Maternal Exposure Results in Long-Term Deoxynivalenol Persistence in Piglets’ Plasma and Modulates the Immune System


Abstract
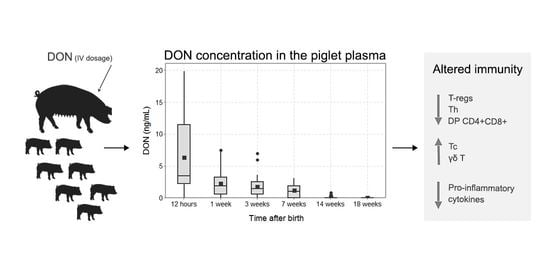
:
1. Introduction
2. Results
2.1. Deoxynivalenol Concentration in the Plasma of the Piglets after Intrauterine Exposure
2.2. Deoxynivalenol Concentration in the Plasma of the Piglets after Intraperitoneal Exposure at 35 Days of Life
2.3. Plasma Enzymes Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT) Determination
2.4. Hematological Parameters
2.5. Total Immunoglobulin Levels in the Plasma of the Piglets
2.6. Lymphocyte Subpopulations in the Blood of the Piglets after Intrauterine DON Exposure
2.7. The Expression of Cytokine mRNA after Non-Specific Stimulation of Blood Leukocytes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals and Blood Sampling
5.2. Liquid Chromatography in Combination with High-Resolution Mass Spectrometry (LC–MS/MS (HR))
5.3. Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT) Determination in the Plasma
5.4. Hematological Parameters
5.5. Serum Immunoglobulin Concentration
5.6. Flow Cytometry Analysis of the Lymphocyte Subpopulations in the Blood
5.7. Non-Specific Stimulation of Blood Leukocytes and Cytokine Gene Expression Determination by Quantitative RT-PCR Analysis
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- European Commission. COMMISSION RECOMMENDATION 2006/576/EC of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- Pestka, J.J.; Smolinski, A.T. Deoxynivalenol: Toxicology and potential effects on humans. J. Toxicol. Environ. Heal.-Part B Crit. Rev. 2005, 8, 39–69. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Lauwers, M.; Croubels, S.; Letor, B.; Gougoulias, C.; Devreese, M. Biomarkers for exposure as a tool for efficacy testing of a mycotoxin detoxifier in broiler chickens and pigs. Toxins 2019, 11, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maul, R.; Warth, B.; Kant, J.S.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. Investigation of the hepatic glucuronidation pattern of the Fusarium mycotoxin deoxynivalenol in various species. Chem. Res. Toxicol. 2012, 25, 2715–2717. [Google Scholar] [CrossRef]
- Schwartz-Zimmermann, H.E.; Hametner, C.; Nagl, V.; Fiby, I.; Macheiner, L.; Winkler, J.; Dänicke, S.; Clark, E.; Pestka, J.J.; Berthiller, F. Glucuronidation of deoxynivalenol (DON) by different animal species: Identification of iso-DON glucuronides and iso-deepoxy-DON glucuronides as novel DON metabolites in pigs, rats, mice, and cows. Arch. Toxicol. 2017, 91, 3857–3872. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef]
- Dänicke, S.; Brezina, U. Kinetics and metabolism of the Fusarium toxin deoxynivalenol in farm animals: Consequences for diagnosis of exposure and intoxication and carry over. Food Chem. Toxicol. 2013, 60, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Young, L.G.; McGirr, L.; Valli, V.E.; Lumsden, J.H.; Lun, A. Vomitoxin in corn fed to young pigs. J. Anim. Sci. 1983, 57, 655–664. [Google Scholar] [CrossRef]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–2. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.M.; Clark, E.S.; Pestka, J.J. Anorexia induction by the trichothecene deoxynivalenol (vomitoxin) is mediated by the release of the gut satiety hormone peptide YY. Toxicol. Sci. 2012, 130, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.R.; Pestka, J.J. Deoxynivalenol (vomitoxin)-induced cholecystokinin and glucagon-like peptide-1 release in the STC-1 enteroendocrine cell model is mediated by calcium- sensing receptor and transient receptor potential ankyrin-1 channel. Toxicol. Sci. 2015, 145, 407–417. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Alizadeh, A.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol Impairs Weight Gain and Affects Markers of Gut Health after Low-Dose, Short-Term Exposure of Growing Pigs. Toxins 2015, 7, 2071–2095. [Google Scholar] [CrossRef]
- Luongo, D.; De Luna, R.; Russo, R.; Severino, L. Effects of four Fusarium toxins (fumonisin B1, α-zearalenol, nivalenol and deoxynivalenol) on porcine whole-blood cellular proliferation. Toxicon 2008, 52, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.-R.; Islam, Z.; Pestka, J.J. Rapid, sequential activation of mitogen-activated protein kinases and transcription factors precedes proinflammatory cytokine mRNA expression in spleens of mice exposed to the trichothecene vomitoxin. Toxicol. Sci. 2003, 72, 130–142. [Google Scholar] [CrossRef] [Green Version]
- Tiemann, U.; Dänicke, S. In vivo and in vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non-reproductive and reproductive organs in female pigs: A review. Food Addit. Contam. 2007, 24, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Jakovac-Strajn, B.; Vengust, A.; Pestevsek, U. Effects of a deoxynivalenol-contaminated diet on the reproductive performance and immunoglobulin concentrations in pigs. Vet. Rec. 2009, 165, 713–718. [Google Scholar] [CrossRef]
- Yang, M.; Wu, X.; Zhang, W.; Ye, P.; Wang, Y.; Zhu, W.; Tao, Q.; Xu, Y.; Shang, J.; Zhao, D.; et al. Transcriptional analysis of deoxynivalenol-induced apoptosis of sow ovarian granulosa cell. Reprod. Domest. Anim. 2020, 55, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Goyarts, T.; Dänicke, S.; Brüssow, K.-P.; Valenta, H.; Ueberschär, K.-H.; Tiemann, U. On the transfer of the Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from sows to their fetuses during days 35–70 of gestation. Toxicol. Lett. 2007, 171, 38–49. [Google Scholar] [CrossRef]
- Dänicke, S.; Brüssow, K.P.; Goyarts, T.; Valenta, H.; Ueberschär, K.H.; Tiemann, U. On the transfer of the Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) from the sow to the full-term piglet during the last third of gestation. Food Chem. Toxicol. 2007, 45, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Wippermann, W.; Heckmann, A.; Jäger, K.; Dänicke, S.; Schoon, H.-A. Exposure of pregnant sows to deoxynivalenol during 35–70 days of gestation does not affect pathomorphological and immunohistochemical properties of fetal organs. Mycotoxin Res. 2018, 34, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, J.A.; Aiyer, V.I.A.; Hamonic, G.; Beaulieu, A.D.; Columbus, D.A.; Wilson, H.L. Molecular and physiological effects on the small intestine of weaner pigs following feeding with deoxynivalenol-contaminated feed. Toxins 2018, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Accensi, F.; Pinton, P.; Callu, P.; Abella-Bourges, N.; Guelfi, J.-F.; Grosjean, F.; Oswald, I.P. Ingestion of low doses of deoxynivalenol does not affect hematological, biochemical, or immune responses of piglets. J. Anim. Sci. 2006, 84, 1935. [Google Scholar] [CrossRef] [Green Version]
- Sayyari, A.; Framstad, T.; Krogenæs, A.K.; Sivertsen, T. Effects of feeding naturally contaminated deoxynivalenol diets to sows during late gestation and lactation in a high-yield specific pathogen-free herd. Porc. Heal. Manag. 2018, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Kollarczik, B.; Gareis, M.; Hanelt, M. In vitro transformation of theFusarium mycotoxins deoxynivalenol and zearalenone by the normal gut microflora of pigs. Nat. Toxins 1994, 2, 105–110. [Google Scholar] [CrossRef]
- Stastny, K.; Stepanova, H.; Hlavova, K.; Faldyna, M. Identification and determination of deoxynivalenol (DON) and deepoxy-deoxynivalenol (DOM-1) in pig colostrum and serum using liquid chromatography in combination with high resolution mass spectrometry (LC-MS/MS (HR)). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1126–1127, 121735. [Google Scholar] [CrossRef]
- Dänicke, S.; Beyer, M.; Breves, G.; Valenta, H.; Humpf, H.U. Effects of oral exposure of pigs to deoxynivalenol (DON) sulfonate (DONS) as the non-toxic derivative of DON on tissue residues of DON and de-epoxy-DON and on DONS blood levels. Food Addit. Contam.-Part A Chem. Anal. Control. Expo. Risk Assess 2010, 27, 1558–1565. [Google Scholar] [CrossRef]
- Prelusky, D.B.; Hartin, K.E.; Trenholm, H.L.; Miller, J.D. Pharmacokinetic Fate of 14 C-Labeled Deoxynivalenol in Swine. Toxicol. Sci. 1988, 10, 276–286. [Google Scholar] [CrossRef]
- Paulick, M.; Winkler, J.; Kersten, S.; Schatzmayr, D.; Schwartz-Zimmermann, H.E.; Dänicke, S. Studies on the bioavailability of deoxynivalenol (DON) and DON sulfonate (DONS) 1, 2, and 3 in pigs fed with sodium sulfite-treated DON-contaminated maize. Toxins 2015, 7, 4622–4644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Krska, R. New insights into the human metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone. Toxicol. Lett. 2013, 220, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, A.C.; Todd See, M.; Hansen, J.A.; Kim, Y.B.; De Souza, A.L.P.; Middleton, T.F.; Kim, S.W. The use of feed additives to reduce the effects of aflatoxin and deoxynivalenol on pig growth, organ health and immune status during chronic exposure. Toxins 2013, 5, 1261–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuča, K.; Yuan, Z. Metabolic pathways of trichothecenes. Drug Metab. Rev. 2010, 42, 250–267. [Google Scholar] [CrossRef] [PubMed]
- Maul, R.; Warth, B.; Schebb, N.H.; Krska, R.; Koch, M.; Sulyok, M. In vitro glucuronidation kinetics of deoxynivalenol by human and animal microsomes and recombinant human UGT enzymes. Arch. Toxicol. 2015, 89, 949–960. [Google Scholar] [CrossRef]
- Gail McCarver, D.; Hines, R.N. The ontogeny of human drug-metabolizing enzymes: Phase II conjugation enzymes and regulatory mechanisms. J. Pharmacol. Exp. Ther. 2002, 300, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Pretheeban, M.; Hammond, G.; Bandiera, S.; Riggs, W.; Rurak, D. Ontogenesis of UDP-glucuronosyltransferase enzymes in sheep. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2011, 159, 159–166. [Google Scholar] [CrossRef]
- Hu, S.X. Age-related change of hepatic uridine diphosphate glucuronosyltransferase and sulfotransferase activities in male chickens and pigs. J. Vet. Pharmacol. Ther. 2017, 40, 270–278. [Google Scholar] [CrossRef]
- Oswald, I.P.; Marin, D.E.; Bouhet, S.; Pinton, P.; Taranu, I.; Accensi, F. Immunotoxicological risk of mycotoxins for domestic animals. Food Addit. Contam. 2005, 22, 354–360. [Google Scholar] [CrossRef]
- Goyarts, T.; Dänicke, S.; Tiemann, U.; Rothkötter, H.J. Effect of the Fusarium toxin deoxynivalenol (DON) on IgA, IgM and IgG concentrations and proliferation of porcine blood lymphocytes. Toxicol. Vitr. 2006, 20, 858–867. [Google Scholar] [CrossRef]
- Ferrari, L.; Cantoni, A.M.; Borghetti, P.; De Angelis, E.; Corradi, A. Cellular immune response and immunotoxicity induced by DON (deoxynivalenol) in piglets. Vet. Res. Commun. 2009, 33, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.E.; Song, J.; Lee, H.J.; Kim, M.; Kim, D.W.; Jung, H.J.; Kim, B.; Lee, Y.; Yu, D.; Kim, D.W.; et al. Effects of high levels of deoxynivalenol and zearalenone on growth performance, and hematological and immunological parameters in pigs. Toxins 2018, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J.; Dong, W.; Warner, R.L.; Rasooly, L.; Bondy, G.S.; Brooks, K.H. Elevated membrane IgA+ and CD4+ (T helper) populations in murine peyer’s patch and splenic lymphocytes during dietary administration of the trichothecene vomitoxin (deoxynivalenol). Food Chem. Toxicol. 1990, 28, 409–420. [Google Scholar] [CrossRef]
- Rasooly, L.; Pestka, J.J. Vomitoxin-induced dysregulation of serum IgA, IgM and IgG reactive with gut bacterial and self antigens. Food Chem. Toxicol. 1992, 30, 499–504. [Google Scholar] [CrossRef]
- Islam, M.R.; Roh, Y.S.; Kim, J.; Lim, C.W.; Kim, B. Differential immune modulation by deoxynivalenol (vomitoxin) in mice. Toxicol. Lett. 2013, 221, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Mechanisms of deoxynivalenol-induced gene expression and apoptosis. Food Addit. Contam.-Part A Chem. Anal. Control. Expo. Risk Assess 2008, 25, 1128–1140. [Google Scholar] [CrossRef]
- Wada, K.; Hashiba, Y.; Ohtsuka, H.; Kohiruimaki, M.; Masui, M.; Kawamura, S.; Endo, H.; Ogata, Y. Effects of mycotoxins on mitogen-stimulated proliferation of bovine peripheral blood mononuclear cells. J. Vet. Med. Sci. 2008, 70, 193–196. [Google Scholar] [CrossRef] [Green Version]
- Novak, B.; Vatzia, E.; Springler, A.; Pierron, A.; Gerner, W.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G.; Mayer, E. Bovine peripheral blood mononuclear cells are more sensitive to deoxynivalenol than those derived from poultry and swine. Toxins 2018, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, M.; Obremski, K.; Gajęcka, M.; Gajęcki, M.; Zielonka, Ł. Changes in the Subpopulations of Porcine Peripheral Blood Lymphocytes Induced by Exposure to Low Doses of Zearalenone (ZEN) and Deoxynivalenol (DON). Molecules 2016, 21, 557. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, M.; Jakimiuk, E.; Baranowski, M.; Gajȩcka, M.; Zielonka, Ł.; Gajȩcki, M.T. The effect of deoxynivalenol on selected populations of immunocompetent cells in porcine blood-a preliminary study. Molecules 2017, 22, 691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlavová, K.; Štěpánová, H.; Šťastný, K.; Levá, L.; Hodkovicová, N.; Vícenová, M.; Matiašovic, J.; Faldyna, M.; Št’astný, K.; Levá, L.; et al. Minimal concentrations of deoxynivalenol reduce cytokine production in individual lymphocyte populations in pigs. Toxins 2020, 12, 190. [Google Scholar] [CrossRef] [Green Version]
- Swamy, H.V.L.N.; Smith, T.K.; MacDonald, E.J.; Karrow, N.A.; Woodward, B.; Boermans, H.J. Effects of feeding a blend of grains naturally contaminated with Fusarium mycotoxins on growth and immunological measurements of starter pigs, and the efficacy of a polymeric glucomannan mycotoxin adsorbent1. J. Anim. Sci. 2003, 81, 2792–2803. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, H.; Samankova, P.; Leva, L.; Sinkora, J.; Faldyna, M. Early postnatal development of the immune system in piglets: The redistribution of T lymphocyte subsets. Cell. Immunol. 2007, 249, 73–79. [Google Scholar] [CrossRef]
- Šinkora, M.; Butler, J.E. The ontogeny of the porcine immune system. Dev. Comp. Immunol. 2009, 33, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Saalmüller, A.; Werner, T.; Fachinger, V. T-helper cells from naive to committed. Vet. Immunol. Immunopathol. 2002, 87, 137–145. [Google Scholar] [CrossRef]
- Vatzia, E.; Pierron, A.; Saalmüller, A.; Mayer, E.; Gerner, W. Deoxynivalenol affects proliferation and expression of activation-related molecules in major porcine T-cell subsets. Toxins 2019, 11, 644. [Google Scholar] [CrossRef] [Green Version]
- Bloom, B.R.; Salgame, P.; Diamond, B. Revisiting and revising suppressor T cells. Immunol. Today 1992, 13, 131–136. [Google Scholar] [CrossRef]
- Käser, T.; Gerner, W.; Mair, K.; Hammer, S.E.; Patzl, M.; Saalmüller, A. Current knowledge on porcine regulatory T cells. Vet. Immunol. Immunopathol. 2012, 148, 136–138. [Google Scholar] [CrossRef]
- Savard, C.; Pinilla, V.; Provost, C.; Gagnon, C.A.; Chorfi, Y. In vivo effect of deoxynivalenol (DON) naturally contaminated feed on porcine reproductive and respiratory syndrome virus (PRRSV) infection. Vet. Microbiol. 2014, 174, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Vandenbroucke, V.; Croubels, S.; Martel, A.; Verbrugghe, E.; Goossens, J.; Van Deun, K.; Boyen, F.; Thompson, A.; Shearer, N.; De Backer, P.; et al. The mycotoxin deoxynivalenol potentiates intestinal inflammation by Salmonella typhimurium in porcine ileal loops. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinton, P.; Accensi, F.; Beauchamp, E.; Cossalter, A.-M.M.; Callu, P.; Grosjean, F.; Oswald, I.P. Ingestion of deoxynivalenol (DON) contaminated feed alters the pig vaccinal immune responses. Toxicol. Lett. 2008, 177, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Savard, C.; Gagnon, C.A.; Chorfi, Y. Deoxynivalenol (DON) naturally contaminated feed impairs the immune response induced by porcine reproductive and respiratory syndrome virus (PRRSV) live attenuated vaccine. Vaccine 2015, 33, 3881–3886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartkiene, E.; Zavistanaviciute, P.; Lele, V.; Ruzauskas, M.; Bartkevics, V.; Bernatoniene, J.; Gallo, P.; Tenore, G.C.; Santini, A. Lactobacillus plantarum LUHS135 and paracasei LUHS244 as functional starter cultures for the food fermentation industry: Characterisation, mycotoxin-reducing properties, optimisation of biomass growth and sustainable encapsulation by using dairy by-products. LWT 2018, 93, 649–658. [Google Scholar] [CrossRef]
- McEwan, A.D.; Fisher, E.W.; Selman, I.E.; Penhale, W.J. A turbidity test for the estimation of immune globulin levels in neonatal calf serum. Clin. Chim. Acta 1970, 27, 155–163. [Google Scholar] [CrossRef]
- Stepanova, H.; Mensikova, M.; Chlebova, K.; Faldyna, M. CD4+ and γδTCR+ T lymphocytes are sources of interleukin-17 in swine. Cytokine 2012, 58, 152–157. [Google Scholar] [CrossRef]
- Volf, J.; Boyen, F.; Faldyna, M.; Pavlova, B.; Navratilova, J.; Rychlik, I. Cytokine response of porcine cell lines to Salmonella enterica serovar Typhimurium and its hilA and ssrA mutants. Zoonoses Public Health 2007, 54, 286–293. [Google Scholar] [CrossRef]
- Stepanova, H.; Pavlova, B.; Stromerova, N.; Matiasovic, J.; Kaevska, M.; Pavlik, I.; Faldyna, M. Cell-mediated immune response in swine infected with Mycobacterium avium subsp. avium. Vet. Immunol. Immunopathol. 2011, 142, 107–112. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Week | 3 Weeks | 18 Weeks | ||||
|---|---|---|---|---|---|---|
| Control (n = 36) | DON (n = 39) | Control (n = 33) | DON (n = 38) | Control (n = 27) | DON (n = 32) | |
| RBCs (1012/L) | 4.1 (0.9) | 3.9 (0.7) | 5.4 (0.5) | 5.4 (0.5) | 6.8 (0.4) | 6.9 (0.4) |
| HTK (%) | 27.9 (4.5) | 28.1 (3.1) | 30.8 (4.0) | 30.9 (4.2) | 35.3 (2.0) | 38.1 (2.6) |
| Hb (g/L) | 87.3 (15.8) | 88.5 (10.8) | 101.1 (13.0) | 102.4 (15.2) | 113.9 (6.8) | 121.8 (9.2) |
| WBC (109/L) | 13.9 (10.0) | 11.7 (3.8) | 8.3 (3.0) | 7.5 (2.4) | 14.5 (3.0) | 12.8 (2.9) |
| Lymphocytes (%) | 47.5 (18.1) | 53.2 (16.2) | 59.9 (14.9) | 62.0 (13.7) | 76.0 (9.5) | 80.5 (9.6) |
| Neutrophils (%) | 49.4 (18.5) | 43.4 (16.5) | 38.1 (14.4) | 36.9 (13.6) | 21.2 (9.7) | 17.0 (9.4) |
| Band neutrophils (%) | 1.4 (1.4) | 1.7 (1.9) | 0.9 (0.7) | 0.4 (0.5) | 0.3 (0.6) | 0.4 (0.5) |
| Monocytes (%) | 0.5 (0.7) | 0.7 (0.7) | 0.3 (0.5) | 0.2 (0.3) | 0.6 (0.7) | 0.5 (0.8) |
| Eosinophils (%) | 0.7 (0.5) | 0.5 (0.6) | 0.5 (0.5) | 0.3 (0.4) | 1.1. (0.8) | 1.1 (0.8) |
| Basophils (%) | 0.4 (0.5) | 0.5 (0.6) | 0.3 (0.4) | 0.2 (0.3) | 0.9 (0.9) | 0.6 (0.6) |
| NRBCs (counts/100 WBCs) | 14.1 (20.6) | 12.9 (12.0) | 5.4 (5.5) | 3.0 (3.8) | 1.1 (1.3) | 0.6 (0.7) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štěpánová, H.; Hlavová, K.; Šťastný, K.; Gopfert, E.; Levá, L.; Faldyna, M. Maternal Exposure Results in Long-Term Deoxynivalenol Persistence in Piglets’ Plasma and Modulates the Immune System. Toxins 2020, 12, 615. https://doi.org/10.3390/toxins12100615
Štěpánová H, Hlavová K, Šťastný K, Gopfert E, Levá L, Faldyna M. Maternal Exposure Results in Long-Term Deoxynivalenol Persistence in Piglets’ Plasma and Modulates the Immune System. Toxins. 2020; 12(10):615. https://doi.org/10.3390/toxins12100615
Chicago/Turabian StyleŠtěpánová, Hana, Karolina Hlavová, Kamil Šťastný, Eduard Gopfert, Lenka Levá, and Martin Faldyna. 2020. "Maternal Exposure Results in Long-Term Deoxynivalenol Persistence in Piglets’ Plasma and Modulates the Immune System" Toxins 12, no. 10: 615. https://doi.org/10.3390/toxins12100615