; Dale, T.; Tate, S.; Robertson, G. Modeling the pore structure of voltage-gated sodium channels in closed, open, and fast-inactivated conformation reveals details of site 1 toxin and local anesthetic binding. J. Mol. Model. 2006, 12, 813–822. [Google Scholar] [CrossRef]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of specific modifications of several hydroxyls of tetrodotoxin on its affinity to rat brain membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar] [PubMed]
- Bentur, Y.; Ashkar, J.; Lurie, Y.; Levy, Y.; Azzam, Z.S.; Litmanovich, M.; Golik, M.; Gurevych, B.; Golani, D.; Eisenman, A. Lessepsian migration and tetrodotoxin poisoning due to Lagocephalus sceleratus in the eastern Mediterranean. Toxicon 2008, 52, 964–968. [Google Scholar] [CrossRef]
- Katikou, P.; Georgantelis, D.; Sinouris, N.; Petsi, A.; Fotaras, T. First report on toxicity assessment of the Lessepsian migrant pufferfish Lagocephalus sceleratus (Gmelin, 1789) from European waters (Aegean Sea, Greece). Toxicon 2009, 54, 50–55. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First Toxicity Report of Tetrodotoxin and 5,6,11-TrideoxyTTX in the Trumpet Shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Azevedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New Gastropod Vectors and Tetrodotoxin Potential Expansion in Temperate Waters of the Atlantic Ocean. Marine Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Higgins, C.; Higman, W.; Hungerford, J. Potential threats posed by tetrodotoxins in UK waters: Examination of detection methodology used in their control. Mar. Drugs 2015, 13, 7357–7376. [Google Scholar] [CrossRef] [PubMed]
- Regulation (EC) No. 853/2004 of the European Parliament and of the Council of 29 April 2004 Laying down Specific Hygiene Rules for Food of Animal Origin OJ L 139, 30.4.2004. Available online: http://eur-lex.europa.eu (accessed on 14 December 2010).
- Regulation (EC) No 854/2004 of the European Parliament and of the Council of 29 April 2004 Laying down Specific Rules for the Organisation of Official Controls on Products of Animal Origin Intended for Human Consumption. OJ L 139, 30.4.2004 (18 June 2015). pp. 206–320. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32007R1246 (accessed on 14 December 2010).
- Gerssen, A.; Bovee, T.; Klijnstra, M.; Poelman, M.; Portier, L.; Hoogenboom, R. First report on the occurrence of tetrodotoxins in bivalve mollusks in the Netherlands. Toxins 2018, 10, 450. [Google Scholar] [CrossRef]
- Knutsen, H.K.; Alexander, J.; Barregard, L.; Bignami, M.; Bruschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 4752. [Google Scholar]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in Greek Shellfish by UPLC–MS/MS potentially linked to the presence of the Dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef]
- Shoji, Y.; Yotsu-Yamashita, M.; Miyazawa, T.; Yasumoto, T. Electrospray ionization mass spectrometry of tetrodotoxin and its analogs: Liquid chromatography/mass spectrometry, tandem mass spectrometry, and liquid chromatography/tandem mass spectrometry. Anal. Biochem. 2001, 290, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First Identification of 5,11-Dideoxytetrodotoxin in Marine Animals, and Characterization of Major Fragment Ions of Tetrodotoxin and Its Analogs by High Resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, P.; Alfonso, A.; Otero, P.; Katikou, P.; Georgantelis, D.; Botana, L.M. Liquid chromatography–mass spectrometry method to detect Tetrodotoxin and Its analogues in the puffer fish Lagocephalus sceleratus (Gmelin, 1789) from European waters. Food Chem. 2012, 132, 1103–1111. [Google Scholar] [CrossRef]
- Bane, V.; Brosnan, B.; Barnes, P.; Lehane, M.; Furey, A. High-resolution mass spectrometry analysis of tetrodotoxin (TTX) and its analogues in puffer fish and shellfish. Food Addit. Contam. Part A Chem. Anal. Control Expos. Risk Assess. 2016, 33, 1468–1489. [Google Scholar] [CrossRef]
- Knox, G.A. Hard Shores. In The Ecology of Seashores; Kennish, M.J., Ed.; CRC Press: Boca Raton, FL, USA, 2001; pp. 20–86. [Google Scholar]
- Crothers, J.H. Common topshells: An introduction to the biology of Osilinus lineatus with notes on other species in the genus. Field Stud. 2001, 10, 115–160. [Google Scholar]
- Forster, G.R. The ecology of Echinus esculentus L. Quantitative distribution and rate of feeding. J. Mar. Biol. Assoc. UK 1959, 38, 361–367. [Google Scholar] [CrossRef]
- Lin, S.J.; Hwang, D.F. Possible source of tetrodotoxin in the starfish Astropecten scoparius. Toxicon 2001, 39, 573–579. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Mebs, D. Occurrence of 11-oxotetrodotoxin in the red-spotted newt, Notophthalmus viridescens, and further studies on the levels of tetrodotoxin and its analogues in the newt’s efts. Toxicon 2003, 41, 893–897. [Google Scholar] [CrossRef]
- Ferguson, J.C. Feeding activity in echinaster and its induction with dissolved nutrients. Biol. Bull. 1969, 136, 374–384. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.L.; Harmelin, J.G.; Almeida, A.J. Structure of fish assemblages on coastal rocky shores of the Azores. Boletim do Museu Municipal do Funchal 2001, 6, 127–138. [Google Scholar]
- Noguchi, T.; Ebesu, J.S.M. Puffer poisoning: Epidemiology and treatment. J. Toxicol. Toxin Rev. 2001, 20, 1–10. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M.; Abbott, J.P.; Flewelling, L.J.; Richardson, R.W.; Dickey, R.W.; Jester, E.L.E.; et al. Saxitoxin Puffer Fish Poisoning in the United States, with the First Report of Pyrodinium bahamense as the Putative Toxin Source. Environ. Health Perspect. 2006, 114, 1502–1507. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, J.; Noguchi, T.; Narita, H.; Nara, M.; Jeon, J.K.; Otsuka, M.; Hashimoto, K. Occurrence of Tetrodotoxin in a Starfish, Astropecten scoparius. Agric. Biol. Chem. 1985, 10, 3069–3070. [Google Scholar]
- Miyazawa, K.; Noguchi, T.; Maruyama, J.; Jeon, J.K.; Otsuka, M.; Hashimoto, K. Occurrence of tetrodotoxin in the starfishes Astropecten polyacanthus and A. scoparius in the Seto Inland Sea. Mar. Biol. 1985, 90, 61–64. [Google Scholar] [CrossRef]
- Sugita, H.; Iwata, J.; Miyajima, C.; Kubo, T.; Noguchi, T.; Hashimoto, K.; Deguchi, Y. Changes in microflora of a puffer fish Fugu niphobles, with different water temperatures. Mar. Biol. 1989, 101, 299–304. [Google Scholar] [CrossRef]
- Llewellyn, L.E. Revisiting the association between sea surface temperature and the epidemiology of fish poisoning in the south Pacific: Reassessing the link between ciguatera and climate change. Toxicon 2010, 56, 691–697. [Google Scholar] [CrossRef]
- Ashton, M.; Tosteson, T.; Tosteson, C. The effect of elevated temperature on the toxicity of the laboratory cultured dinoflagellate Ostreopsis lenticularis (Dinophyceae). Revista de Biología Tropical 2003, 51, 1–6. [Google Scholar]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtza, J.; Bean, B.P.; Baker-Austin, C.; et al. New Invasive Nemertean Species (Cephalothrix simula) in England with High Levels of Tetrodotoxin and a Microbiome Linked to Toxin Metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef]
- Dayrat, B. Review of the current knowledge of the systematics of onchidiidae (mollusca: Gastropoda: Pulmonata) with a checklist of nominal species. Zootaxa 2009, 2068, 1–26. [Google Scholar]
- Carefoot, T.H. Aplysia: Its biology and ecology. Oceanogr. Mar. Biol. Annu. Rev. 1987, 25, 167–284. [Google Scholar]
- Ito, K.; Okabe, S.; Asakawa, M.; Bessho, K.; Taniyama, S.; Shida, Y.; Ohtsuka, S. Detection of tetrodotoxin (TTX) from two copepods infecting the grass puffer Takifugu niphobles: TTX attracting the parasites? Toxicon 2006, 48, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Jen, H.C.; Lin, S.J.; Tsai, Y.H.; Chen, C.H.; Lin, Z.C.; Hwang, D.F. Tetrodotoxin poisoning evidenced by solid-phase extraction combining with liquid chromatography-tandem mass spectrometry. J. Chromatogr. B: Anal. Technol. Biomed. Life Sci. 2008, 871, 95–100. [Google Scholar] [CrossRef]
- Tsai, Y.H.; Ho, P.H.; Hwang, C.C.; Hwang, P.A.; Cheng, C.A.; Hwang, D.F. Tetrodotoxin in several species of xanthid crabs in southern Taiwan. Food Chem. 2006, 95, 205–212. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Structure | Analogue | R1 | R2 | R3 | R4 | Molecular Formula | Exact Mass |
|---|---|---|---|---|---|---|---|
 | TTX | H | OH | OH | CH2OH | C11H17N3O8 | 320.1088 |
| 4-epiTTX | OH | H | OH | CH2OH | C11H17N3O8 | 320.1088 | |
| 6-epiTTX | H | OH | CH2OH | OH | C11H17N3O8 | 320.1088 | |
| 11-deoxyTTX | H | OH | OH | CH3 | C11H17N3O7 | 304.1139 | |
| 11-norTTX-6(R/S)-ol | H | OH | H | OH | C10H15N3O7 | 290.0983 | |
| 6,11-dideoxyTTX | H | OH | H | CH3 | C11H17N3O6 | 288.1190 | |
| 11-oxoTTX | H | OH | OH | CHO | C11H17N3O9 | 336.1038 | |
 | 5-deoxyTTX | H | OH | OH | CH2OH | C11H17N3O7 | 304.1139 |
| 5,11-dideoxyTTX | H | OH | H | CH3 | C11H17N3O6 | 288.1190 | |
| 5,6,11-trideoxyTTX | H | OH | H | CH3 | C11H17N3O5 | 272.1214 | |
 | 4,9-anhydroTTX | --- | --- | OH | CH2OH | C11H15N3O7 | 302.0983 |
| 6-epi-4,9-anhydroTTX | --- | --- | CH2OH | OH | C11H15N3O7 | 302.0983 | |
 | Tetrodonic acid | --- | --- | --- | --- | C11H17N3O8 | 320.1088 |
| Suspicious TTXs | Multiple Reaction Monitoring (MRM) | Product Ions Scan (PIS) | |||
|---|---|---|---|---|---|
| Precursor Ion | Product Ions | [M+H]+ | [M+H-H2O]+ | Other Product Ions | |
| oxoTTX | 336 | 318/162 | 336 | 318 | 300/176/162 |
| TTX and epimers | 320 | 302/162 | 320 | 302 | 284/256/178/162 |
| deoxyTTX | 304 | 286/162 | 304 | 286 | 268/176/162 |
| anhydroTTX | 302 | 256/162 | 302 | 284 | 256/178/162 |
| norTTX | 290 | 272/162 | 290 | 272 | 176/162 |
| dideoxyTTX | 288 | 270/224 | 288 | 270 | 224/178/162 |
| trideoxyTTX | 272 | 254/162 | 272 | 254 | 236/178/162 |
| anhydrotrideoxyTTX | 254 | 236/162 | 254 | 236 | 178/162 |
| Sampling Date | Sampling Site | Sample | Species | Trophic Level [Ref] | Edible | Code | Suspicious TTXs | RT (min) | Elemental Composition | Theoretical m/z | Experimental m/z | Error (mDa) | TTX eq (µg/Kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| September 2011 | Porto Côvo | Limpet | Patella depressa | Grazer [27] | Yes | 225 | 11-oxoTTX | 7.2 | C11H17N3O9 | 336.1038 | 336.1000 | −3.8 | <LOQ |
| Gastropod | Phorcus lineatus | Grazer [28] | Yes | 226 | trideoxyTTX | 7.7 | C11H17N3O5 | 272.1241 | 272.1232 | −0.9 | <LOQ | ||
| October 2011 | Pedra de Jacó | Sea-urchin | Echinus esculentus | Grazer [29] | Yes | 229_2 | 11-oxoTTX | 7.2 | C11H17N3O9 | 336.1038 | 336.1020 | −1.8 | <LOQ |
| Ovar | Gastropod | Charonia lampas | 3rd Level Predator [30] | Yes | 232_2 | trideoxyTTX | 7.7 | C11H17N3O5 | 272.1241 | 272.1230 | −1.1 | <LOQ | |
| 232_3 | trideoxyTTX | 7.7 | C11H17N3O5 | 272.1241 | 272.1251 | 1.0 | <LOQ | ||||||
| November 2011 | Memória | Gastropod | Nucella lapillus | 1st Level Predator [27] | Yes | 236 | trideoxyTTX | 7.7 | C11H17N3O5 | 272.1241 | 272.1239 | −0.2 | <LOQ |
| Phorcus lineatus | Grazer [28] | Yes | 239 | 11-oxoTTX | 7.2 | C11H17N3O9 | 336.1038 | 336.1000 | −3.8 | <LOQ | |||
| Gibbula umbilicalis | Grazer [28] | Yes | 240 | trideoxyTTX | 7.7 | C11H17N3O5 | 272.1241 | 272.1238 | −1.1 | <LOQ |
| Sampling Date | Sampling Site | Sample | Species | Trophic Level [Ref] | Edible | Code | Standard TTX | Suspicious TTXs | RT (min) | Elemental Composition | Theoretical m/z | Experimental m/z | Error (mDa) | TTX eq (µg/Kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| June 2013 | Lagoa | Starfish | Ophidiaster ophidianus | Detritivorous [32] | No | 412 | norTTX | 3.1 | C10H15N3O7 | 290.0983 | 290.2489 | 150.6 | 30,902 | |
| norTTX | 4.6 | C10H15N3O7 | 290.0983 | 290.2303 | 132.0 | 699 | ||||||||
| June 2013 | Ilhéu de São Roque | Starfish | Ophidiaster ophidianus | Detritivorous [32] | No | 424 | norTTX | 3.1 | C10H15N3O7 | 290.0983 | 290.2475 | 149.2 | 42,604 | |
| norTTX | 4.6 | C10H15N3O7 | 290.0983 | 290.2298 | 131.5 | 1638 | ||||||||
| June 2013 | Caloura | Starfish | Ophidiaster ophidianus | Detritivorous [32] | No | 435 | norTTX | 3.1 | C10H15N3O7 | 290.0983 | 290.2478 | 149.9 | 20,853 | |
| June 2013 | Caloura | Fish (Gonads) | Sphoeroides marmoratus | 3rd Level Predator [33] | Yes | 438 G | TTX | 8.1 | C11H17N3O8 | 320.1088 | 320.1088 | 0.0 | 352,886 | |
| 4-epiTTX | 7.8 | C11H17N3O8 | 320.1088 | 320.1082 | −0.6 | 3157 | ||||||||
| deoxyTTX | 7.7 | C11H17N3O7 | 304.1139 | 304.1129 | −1.0 | 446 | ||||||||
| 11-deoxyTTX | 7.0 | C11H17N3O7 | 304.1139 | 304.1126 | −1.3 | 157,156 | ||||||||
| deoxyTTX | 6.5 | C11H17N3O7 | 304.1139 | 304.1124 | −1.5 | 108,375 | ||||||||
| 4,9-anhydroTTX | 7.1 | C11H15N3O7 | 302.0983 | 302.1005 | 2.2 | 91 | ||||||||
| dideoxyTTX | 6.4 | C11H17N3O6 | 288.1190 | 288.1157 | −3.3 | 22,232 | ||||||||
| trideoxyTTX | 5.0 | C11H17N3O5 | 272.1241 | 272.1198 | −4.3 | 12,708 | ||||||||
| trideoxyTTX | 4.8 | C11H17N3O5 | 272.1241 | 272.1228 | −1.3 | 37,367 | ||||||||
| trideoxyTTX | 4.6 | C11H17N3O5 | 272.1241 | 272.1227 | −1.4 | 1849 | ||||||||
| anhydrotrideoxyTTX | 4.7 | C11H15N3O4 | 254.1135 | 254.1122 | −1.3 | 364 | ||||||||
| June 2013 | Caloura | Fish (Liver) | Sphoeroides marmoratus | 3rd Level Predator [33] | Yes | 438 F | TTX | 8.1 | C11H17N3O8 | 320.1088 | 320.1098 | 1.0 | 41 | |
| 4-epiTTX | 7.8 | C11H17N3O8 | 320.1088 | nd | 6 | |||||||||
| 11-deoxyTTX | 7.0 | C11H17N3O7 | 304.1139 | nd | <LOQ | |||||||||
| deoxyTTX | 6.9 | C11H17N3O7 | 304.1139 | nd | 4 | |||||||||
| deoxyTTX | 6.5 | C11H17N3O7 | 304.1139 | 304.1153 | 1.4 | 136 | ||||||||
| 4,9-anhydroTTX | 7.1 | C11H15N3O7 | 302.0983 | nd | <LOQ | |||||||||
| dideoxyTTX | 6.4 | C11H17N3O6 | 288.1190 | nd | 283 | |||||||||
| trideoxyTTX | 5.0 | C11H17N3O5 | 272.1241 | nd | 31 | |||||||||
| trideoxyTTX | 4.8 | C11H17N3O5 | 272.1241 | 272.1263 | 2.2 | 250 | ||||||||
| trideoxyTTX | 4.6 | C11H17N3O5 | 272.1241 | nd | 14 | |||||||||
| June 2013 | Caloura | Fish (Muscle) | Sphoeroides marmoratus | 3rd Level Predator [33] | Yes | 438 M | TTX | 8.1 | C11H17N3O8 | 320.1088 | 320.1088 | 0.0 | 9918 | |
| 4-epiTTX | 7.8 | C11H17N3O8 | 320.1088 | 320.1067 | −2.1 | 118 | ||||||||
| deoxyTTX | 7.7 | C11H17N3O7 | 304.1139 | 304.1164 | 2.5 | 29 | ||||||||
| 11-deoxyTTX | 7.0 | C11H17N3O7 | 304.1139 | 304.1172 | 3.3 | 48 | ||||||||
| deoxyTTX | 6.9 | C11H17N3O7 | 304.1139 | 304.1123 | −1.6 | 65 | ||||||||
| deoxyTTX | 6.5 | C11H17N3O7 | 304.1139 | 304.1100 | −3.9 | 2085 | ||||||||
| 4,9-anhydroTTX | 7.1 | C11H15N3O7 | 302.0983 | 302.0958 | −2.5 | 45 | ||||||||
| dideoxyTTX | 6.4 | C11H17N3O6 | 288.1190 | 288.1145 | −4.5 | 2953 | ||||||||
| trideoxyTTX | 5.0 | C11H17N3O5 | 272.1241 | nd | 122 | |||||||||
| trideoxyTTX | 4.8 | C11H17N3O5 | 272.1241 | 272.1217 | −2.4 | 507 | ||||||||
| trideoxyTTX | 4.6 | C11H17N3O5 | 272.1241 | nd | 26 | |||||||||
| anhydrotrideoxyTTX | 4.7 | C11H15N3O4 | 254.1135 | 254.1129 | −0.6 | 5 |
| Sampling Date | Sampling Site | Sample | Species | Trophic Level [Ref] | Edible | Code | Standard TTX | Suspicious TTXs | RT (min) | Elemental Composition | Theoretical m/z | Experimental m/z | Error (mDa) | TTX eq (µg/Kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| July 2013 | Mrizika | Gastropod | Onchidella celtica | Grazer [43] | No | 474 | TTX | 8.1 | C11H17N3O8 | 320.1088 | 320.1082 | −0.6 | 12 | |
| 4-epiTTX | 7.8 | C11H17N3O8 | 320.1088 | 320.1071 | 1.7 | 8 | ||||||||
| TTX epimer | 7.5 | C11H17N3O8 | 320.1088 | nd | 4 | |||||||||
| July 2013 | Oualidia | Gastropod | Aplysia depilans | Grazer [44] | No | 476 | TTX | 8.1 | C11H17N3O8 | 320.1088 | nd | <LOQ | ||
| 4-epiTTX | 7.8 | C11H17N3O8 | 320.1088 | nd | <LOQ |
| Sampling Sites | Coefficents ± SE | z | p |
|---|---|---|---|
| São Miguel | 8.96 ± 0.001 | 5000.3 | <0.001 |
| Madeira | −19.26 ± 21.38 | −0.9 | 0.37 |
| Morocco | −10.67 ± 0.38 | −28.2 | <0.001 |
| Continental Portugal | −19.26 ± 13.30 | −1.4 | 0.15 |
| Date | Location | Sampling Site | Geographic Coordinates |
|---|---|---|---|
| September 2011 | Continental Portugal | Luz | 37°04′37″ N; 8°44′51″ W |
| Camilo | 37°05′14″ N; 8°40′06″ W | ||
| Porto Côvo | 37°53′33″ N; 8°47′38″ W | ||
| Pelo Negro | 41°19′22″ N; 8°71′69″ W | ||
| October 2011 | Continental Portugal | Pedra de Jacó | 41°17′14″ N; 8°70′78″ W |
| Ovar | 40°52′58″ N; 8°40′31″ W | ||
| Esposende | 41°29′5″ N; 8°46′45″ W | ||
| November 2011 | Continental Portugal | Memória | 41°13′50″ N; 8°43′18″ W |
| December 2011 | Continental Portugal | Esposende | 41°29′5″ N; 8°46′45″ W |
| Valadares | 41°5′29″ N; 8°39′27″ W | ||
| January 2012 | Continental Portugal | Póvoa do Varzim | 41°22′41″ N; 8°46′7″ W |
| Angeiras | 41°15′50″ N; 8°43′37″ W | ||
| September 2012 | Madeira Island | Reis Magos | 32°39′16″ N; 16°49′05″ W |
| Caniçal | 32°44′20″ N; 16°44′17″ W | ||
| June 2013 | São Miguel Island | Cruzeiro | 37° 50′31″ N; 25° 41′33″ W |
| Étar de Ponta Delgada | 37°44′19″ N; 25°39′38″ W | ||
| São Roque | 37°45′15″ N; 25°38′31″ W | ||
| Mosteiros | 37°53′25″ N; 25°49′14″ W | ||
| Lagoa | 37°44′42″ N; 25°19′47″ W | ||
| Caloura | 37°42′49″ N; 25°29′54″ W | ||
| July 2013 | Morocco Coast | Casablanca corniche | 33°36′01″ N; 7°39′57″ W |
| El Jadida Haras | 33°14′42″ N; 8°28′37″ W | ||
| El Jadida Sâada | 33°14′42″ N; 8°32′26″ W | ||
| Sidi Bouzid | 33°13′57″ N; 8°33′20″ W | ||
| Mrizika | 32°57′21″ N; 8°46′53″ W | ||
| Oualidia | 32°43′55″ N; 9°02′57″ W |
| Compound | Precursor Ion | Product Ion | CE |
|---|---|---|---|
| 11-oxo-TTX | 336 | 318 | 24 |
| 178 | 40 | ||
| TTX 4-epiTTX TTX epimers Tetrodonic acid | 320 | ||
| 302 | 24 | ||
| 161.9 | 36 | ||
| 11-deoxyTTX deoxyTTX | 304 | 286 | 24 |
| 176 | 40 | ||
| 4,9-anhydro-TTX anhydroTTX | 302 | 256 | 28 |
| 161.9 | 40 | ||
| 11-norTTX-6(R/S)-ol | 290 | 272 | 24 |
| 162 | 40 | ||
| dideoxyTTX | 288 | 270 | 24 |
| 224 | 40 | ||
| hydroxytrideoxyTTX | 288 | 270 | 24 |
| 162 | 40 | ||
| trideoxyTTX | 272 | 254 | 24 |
| 162 | 40 | ||
| hydroxyanhydrotrideoxyTTX | 270 | 252 | 24 |
| 162 | 40 | ||
| anhydrotrideoxyTTX | 254 | 236 | 24 |
| 162 | 40 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.; Rodríguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. Tetrodotoxins Occurrence in Non-Traditional Vectors of the North Atlantic Waters (Portuguese Maritime Territory, and Morocco Coast). Toxins 2019, 11, 306. https://doi.org/10.3390/toxins11060306
Silva M, Rodríguez I, Barreiro A, Kaufmann M, Neto AI, Hassouani M, Sabour B, Alfonso A, Botana LM, Vasconcelos V. Tetrodotoxins Occurrence in Non-Traditional Vectors of the North Atlantic Waters (Portuguese Maritime Territory, and Morocco Coast). Toxins. 2019; 11(6):306. https://doi.org/10.3390/toxins11060306
Chicago/Turabian StyleSilva, Marisa, Inés Rodríguez, Aldo Barreiro, Manfred Kaufmann, Ana Isabel Neto, Meryem Hassouani, Brahim Sabour, Amparo Alfonso, Luis M. Botana, and Vitor Vasconcelos. 2019. "Tetrodotoxins Occurrence in Non-Traditional Vectors of the North Atlantic Waters (Portuguese Maritime Territory, and Morocco Coast)" Toxins 11, no. 6: 306. https://doi.org/10.3390/toxins11060306