Towards Managing and Controlling Aflatoxin Producers Within Aspergillus Species in Infested Rice Grains Collected from Local Markets in Kenya



Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
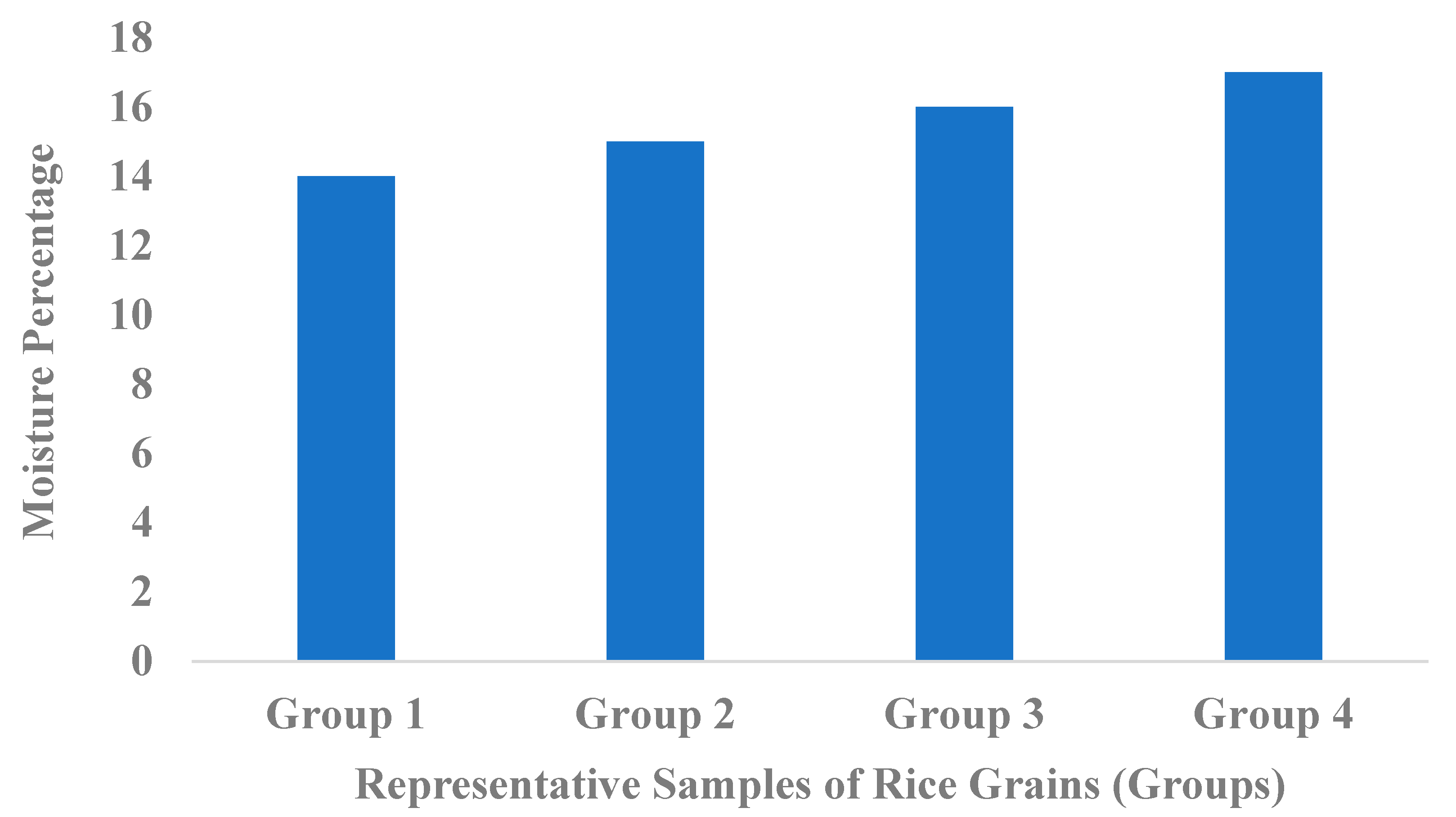
2.1.1. Moisture Content
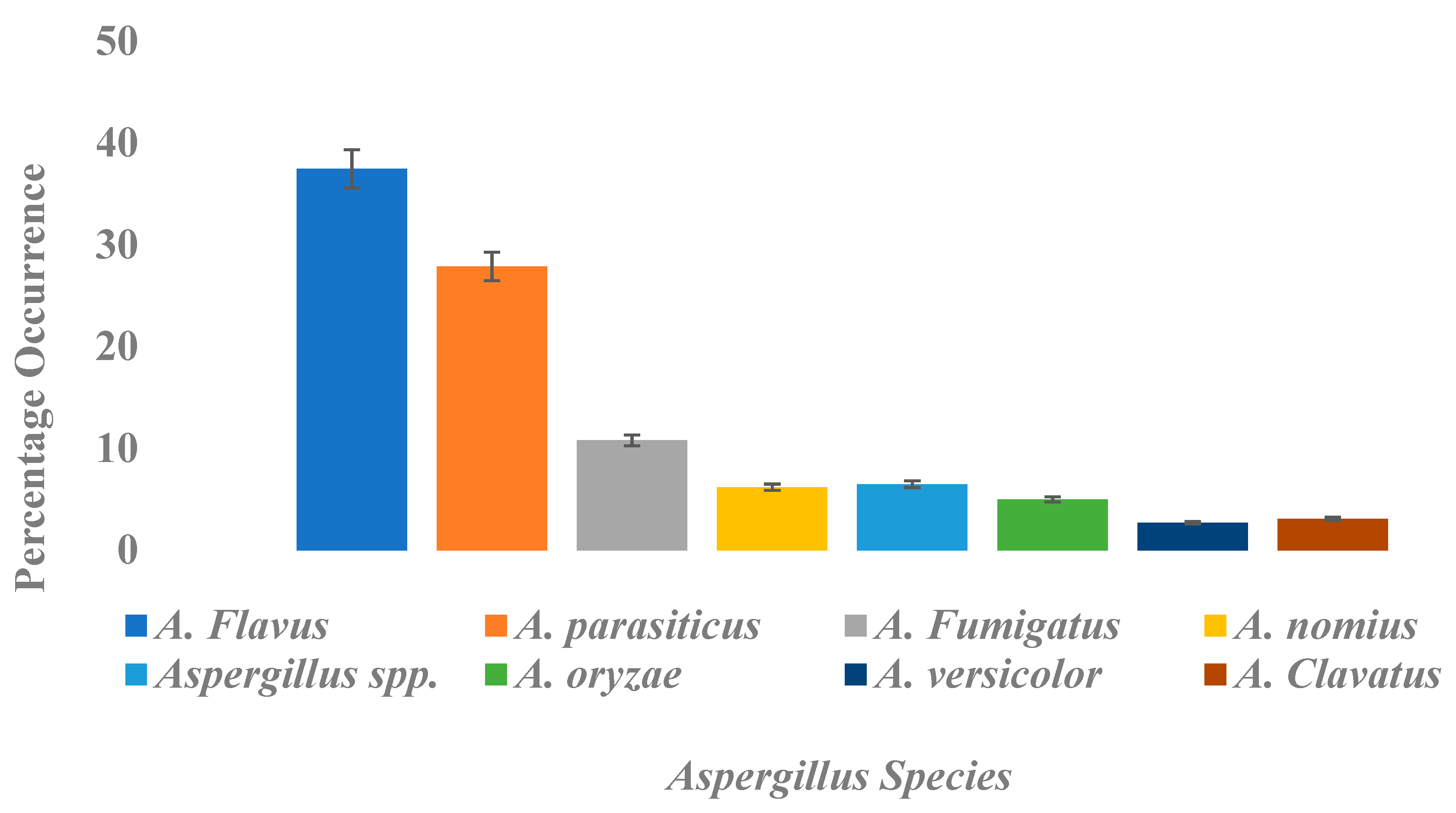
2.1.2. Isolation of Aspergillus Species
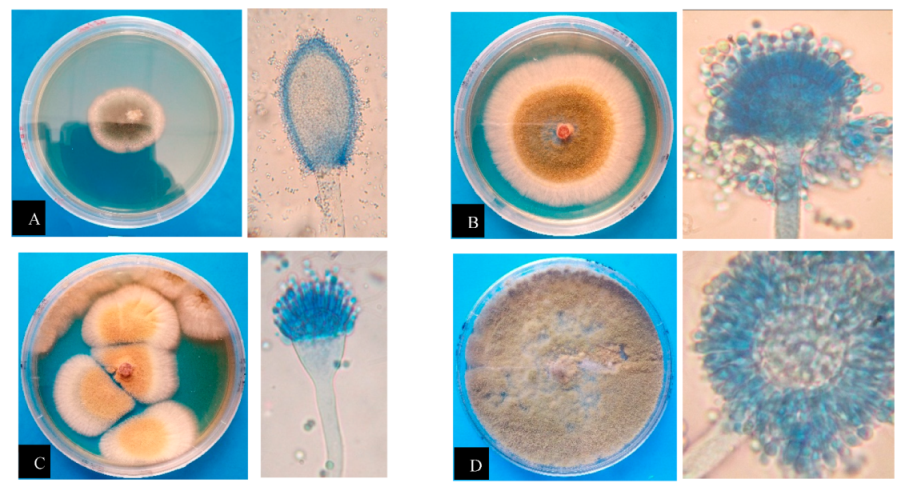
2.1.3. Morphological Analysis
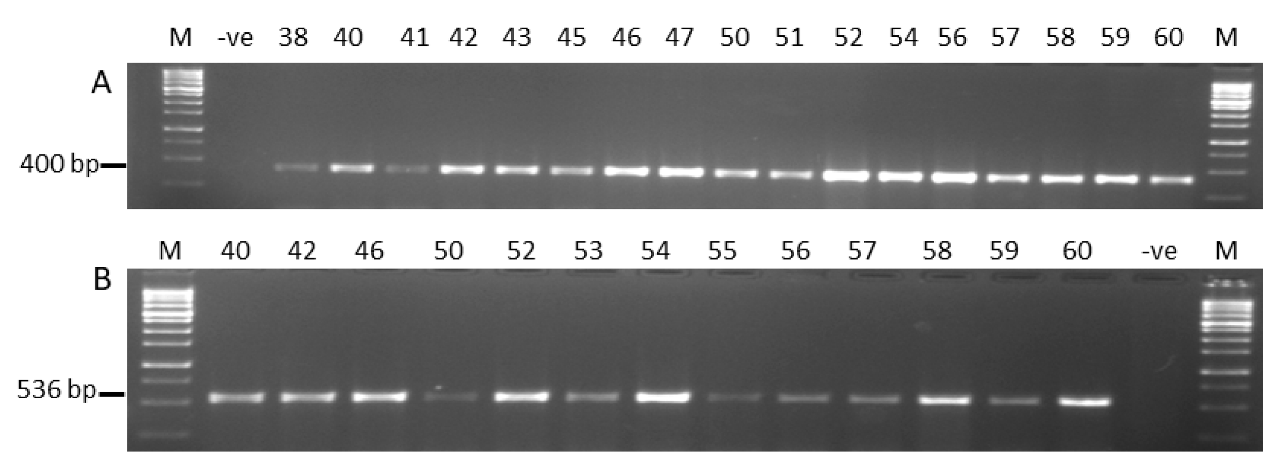
2.1.4. Molecular Characterization
2.2. Discussion
3. Conclusions
4. Materials and Methods
4.1. Samples
4.2. Moisture Content Analysis
4.3. Isolation and Enumeration of Fungal Species
4.4. Phenotypic Characterization
4.5. Molecular Characterization of the Isolates
4.6. Diagnostic PCR Using AspergillusUniversal Primers
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Haitham, S.A.; Ali, A.B.; Kamel, A.A.; Khalid, S.E.; Abdallah, M.E.; Alsum, B.A. Detection of Aspergillus and Penicillium species producing aflatoxin in rice grains imported into Saudi Arabia. Wulfenia 2013, 20, 313–323. [Google Scholar]
- Reiter, E.; Vouk, F.; Bohm, J.; Razzazi-Fazeli, E. Aflatoxins in rice a limited survey of products marketed in Austria. Food Control 2010, 21, 988–991. [Google Scholar] [CrossRef]
- Lai, X.; He, Z.; Liu, R.; Liu, C. Potential for aflatoxin B1 and B2 production by Aspergillus flavus strains isolated from rice samples. Saudi J. Biol. Sci. 2015, 22, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Sohaib, A.; Ali, S.W.; Ahmed, A.; Mahmood, R. Molecular Characterization of Fungal Species Isolated from Rice Grains. Institute of Agricultural Sciences; University of the Punjab, Quid-i-Azam Campus: Lahore, Pakistan, 2019. [Google Scholar]
- Schmidt, M.; Zannini, E.; Arendt, E.K. Recent Advances in Physical Post-Harvest Treatments for Shelf-Life Extension of Cereal Crops. Foods 2018, 7, 45. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef]
- Lereau, M.; Gouas, D.; Villar, S.; Besaratinia, A.; Hautefeuille, A.; Berthillon, P.; Martel-Planche, G.; da Costa, A.N.; Ortiz-Cuaran, S.; Hantz, O.; et al. Interactions between hepatitis B virus and aflatoxin B1 Effects on p53 induction in Hepa RG cells. J. Gen. Virol. 2012, 93, 640–650. [Google Scholar] [CrossRef] [PubMed]
- IARC Monograph Aflatoxins. IARC Monograph Aflatoxins. IARC monographs on the evaluation of carcinogenic risks to Humans. In Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene; IARC Press: Lyon, France, 2002; Volume 82, pp. 171–300. [Google Scholar]
- Gemeda, N.; Woldeamanuel, Y.; Asrat, D.; Debela, A.; Lemma, H.; Belete, Y. Assessment of Aflatoxigeinic AspergillusSpecies in Food Commodities from Local Market of Addis Ababa. Research 2014, 1. [Google Scholar] [CrossRef]
- Lee, J.; Her, J.Y.; Lee, K.G. Reduction of aflatoxins (B1, B2, G1, and G2) in soybean-based model systems. Food Chem. 2015, 189, 45–51. [Google Scholar] [CrossRef]
- Ruadrew, S.; Craft, J.; Aidoo, K. Occurrence of toxigenic Aspergillus spp. and aflatoxins in selected food commodities of Asian origin sourced in the West of Scotland. Food Chem. Toxicol. 2013, 55, 653–658. [Google Scholar] [CrossRef]
- Naseer, R.; Sultana, B.; Khan, M.; Naseer, D.; Nigam, P. Utilization of waste fruit-peels to inhibit aflatoxins synthesis by Aspergillus flavus: A biotreatment of rice for safer storage. Bioresour. Technol. 2014, 172, 423–428. [Google Scholar] [CrossRef]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: Bioscience; Environment & Agriculture; Environmental Studies & Management; Food Science & Technology. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar]
- Raper, K.B.; Fennell, D.I. The Genus Aspergillus; The Williams & Wilkins Company: Philadelphia, PA, USA, 1965; pp. 370–376. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetic. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Abbas, K.H. Aflatoxins and Food Safety; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Magan, N.; Hope, R.; Cairns, V.; Aldred, D. Post-harvest fungal ecology: Impact of fungal growth and mycotoxin accumulation in stored grain. Eur. J. Plant Pathol. 2003, 109, 723–730. [Google Scholar] [CrossRef]
- Shanakht, H.; Shahid, A.A.; Ali, S.W. characterization of fungal microbiota on rice grains from local markets of Lahore. J. Hyg. Eng. Des. 2014, 37, 35–40. [Google Scholar]
- Renu, K.; Agarwal, M.K.; Bhagayavant, S.S.; Verma, P.; Nagar, D.P. Detection of Aspergillus flavus using PCR method from fungus infested food grains collected from local market. Ann. Plant Sci. 2018, 7, 2073–2077. [Google Scholar]
- Ibrahim, F.; Jalal, H.; Khan, A.B.; Asghar, M.A.; Iqbal, J.; Ahmed, A.; Nadeem, G. Prevalence of Aflatoxigenic Aspergillus in Food and Feed Samples from Karachi. J. Infect. Mol. Biol. Preval. 2016, 4, 1–8. [Google Scholar] [CrossRef]
- Reddy, K.R.N.; Reddy, C.S.; Mangala, U.N.; Muralidharan, K. Site of Infection of Aspergillus sp. in seeds of rice cultivars. J. Mycol. 2006, 36, 271–277. [Google Scholar]
- Aydin, A.; Aksu, H.; Gunsen, U. Mycotoxin levels and incidence of mould in Turkish rice. Environ. Monit. Assess. 2010, 10, 1661–1688. [Google Scholar] [CrossRef]
- Majid, Z.; Maryam, E. Molecular variation analysis of Aspergillus flavus using polymerase chain reaction-restriction fragment length polymorphism of the internal transcribed spacer rDNA region. Exp. Ther. Med. 2016, 12, 1628–1632. [Google Scholar]
- Ehrlich, K.C. Non-aflatoxigenic Aspergillus flavus to prevent aflatoxin contamination in crops: advantages and limitations. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Hadi, A.A.; Carter, D.; Magan, N. Discrimination between aflatoxigenic and non-aflatoxigenic Aspergillus section Flavi strains from Egyptian peanuts using molecular and analytical techniques. World Mycotoxin J. 2011, 4, 69–77. [Google Scholar] [CrossRef]
- Rodrigues, P.; Soares, C.; Kozakiewicz, Z.; Paterson, R.R.M.; Lima, N.; Venâncio, A. Identification and characterization of Aspergillus flavus and aflatoxins. In Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Méndez-Villas, A., Ed.; Formatex: Badajoz, Spain, 2007; pp. 527–534. [Google Scholar]
- Rashid, M.; Khalil, S.; Ayub, N.; Ahmed, W.; Khan, A.G. Categorization of Aspergillus flavus and Aspergillus parasiticus isolates of stored wheat grains into aflatoxinogenics and non-aflatoxinogenics. Pak. J. Bot. 2009, 40, 2177–2192. [Google Scholar]
- Degola, F.; Berni, E.; Dall’Asta, C.; Spotti, E.; Marchelli, R.; Ferrero, I.; Restivo, F. A multiplex RT-PCR Approach to Detect Aflatoxigenic Strains of Aspergillus flavus. J. Appl. Microbiol. 2007, 103, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Houshyarfard, M.; Rouhani, H.; Falahati-Rastegar, M.; Malekzadeh-Shafaroudi, S.; Mehdikhani Moghaddam, E.; Chang, P. Gene Deletion Patterns in non-aflatoxigenic Strains of Aspergillus flavus. Mycol. Iran. 2014, 1, 87–97. [Google Scholar]
- Al-Shuhaib, M.B.S.; Albakri, A.H.; Alwan, S.H.; Almandil, N.B.; AbdulAzeez, S.; Borgio, J.F. Optimal pcr primers for rapid and accurate detection of Aspergillus flavus isolates. Microb. Pathog. 2018, 116, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Shekhany, K.A.M.; Rostam, S.R.K. Detection of Aflatoxigenic Aspergillus flavus in Maize Grains and Soils in Sulaimani Province using Molecular Approaches. J. Zankoy Sulaimani 2016, 18, 25–36. [Google Scholar] [CrossRef]
- Hussain, A.; Afzal, A.; Irfan, M.; Malik, K.A. Molecular Detection of Aflatoxin Producing Strains of Aspergillus flavus from Peanut (Arachis Hypogaea). Turk. J. Agric. Food Sci. Technol. 2015, 3, 335–341. [Google Scholar] [CrossRef]
- Commission Regulation (EC). Laying down the Methods of Sampling and Analysis for the Official Control of the Levels of Mycotoxins in Foodstuffs; Official Journal of the European Union: Brussels, Belgium, 2006. [Google Scholar]
- Botton, B.; Breton, A.; Fèvre, M.; Gauthier, S.; Guy, P.; Larpent, J.P.; Reymond, P.; Sanglier, J.J.; Vayssier, Y.; et Veau, P. Moisissures Utiles et Nuisibles, Importance Industrielle; Masson: Paris, France, 1990; p. 349. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage; Blackie Academic and Professional: Cambridge, UK, 1997. [Google Scholar]
- Atlas, R.M. Handbook of Microbiological Media; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Varga, J.; Frisvad, J.C.; Samson, R.A. Two new aflatoxin producing species and an overview of Aspergillus section Flavi. Stud. Mycol. 2011, 69, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.A.; Matur, S.B.; Neergaard, P. Detection of seed borne fungi in sorghum and location of Fusarium moniliforme in seed. Seed Sci. Technol. 1975, 3, 683–690. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Set | Primer Name | Sequences (5′–3′) | Length of PCR Product (bp) |
|---|---|---|---|
| 1 | ITS1 | F-TCCGTAGGTGAACCTGCGG | 598 |
| ITS4 | R-TCCTCCGCTTATTGATATGC | ||
| 2 | aflD | F-ACCGCTACGCCGGCACTCTCGGCAC | 400 |
| R-GTTGGCCGCCAGCTTCGACACTCCG | |||
| 3 | aflM | F-GCCGCAGGCCGCGGAGAAAGTGGT | 536 |
| R-GGGGATATACTCCCGCGACACAGCC |
| Set | PCR Fragment | Initial Denaturation | Denaturation | Annealing | Elongation | Final Elongation | Number of Cycles | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | ITS | 94 °C | 5 min | 95 °C | 60 s | 52 °C | 60 s | 72 °C | 60 s | 72 °C | 10 min | 35 |
| 2 | aflD | 94 °C | 5 min | 94 °C | 60 s | 64 °C | 60 s | 72 °C | 60 s | 72 °C | 10 min | 33 |
| 3 | aflM | 95 °C | 5 min | 95 °C | 60 s | 65 °C | 60 s | 72 °C | 2 min | 72 °C | 10 min | 33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Douksouna, Y.; Masanga, J.; Nyerere, A.; Runo, S.; Ambang, Z. Towards Managing and Controlling Aflatoxin Producers Within Aspergillus Species in Infested Rice Grains Collected from Local Markets in Kenya. Toxins 2019, 11, 544. https://doi.org/10.3390/toxins11090544
Douksouna Y, Masanga J, Nyerere A, Runo S, Ambang Z. Towards Managing and Controlling Aflatoxin Producers Within Aspergillus Species in Infested Rice Grains Collected from Local Markets in Kenya. Toxins. 2019; 11(9):544. https://doi.org/10.3390/toxins11090544
Chicago/Turabian StyleDouksouna, Youmma, Joel Masanga, Andrew Nyerere, Steven Runo, and Zachée Ambang. 2019. "Towards Managing and Controlling Aflatoxin Producers Within Aspergillus Species in Infested Rice Grains Collected from Local Markets in Kenya" Toxins 11, no. 9: 544. https://doi.org/10.3390/toxins11090544
APA StyleDouksouna, Y., Masanga, J., Nyerere, A., Runo, S., & Ambang, Z. (2019). Towards Managing and Controlling Aflatoxin Producers Within Aspergillus Species in Infested Rice Grains Collected from Local Markets in Kenya. Toxins, 11(9), 544. https://doi.org/10.3390/toxins11090544