First Genome-Based Characterisation and Staphylococcal Enterotoxin Production Ability of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus Strains Isolated from Ready-to-Eat Foods in Algiers (Algeria)

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Genomic Statistics
2.2. Multilocus Sequence and spa Typing
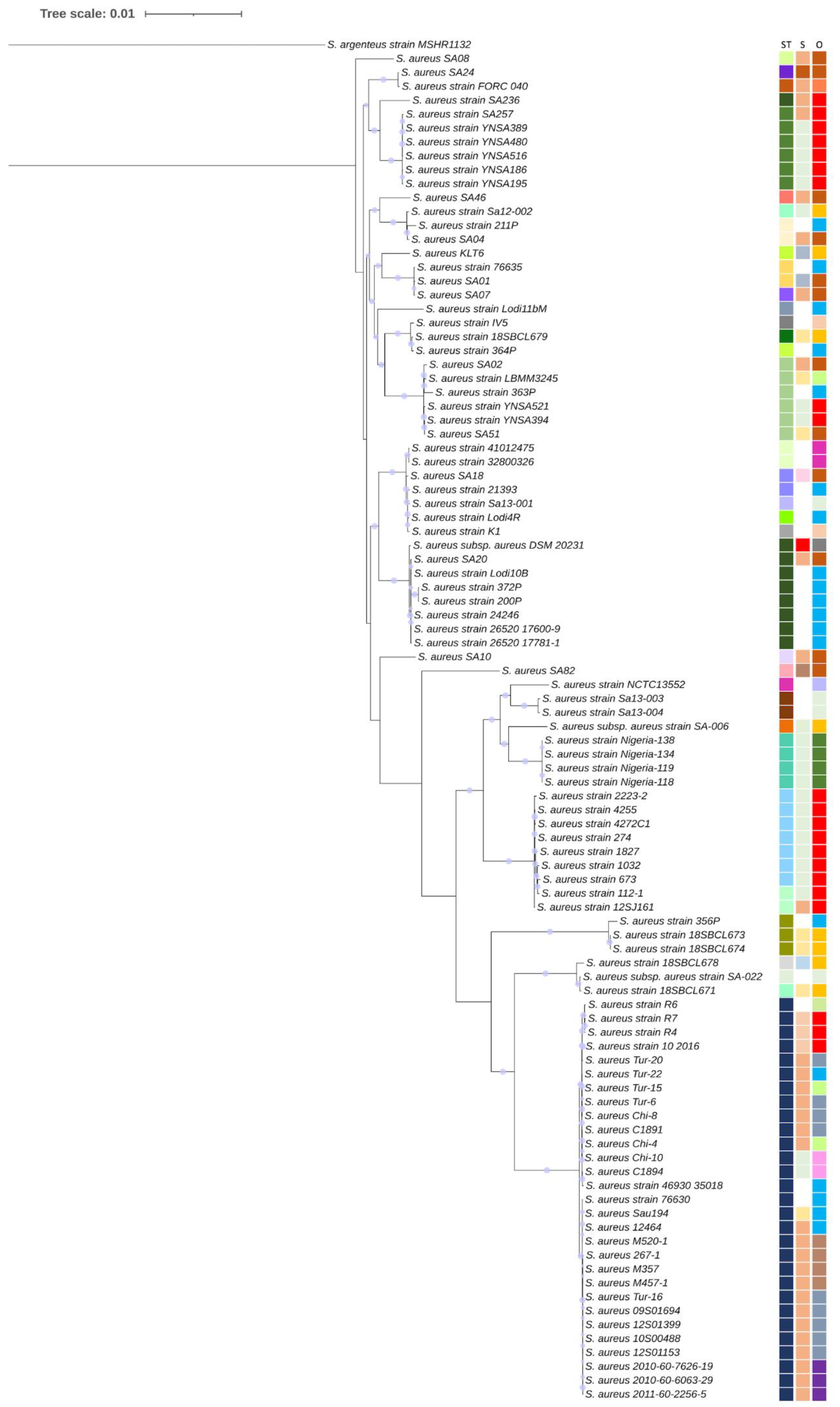
2.3. Phylogenetic Analysis
2.4. Pan-Genome Analysis
2.5. Enterotoxin Genes Presence and Localisation
2.6. Antibiotic Resistance Genetic Determinants
2.7. Staphylococcal Chromosomal Cassette mec
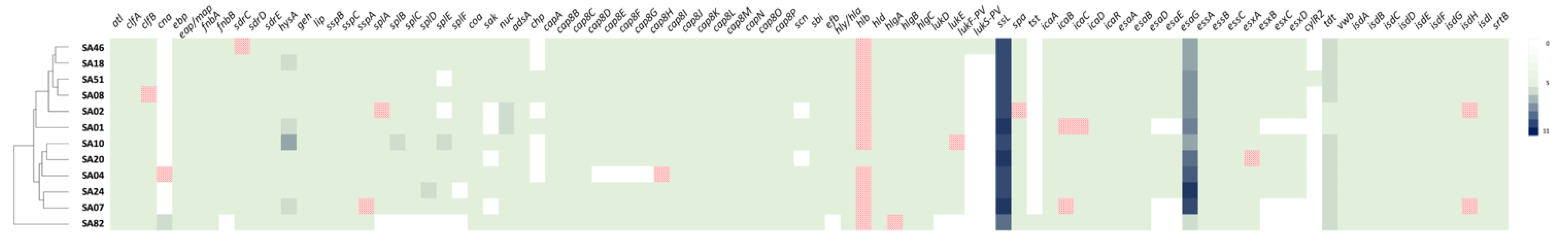
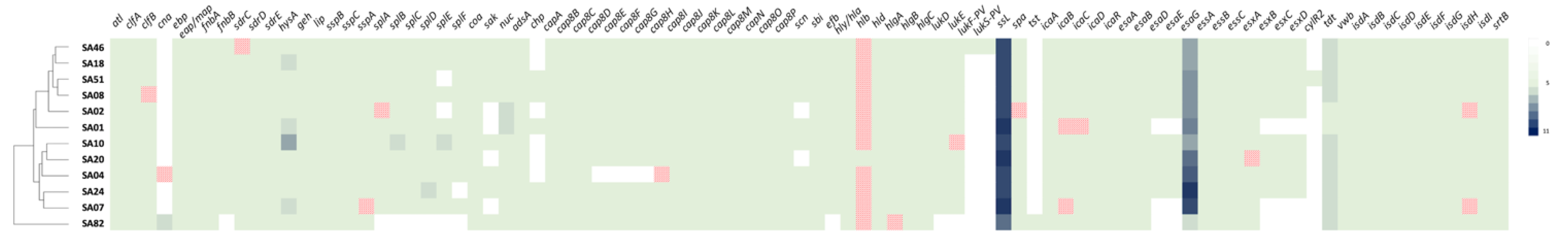
2.8. Virulence Determinants
2.9. Growth of S. aureus Strains in BHI + YE Broth and Production of Staphylococcal Enterotoxins
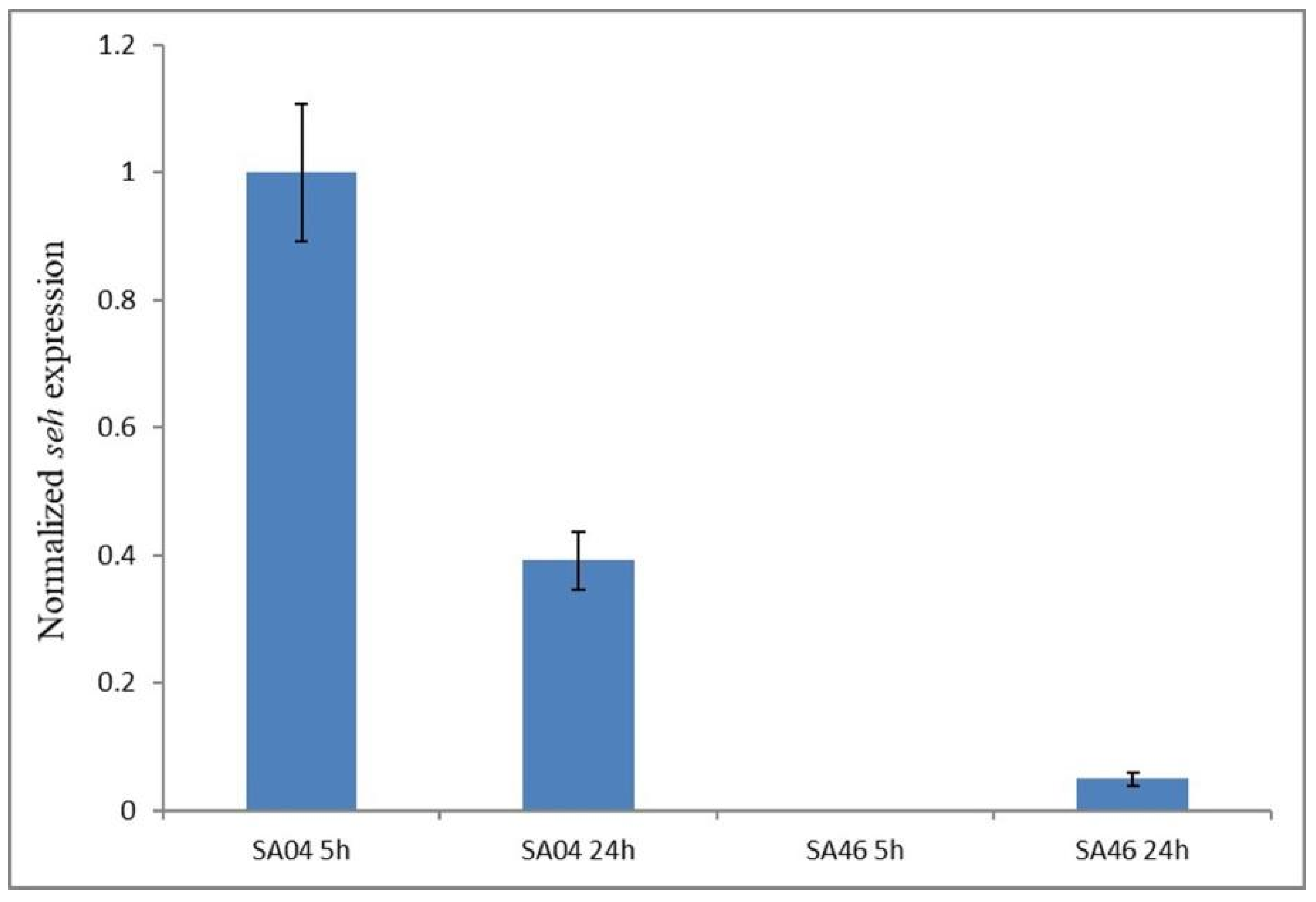
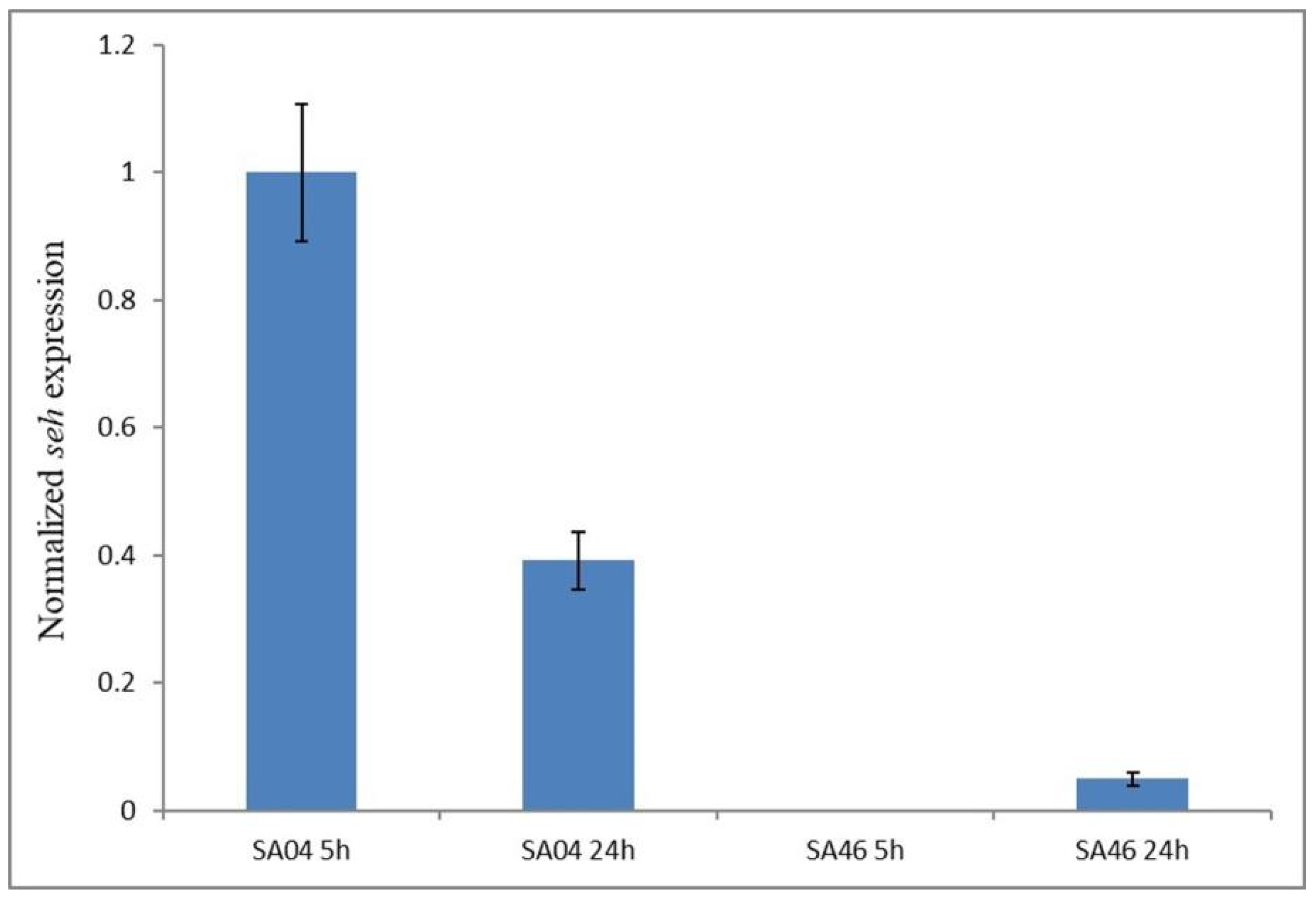
2.10. Expression of the seh Gene
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. S. aureus Strains Used in the Study
5.2. Whole-Genome Sequencing and Analysis
5.3. Bioinformatic Methods
5.3.1. Gene Prediction, ANI, and Phylogenomic Analysis
5.3.2. MLST and spa Typing
5.3.3. Antibiotic Resistance and Virulence Determinants Analyses
5.4. Cultivation of S. aureus for Staphylococcal Enterotoxin Production Assessment
5.5. Sandwich ELISA for SEA, SEB, SED, SEH, and SER Detection
5.6. RNA Extraction and Reverse Transcription–Quantitative PCR to Assess seh Gene Expression
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dicks, J.; Turnbull, J.D.; Russell, J.; Parkhill, J.; Alexander, S. Genome sequencing of a historic Staphylococcus aureus collection reveals new enterotoxin genes and sheds light on the evolution and genomic organization of this key virulence gene family. J. Bacteriol. 2021, 203, e00587-20. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Chieffi, D.; Fanelli, F.; Logrieco, A.F.; Cho, G.-S.; Kabisch, J.; Böhnlein, C.; Franz, C.M.A.P. Microbial quality and safety of milk and milk products in the 21st century. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2013–2049. [Google Scholar] [CrossRef] [PubMed]
- Mekhloufi, O.A.; Chieffi, D.; Hammoudi, A.; Bensefia, S.A.; Fanelli, F.; Fusco, V. Prevalence, enterotoxigenic potential and antimicrobial resistance of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA) isolated from Algerian ready to eat foods. Toxins 2021, 13, 835. [Google Scholar] [CrossRef]
- Fishovitz, J.; Hermoso, J.A.; Chang, M.; Mobashery, S. Penicillin-binding protein 2a of methicillin-resistant Staphylococcus aureus. IUBMB Life 2014, 66, 572–577. [Google Scholar] [CrossRef] [Green Version]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol. 2001, 166, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chieffi, D.; Fanelli, F.; Cho, G.-S.; Schubert, J.; Blaiotta, G.; Franz, C.M.A.P.; Bania, J.; Fusco, V. Novel insights into the enterotoxigenic potential and genomic background of Staphylococcus aureus isolated from raw milk. Food Microbiol. 2020, 90, 103482. [Google Scholar] [CrossRef]
- Achek, R.; El-Adawy, H.; Hotzel, H.; Hendam, A.; Tomaso, H.; Ehricht, R.; Neubauer, H.; Nabi, I.; Hamdi, T.M.; Monecke, S. Molecular Characterization of Staphylococcus aureus Isolated from Human and Food Samples in Northern Algeria. Pathogens 2021, 10, 1276. [Google Scholar] [CrossRef]
- Chaalal, W.; Chaalal, N.; Bourafa, N.; Kihal, M.; Diene, S.M.; Rolain, J.-M. Characterization of Staphylococcus aureus Isolated from Food Products in Western Algeria. Foodborne Pathog. Dis. 2018, 15, 353–360. [Google Scholar] [CrossRef]
- Titouche, Y.; Hakem, A.; Houali, K.; Meheut, T.; Vingadassalon, N.; Ruiz-Ripa, L.; Salmi, D.; Chergui, A.; Chenouf, N.; Hennekinne, J.A.; et al. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) ST8 in raw milk and traditional dairy products in the Tizi Ouzou area of Algeria. J. Dairy Sci. 2019, 102, 6876–6884. [Google Scholar] [CrossRef]
- Titouche, Y.; Houali, K.; Ruiz-Ripa, L.; Vingadassalon, N.; Nia, Y.; Fatihi, A.; Cauquil, A.; Bouchez, P.; Bouhier, L.; Torres, C.; et al. Enterotoxin genes and antimicrobial resistance in Staphylococcus aureus isolated from food products in Algeria. J. Appl. Microbiol. 2020, 129, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Breurec, S.; Fall, C.; Pouillot, R.; Boisier, P.; Brisse, S.; Diene-Sarr, F.; Djibo, S.; Etienne, J.; Fonkoua, M.C.; Perrier-Gros-Claude, J.D.; et al. Epidemiology of methicillin-susceptible Staphylococcus aureus lineages in five major African towns: High prevalence of Panton-Valentine leukocidin genes. Clin. Microbiol. Infect. 2011, 17, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Gharsa, H.; Ben Slama, K.; Zarazaga, M.; Torres, C. Staphylococcus aureus in animals and food: Methicillin resistance, prevalence and population structure. A review in the African continent. Microorganisms 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aung, M.; San, T.; Aye, M.; Mya, S.; Maw, W.; Zan, K.; Htut, W.; Kawaguchiya, M.; Urushibara, N.; Kobayashi, N. Prevalence and Genetic Characteristics of Staphylococcus aureus and Staphylococcus argenteus Isolates Harboring Panton-Valentine Leukocidin, Enterotoxins, and TSST-1 Genes from Food Handlers in Myanmar. Toxins 2017, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.J.; Seo, K.S.; Cartwright, R.A.; Connelley, T.; Chuang-Smith, O.N.; Merriman, J.A.; Guinane, C.M.; Park, J.Y.; Bohach, G.A.; Schlievert, P.M.; et al. A novel core genome-encoded superantigen contributes to lethality of community-associated MRSA necrotizing pneumonia. PLoS Pathog. 2011, 7, e1002271. [Google Scholar] [CrossRef] [Green Version]
- Roetzer, A.; Haller, G.; Beyerly, J.; Geier, C.B.; Wolf, H.M.; Gruener, C.S.; Model, N.; Eibl, M.M. Genotypic and phenotypic analysis of clinical isolates of Staphylococcus aureus revealed production patterns and hemolytic potentials unlinked to gene profiles and source. BMC Microbiol. 2016, 16, 13. [Google Scholar] [CrossRef] [Green Version]
- Borst, D.W.; Betley, M.J. Phage-associated differences in staphylococcal enterotoxin A gene (sea) expression correlate with sea allele class. Infect. Immun. 1994, 62, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Johler, S.; Sihto, H.-M.; Macori, G.; Stephan, R. Sequence variability in staphylococcal enterotoxin genes seb, sec, and sed. Toxins 2016, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Baba, T.; Takeuchi, F.; Kuroda, M.; Yuzawa, H.; Aoki, K.; Oguchi, A.; Nagai, Y.; Iwama, N.; Asano, K.; Naimi, T.; et al. Genome and virulence determinants of high virulence community-acquired MRSA. Lancet 2002, 359, 1819–1827. [Google Scholar] [CrossRef]
- Betley, M.J.; Mekalanos, J.J. Nucleotide sequence of the type A staphylococcal enterotoxin gene. J. Bacteriol. 1988, 170, 34–41. [Google Scholar] [CrossRef]
- Chua, K.; Seemann, T.; Harrison, P.F.; Davies, J.K.; Coutts, S.J.; Chen, H.; Haring, V.; Moore, R.; Howden, B.P.; Stinear, T.P. Complete Genome Sequence of Staphylococcus aureus Strain JKD6159, a Unique Australian Clone of ST93-IV Community Methicillin-Resistant Staphylococcus aureus. J. Bacteriol. 2010, 192, 5556–5557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collery, M.M.; Smyth, C.J. Rapid differentiation of Staphylococcus aureus isolates harbouring egc loci with pseudogenes psient1 and psient2 and the selu or seluv gene using PCR-RFLP. J. Med. Microbiol. 2007, 56, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egyir, B.; Hadjirin, N.F.; Gupta, S.; Owusu, F.; Agbodzi, B.; Adogla-Bessa, T.; Addo, K.K.; Stegger, M.; Larsen, A.R.; Holmes, M.A. Whole-genome sequence profiling of antibiotic-resistant Staphylococcus aureus isolates from livestock and farm attendants in Ghana. J. Glob. Antimicrob. Resist. 2020, 22, 527–532. [Google Scholar] [CrossRef]
- Gawlik, D.; Ruppelt-Lorz, A.; Muller, E.; Reißig, A.; Hotzel, H.; Braun, S.D.; Söderquist, B.; Ziegler-Cordts, A.; Stein, C.; Pletz, M.W.; et al. Molecular investigations on a chimeric strain of Staphylococcus aureus sequence type 80. PLoS ONE 2020, 15, e0232071. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, H.J.; Mathisen, T.; Lovseth, A.; Omoe, K.; Qvale, K.S.; Loncarevic, S. An outbreak of staphylococcal food poisoning caused by enterotoxin H in mashed potato made with raw milk. FEMS Microbiol. Lett. 2005, 252, 267–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley, R.J.; Ting, Y.T.; Clow, F.; Young, P.G.; Radcliff, F.J.; Choi, J.M.; Sequeira, R.P.; Holtfreter, S.; Baker, H.; Fraser, J.D. Staphylococcal enterotoxin-like X (SElX) is a unique superantigen with functional features of two major families of staphylococcal virulence factors. PLoS Pathog. 2017, 13, e1006549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabirova, J.S.; Xavier, B.B.; Hernalsteens, J.P.; De Greve, H.; Ieven, M.; Goossens, H.; Malhotra-Kumar, S. Complete Genome Sequences of Two Prolific Biofilm-Forming Staphylococcus aureus Isolates Belonging to USA300 and EMRSA-15 Clonal Lineages. Genome Announc. 2014, 2, e00610-14. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.Y.; Jarraud, S.; Lemercier, B.; Cozon, G.; Echasserieau, K.; Etienne, J.; Gougeon, M.L.; Lina, G.; Vandenesch, F. Staphylococcal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect. Immun. 2006, 74, 4724–4734. [Google Scholar] [CrossRef] [Green Version]
- Tuffs, S.W.; James, D.B.A.; Bestebroer, J.; Richards, A.C.; Goncheva, M.I.; O’Shea, M.; Wee, B.A.; Seo, K.S.; Schlievert, P.M.; Lengeling, A.; et al. The Staphylococcus aureus superantigen SElX is a bifunctional toxin that inhibits neutrophil function. PLoS Pathog. 2017, 13, e1006461. [Google Scholar] [CrossRef] [Green Version]
- Utter, B.; Deutsch, D.R.; Schuch, R.; Winer, B.Y.; Verratti, K.; Bishop-Lilly, K.; Sozhamannan, S.; Fischetti, V.A. Beyond the Chromosome: The Prevalence of Unique Extra-Chromosomal Bacteriophages with Integrated Virulence Genes in Pathogenic Staphylococcus aureus. PLoS ONE. 2014, 9, E100502. [Google Scholar] [CrossRef]
- Wan, T.W.; Liu, Y.J.; Wang, Y.T.; Lin, Y.T.; Hsu, J.C.; Tsai, J.C.; Chiu, H.C.; Hsueh, P.R.; Hung, W.C.; Teng, L.J. Potentially conjugative plasmids harboring Tn6636, a multidrug-resistant and composite mobile element, in Staphylococcus aureus. J. Microbiol. Immunol. Infect. 2022, 55, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Okumura, K.; Shimomura, Y.; Murayama, S.Y.; Yagi, J.; Ubukata, K.; Kirikae, T.; Miyoshi-Akiyama, T. Evolutionary paths of streptococcal and staphylococcal superantigens. BMC Genom. 2012, 13, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kläui, A.J.; Boss, R.; Graber, H.U. Characterization and comparative analysis of the Staphylococcus aureus genomic island vSaβ: An in silico Approach. J. Bacteriol. 2019, 201, e00777-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, K.A.; Peng, H.; Luebke, J.L.; Chang, F.M.; Giedroc, D.P. Conformational analysis and chemical reactivity of the multidomain sulfurtransferase, Staphylococcus aureus CstA. Biochemistry 2015, 54, 2385–2398. [Google Scholar] [CrossRef] [PubMed]
- Sato’o, Y.; Omoe, K.; Ono, H.K.; Nakane, A.; Hu, D.L. A novel comprehensive analysis method for Staphylococcus aureus pathogenicity islands. Microbiol. Immunol. 2013, 57, 91–99. [Google Scholar] [CrossRef]
- Shearer, J.E.; Wireman, J.; Hostetler, J.; Forberger, H.; Borman, J.; Gill, J.; Sanchez, S.; Mankin, A.; Lamarre, J.; Lindsay, J.A.; et al. Major families of multiresistant plasmids from geographically and epidemiologically diverse staphylococci. G3 Genes Genomes Genet. 2011, 1, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Rowland, S.J.; Dyke, K.G. Characterization of the staphylococcal beta-lactamase transposon Tn552. EMBO J. 1989, 18, 2761–2773. [Google Scholar] [CrossRef]
- Rowland, S.J.; Dyke, K.G. Tn552, a novel transposable element from Staphylococcus aureus. Mol Microbiol. 1990, 4, 961–975. [Google Scholar] [CrossRef]
- Jensen, S.O.; Lyon, B.R. Genetics of antimicrobial resistance in Staphylococcus aureus. Future Microbiol. 2009, 4, 565–582. [Google Scholar] [CrossRef]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Gordon, N.C.; Price, J.R.; Cole, K.; Everitt, R.; Morgan, M.; Finney, J.; Kearns, A.M.; Pichon, B.; Young, B.; Wilson, D.J.; et al. Prediction of Staphylococcus aureus antimicrobial resistance by whole-genome sequencing. J. Clin. Microbiol. 2014, 52, 1182–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumitrescu, O.; Dauwalder, O.; Boisset, S.; Reverdy, M.É.; Tristan, A.; Vandenesch, F. Staphylococcus aureus resistance to antibiotics: Key points in 2010. Med. Sci. 2010, 26, 943–949. [Google Scholar] [CrossRef]
- Heatmapper. Available online: http://www.heatmapper.ca (accessed on 31 March 2022).
- McCarthy, A.J.; Lindsay, J.A. Staphylococcus aureus innate immune evasion is lineage-specific: A bioinfomatics study. Infect. Genet. Evol. 2013, 19, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Aswani, V.; Najar, F.; Pantrangi, M.; Mau, B.; Schwan, W.R.; Shukla, S.K. Virulence factor landscape of a Staphylococcus aureus sequence type 45 strain, MCRF184. BMC Genom. 2019, 20, 123. [Google Scholar] [CrossRef] [PubMed]
- Gravet, A.; Colin, D.A.; Keller, D.; Girardot, R.; Monteil, H.; Prévost, G. Characterization of a novel structural member, LukE-LukD, of the bi-component staphylococcal leucotoxins family. FEBS Lett. 1998, 436, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Rhem, M.N.; Lech, E.M.; Patti, J.M.; McDevitt, D.; Höök, M.; Jones, D.B.; Wilhelmus, K.R. The collagen-binding adhesin is a virulence factor in Staphylococcus aureus keratitis. Infect. Immun. 2000, 68, 3776–3779. [Google Scholar] [CrossRef] [Green Version]
- Agabou, A.; Ouchenane, Z.; Ngba Essebe, C.; Khemissi, S.; Chehboub, M.T.E.; Chehboub, I.B.; Sotto, A.; Dunyach-Remy, C.; Lavigne, J.P. Emergence of nasal carriage of ST80 and ST152 PVL+ Staphylococcus aureus isolates from livestock in Algeria. Toxins 2017, 9, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mairi, A.; Touati, A.; Pantel, A.; Zenati, K.; Martinez, A.Y.; Dunyach-Remy, C.; Sotto, A.; Lavigne, J.P. Distribution of toxinogenic Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus from different ecological niches in Algeria. Toxins 2019, 11, 500. [Google Scholar] [CrossRef] [Green Version]
- Ghebremedhin, B.; Olugbosi, M.O.; Raji, A.M.; Layer, F.; Bakare, R.A.; König, B.; König, W. Emergence of a community-associated methicillin-resistant Staphylococcus aureus strain with a unique resistance profile in Southwest Nigeria. J. Clin. Microbiol. 2009, 47, 2975–2980. [Google Scholar] [CrossRef] [Green Version]
- Schaumburg, F.; Ngoa, U.A.; Kösters, K.; Köck, R.; Adegnika, A.A.; Kremsner, P.G.; Lell, B.; Peters, G.; Mellmann, A.; Becker, K. Virulence factors and genotypes of Staphylococcus aureus from infection and carriage in Gabon. Clin. Microbiol. Infect. 2011, 17, 1507–1513. [Google Scholar] [CrossRef]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thapaliya, D.; Forshey, B.M.; Kadariya, J.; Quick, M.K.; Farina, S.; O’ Brien, A.; Nair, R.; Nworie, A.; Hanson, B.; Kates, A.; et al. Prevalence and molecular characterization of Staphylococcus aureus in commercially available meat over a one-year period in Iowa, USA. Food Microbiol. 2017, 65, 122–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mama, O.M.; Morales, L.; Ruiz-Ripa, L.; Zarazaga, M.; Torres, C. High prevalence of multidrug resistant S. aureus-CC398 and frequent detection of enterotoxin genes among non-CC398 S. aureus from pig-derived food in Spain. Int. J. Food Microbiol. 2020, 320, 108510. [Google Scholar] [CrossRef] [PubMed]
- Casman, E.P.; Bennett, R.W.; Dorsey, A.E.; Issa, J.A. Identification of a fourth staphylococcal enterotoxin, enterotoxin D. J. Bacteriol. 1967, 94, 1875–1882. [Google Scholar] [CrossRef] [Green Version]
- Bergdoll, M.S.; Borja, C.R.; Robbins, R.N.; Weiss, K.F. Identification of enterotoxin E. Infect Immun. 1971, 4, 593–595. [Google Scholar] [CrossRef] [Green Version]
- Benkerroum, N. Staphylococcal enterotoxins and enterotoxin-like toxins with special reference to dairy products: An overview. Crit. Rev. Food Sci. Nutr. 2018, 58, 1943–1970. [Google Scholar] [CrossRef]
- Ikeda, T.; Tamate, N.; Yamaguchi, K.; Makino, S. Mass outbreak of food poisoning disease caused by small amounts of staphylococcal enterotoxins A and H. Appl. Environ. Microbiol. 2005, 71, 2793–2795. [Google Scholar] [CrossRef] [Green Version]
- Johler, S.; Giannini, P.; Jermini, M.; Hummerjohann, J.; Baumgartner, A.; Stephan, R. Further evidence for staphylococcal food poisoning outbreaks caused by egc-encoded enterotoxins. Toxins 2015, 7, 997–1004. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kobayashi, M.; Matsushita, S.; Uehara, S.; Kato, R.; Sato’o, Y.; Ono, H.K.; Sadamasu, K.; Kai, A.; Kamata, Y. Detection of the staphylococcal enterotoxin D-like gene from staphylococcal food poisoning isolates over the last two decades in Tokyo. J. Vet. Med. Sci. 2015, 77, 905–911. [Google Scholar] [CrossRef] [Green Version]
- Umeda, K.; Nakamura, H.; Yamamoto, K.; Nishina, N.; Yasufuku, K.; Hirai, Y.; Hirayama, T.; Goto, K.; Hase, A.; Ogasawara, J. Molecular and epidemiological characterization of staphylococcal foodborne outbreak of Staphylococcus aureus harboring seg, sei, sem, sen, seo, and selu genes without production of classical enterotoxins. Int. J. Food Microbiol. 2017, 256, 30–35. [Google Scholar] [CrossRef]
- Denayer, S.; Delbrassinne, L.; Nia, Y.; Botteldoorn, N. Food-Borne Outbreak investigation and molecular typing: High diversity of Staphylococcus aureus strains and importance of toxin detection. Toxins 2017, 9, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johler, S.; Weder, D.; Bridy, C.; Huguenin, M.-C.; Robert, L.; Hummerjohann, J.; Stephan, R. Outbreak of staphylococcal food poisoning among children and staff at a Swiss boarding school due to soft cheese made from raw milk. J. Dairy Sci. 2015, 98, 2944–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asao, T.; Kumeda, Y.; Kawai, T.; Shibata, T.; Oda, H.; Haruki, K.; Nakazawa, H.; Kozaki, S. An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: Estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 2003, 130, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Evenson, M.L.; Ward Hinds, M.; Bernstein, R.S.; Bergdoll, M.S. Estimation of human dose of staphylococcal enterotoxin A from a large outbreak of staphylococcal food poisoning involving chocolate milk. Int. J. Food Microbiol. 1988, 7, 311–316. [Google Scholar] [CrossRef]
- Harbrecht, D.F.; Bergdoll, M. Staphylococcal enterotoxin B production in hard-boiled eggs. J. Food Sci. 1980, 45, 307–309. [Google Scholar] [CrossRef]
- Guidi, F.; Duranti, A.; Gallina, S.; Nia, Y.; Petruzzelli, A.; Romano, A.; Travaglini, V.; Olivastri, A.; Calvaresi, V.; Decastelli, L.; et al. Characterization of A Staphylococcal Food Poisoning Outbreak in A Workplace Canteen during the Post-Earthquake Reconstruction of Central Italy. Toxins 2018, 10, 523. [Google Scholar] [CrossRef] [Green Version]
- Lis, E.; Podkowik, M.; Schubert, J.; Bystroń, J.; Stefaniak, T.; Bania, J. Production of Staphylococcal Enterotoxin R by Staphylococcus aureus Strains. Foodborne Pathog. Dis. 2012, 9, 762–766. [Google Scholar] [CrossRef]
- Hait, J.M.; Bennett, R.W.; Monday, S.R. Staphylococcal enterotoxin type r pseudogene presence in Staphylococcus aureus reference and outbreak strains. J. AOAC Int. 2018, 101, 216–220. [Google Scholar] [CrossRef]
- Lis, E.; Podkowik, M.; Bystroń, J.; Stefaniak, T.; Bania, J. Temporal expression of staphylococcal enterotoxin H in comparison with accessory gene regulator–dependent and –independent enterotoxins. J. Food Prot. 2012, 75, 238–244. [Google Scholar] [CrossRef]
- Jones, T.F.; Kellum, M.E.; Porter, S.S.; Bell, M.; Schaffner, W. An outbreak of community-acquired foodborne illness caused by methicillin-resistant Staphylococcus aureus. Emerg. Infect. Dis. 2002, 8, 82–84. [Google Scholar] [CrossRef]
- Peeters, L.E.J.; Argudín, M.A.; Azadikhah, S.; Butaye, P. Antimicrobial resistance and population structure of Staphylococcus aureus recovered from pigs farms. Vet. Microbiol. 2015, 180, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Aung, K.T.; Hsu, L.Y.; Koh, T.H.; Hapuarachchi, H.C.; Chau, M.L.; Gutiérrez, R.A.; Ng, L.C. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in retail food in Singapore. Antimicrob. Resist. Infect. Control. 2017, 6, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Shen, Y.; Dong, M. Distribution, polymorphism and temporal expression of egc in Staphylococcus aureus isolates from various foods in China. Food Control 2013, 29, 279–285. [Google Scholar] [CrossRef]
- Fischer, A.J.; Kilgore, S.H.; Singh, S.B.; Allen, P.D.; Hansen, A.R.; Limoli, D.H.; Schlievert, P.M. High prevalence of Staphylococcus aureus enterotoxin gene cluster superantigens in cystic fibrosis clinical isolates. Genes 2019, 10, 1036. [Google Scholar] [CrossRef] [Green Version]
- Grema, H.A.; Geidam, Y.A.; Gadzama, G.B.; Ameh, J.A.; Suleiman, A. Methicillin resistant Staphylococcus aureus (MRSA): A review. Adv. Anim. Vet. Sci. 2015, 3, 79–98. [Google Scholar] [CrossRef] [Green Version]
- Pekana, A.; Green, E. Antimicrobial resistance profiles of Staphylococcus aureus isolated from meat carcasses and bovine milk in abattoirs and dairy farms of the Eastern Cape, South Africa. Int. J. Environ. Res. Public Health 2018, 15, 2223. [Google Scholar] [CrossRef] [Green Version]
- Okorie-Kanu, O.J.; Anyanwu, M.U.; Ezenduka, E.V.; Mgbeahuruike, A.C.; Thapaliya, D.; Gerbig, G.; Ugwuijem, E.E.; Okorie-Kanu, C.O.; Agbowo, P.; Olorunleke, S.; et al. Molecular epidemiology, genetic diversity and antimicrobial resistance of Staphylococcus aureus isolated from chicken and pig carcasses, and carcass handlers. PLoS ONE 2020, 15, e0232913. [Google Scholar] [CrossRef]
- Sadat, A.; Shata, R.R.; Farag, A.M.M.; Ramadan, H.; Alkhedaide, A.; Soliman, M.M.; Elbadawy, M.; Abugomaa, A.; Awad, A. Prevalence and characterization of PVL-positive Staphylococcus aureus isolated from raw cow’s milk. Toxins 2022, 14, 97. [Google Scholar] [CrossRef]
- Chao, G.; Zhang, X.; Zhang, X.; Huang, Y.; Xu, L.; Zhou, L.; Yang, W.; Jiang, Y.; Xue, F.; Wu, Y. Phenotypic and genotypic characterization of methicillin-resistant Staphylococcus aureus (MRSA) and methicillin-susceptible Staphylococcus aureus (MSSA) from different sources in China. Foodborne Pathog. Dis. 2013, 10, 214–221. [Google Scholar] [CrossRef]
- Ge, B.; Mukherjee, S.; Hsu, C.H.; Davis, J.A.; Tran, T.T.T.; Yang, Q.; Abbott, J.W.; Ayers, S.L.; Young, S.R.; Crarey, E.T.; et al. MRSA and multidrug-resistant Staphylococcus aureus in U.S. retail meats, 2010–2011. Food Microbiol. 2017, 62, 289–297. [Google Scholar] [CrossRef]
- Drougka, E.; Foka, A.; Giormezis, N.; Sergelidis, D.; Militsopoulou, M.; Jelastopulu, E.; Komodromos, D.; Sarrou, S.; Anastassiou, E.D.; Petinaki, E.; et al. Multidrug-resistant enterotoxigenic Staphylococcus aureus lineages isolated from animals, their carcasses, the personnel, and the environment of an abattoir in Greece. J. Food Process. Preserv. 2019, 43, e13961. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; 22nd Informational Supplement, CLSI document M100-S22; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Pereira, L.A.; Harnett, G.B.; Hodge, M.M.; Cattell, J.A.; Speers, D.J. Real-time PCR assay for detection of blaZ genes in Staphylococcus aureus clinical isolates. J. Clin. Microbiol. 2014, 52, 1259–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, S.S.; Doern, G.V.; Heilmann, K.P.; Miner, S.; Tendolkar, S.; Riahi, F.; Diekema, D.J. Detection and prevalence of penicillin-susceptible Staphylococcus aureus in the United States in 2013. J. Clin. Microbiol. 2016, 54, 812–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Berger-Bächi, B.; Coombs, G.; Holmes, A.; Kay, I.; Kearns, A.; Linde, H.-J.; O’Brien, F.; Slickers, P.; Ehricht, R. Comparative genomics and DNA array-based genotyping of pandemic Staphylococcus aureus strains encoding Panton-Valentine leukocidin. Clin. Microbiol. Infect. 2007, 13, 236–249. [Google Scholar] [CrossRef] [Green Version]
- Arêde, P.; Ministro, J.; Oliveira, D.C. Redefining the role of the β-lactamase locus in methicillin-resistant Staphylococcus aureus: β-lactamase regulators disrupt the MecI-mediated strong repression on mecA and optimize the phenotypic expression of resistance in strains with constitutive mecA expression. Antimicrob. Agents Chemother. 2013, 57, 3037–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wysocka, M.; Monteiro, T.; de Pina, C.; Gonçalves, D.; de Pina, S.; Ludgero-Correia, A.; Moreno, J.; Zamudio, R.; Almebairik, N.; Gray, L.J.; et al. Whole-genome analysis uncovers loss of blaZ associated with carriage isolates belonging to methicillin-resistant Staphylococcus aureus (MRSA) clone ST5-VI in Cape Verde. J. Glob. Antimicrob. Resist. 2021, 26, 77–83. [Google Scholar] [CrossRef]
- Sabat, A.J.; Pournaras, S.; Akkerboom, V.; Tsakris, A.; Grundmann, H.; Friedrich, A.W. Whole-genome analysis of an oxacillin-susceptible CC80 mecA-positive Staphylococcus aureus clinical isolate: Insights into the mechanisms of cryptic methicillin resistance. J. Antimicrob. Chemother. 2015, 70, 2956–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mairi, A.; Touati, A.; Lavigne, J.P. Methicillin-resistant Staphylococcus aureus ST80 clone: A systematic review. Toxins 2020, 12, 119. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wu, C.; Wang, X.; Meng, J. Prevalence and characterization of methicillin susceptible Staphylococcus aureus ST398 isolates from retail foods. Int. J. Food Microbiol. 2015, 196, 94–97. [Google Scholar] [CrossRef]
- Naorem, R.S.; Blom, J.; Fekete, C. Genome-wide comparison of four MRSA clinical isolates from Germany and Hungary. PeerJ 2021, 9, e10185. [Google Scholar] [CrossRef]
- Bruce, S.A.; Smith, J.T.; Mydosh, J.L.; Ball, J.; Needle, D.B.; Gibson, R.; Andam, C.P. Shared antibiotic resistance and virulence genes in Staphylococcus aureus from diverse animal hosts. Sci. Rep. 2022, 12, 4413. [Google Scholar] [CrossRef] [PubMed]
- Muneeb, K.H.; Sudha, S.; Sivaraman, G.K.; Ojha, R.; Mendem, S.K.; Murugesan, D.; Raisen, C.L.; Shome, B.; Holmes, M. Whole-genome sequence analysis of Staphylococcus aureus from retail fish acknowledged the incidence of highly virulent ST672-MRSA-IVa/t1309, an emerging Indian clone, in Assam, India. Environ. Microbiol. Rep. 2022, 14, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Chairat, S.; Gharsa, H.; Lozano, C.; Gómez-Sanz, E.; Gómez, P.; Zarazaga, M.; Boudabous, A.; Torres, C.; Ben Slama, K. Characterization of Staphylococcus aureus from raw meat samples in Tunisia: Detection of clonal lineage ST398 from the African continent. Foodborne Pathog. Dis. 2015, 12, 686–692. [Google Scholar] [CrossRef] [PubMed]
- Omwenga, I.; Aboge, G.O.; Mitema, E.S.; Obiero, G.; Ngaywa, C.; Ngwili, N.; Wamwere, G.; Wainaina, M.; Bett, B. Antimicrobial usage and detection of multidrug-resistant Staphylococcus aureus, including methicillin-resistant strains in raw milk of livestock from Northern Kenya. Microb. Drug Resist. 2021, 27, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Amoako, D.G.; Somboro, A.M.; Abia, A.L.K.; Allam, M.; Ismail, A.; Bester, L.; Essack, S.Y. Genomic analysis of methicillin-resistant Staphylococcus aureus isolated from poultry and occupational farm workers in Umgungundlovu District, South Africa. Sci. Total Environ. 2019, 670, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.; Hartleib, J.; Hussain, M.S.; Peters, G. The multifunctional Staphylococcus aureus autolysin aaa mediates adherence to immobilized fibrinogen and fibronectin. Infect. Immun. 2005, 73, 4793–4802. [Google Scholar] [CrossRef] [Green Version]
- Menzies, B.E. The role of fibronectin binding proteins in the pathogenesis of Staphylococcus aureus infections. Curr. Opin. Infect. Dis. 2003, 16, 225–229. [Google Scholar] [CrossRef]
- Bjerketorp, J.; Jacobsson, K.; Frykberg, L. The von Willebrand factor-binding protein (vWbp) of Staphylococcus aureus is a coagulase. FEMS Microbiol Lett. 2004, 234, 309–314. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Rohmer, C.; Wolz, C. The role of hlb-converting bacteriophages in Staphylococcus aureus host adaption. Microb. Physiol. 2021, 31, 109–122. [Google Scholar] [CrossRef]
- Hayashida, A.; Bartlett, A.H.; Foster, T.J.; Park, P.W. Staphylococcus aureus beta-toxin induces lung injury through syndecan-1. Am. J. Pathol. 2009, 174, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, Y.; Baba, T.; Sekine, M.; Fukuda, M.; Hiramatsu, K. Beta-hemolysin promotes skin colonization by Staphylococcus aureus. J. Bacteriol. 2013, 195, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Rasigade, J.P.; Laurent, F.; Lina, G.; Meugnier, H.; Bes, M.; Vandenesch, F.; Etienne, J.; Tristan, A. Global distribution and evolution of panton-valentine leukocidin-positive methicillin-susceptible Staphylococcus aureus, 1981–2007. J. Infect. Dis. 2010, 201, 1589–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle, D.L., Jr.; Paclibare, P.A.; Cabrera, E.C.; Rivera, W.L. Molecular and phenotypic characterization of methicillin-resistant Staphylococcus aureus isolates from a tertiary hospital in the Philippines. Trop. Med. Health 2016, 44, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC). Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar] [CrossRef] [Green Version]
- Uehara, Y. Current status of Staphylococcal Cassette Chromosome mec (SCCmec). Antibiotics 2022, 11, 86. [Google Scholar] [CrossRef]
- Mairi, A.; Touati, A.; Pantel, A.; Yahiaoui Martinez, A.; Ahmim, M.; Sotto, A.; Dunyach-Remy, C.; Lavigne, J.P. First Report of CC5-MRSA-IV-SCCfus “Maltese Clone” in Bat Guano. Microorganisms 2021, 9, 2264. [Google Scholar] [CrossRef]
- Aouati, H.; Hadjadj, L.; Aouati, F.; Agabou, A.; Ben Khedher, M.; Bousseboua, H.; Bentchouala, C.; Rolain, J.M.; Diene, S.M. Emergence of methicillin-resistant Staphylococcus aureus ST239/241 SCCmec-III mercury in Eastern Algeria. Pathogens 2021, 10, 1503. [Google Scholar] [CrossRef]
- Fusco, V.; Quero, G.M.; Morea, M.; Blaiotta, G.; Visconti, A. Rapid and reliable identification of Staphylococcus aureus harbouring the enterotoxin gene cluster (egc) and quantitative detection in raw milk by real time PCR. Int. J. Food Microbiol. 2011, 144, 528–537. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2021, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-R, L.M.; Gunturu, S.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. The Microbial Genomes Atlas (MiGA) webserver: Taxonomic and gene diversity analysis of Archaea and Bacteria at the whole genome level. Nucleic Acids Res. 2018, 46, W282–W288. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- BLAST. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 15 April 2022).
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. Peer J. 2016, 4, e1900v1. [Google Scholar] [CrossRef]
- BV-BRC: Bacterial And Viral Bioinformatics Resource Center. Available online: https://www.bv-brc.org/ (accessed on 10 May 2022).
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- MLST. Available online: https://cge.cbs.dtu.dk/services/MLST (accessed on 8 January 2022).
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total genome sequenced bacteria. Clin. Micobiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- spaTyper. Available online: https://cge.cbs.dtu.dk/services/spaTyper (accessed on 8 January 2022).
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarløv, J.O.; Boye, K.; Larsen, A.R.; et al. Comparing whole-genome sequencing with Sanger sequencing for spa typing of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.J.; Boisvert, S.; Brettin, T.; Kenyon, R.W.; Mao, C.; Olson, R.; Overbeek, R.; Santerre, J.; Shukla, M.; Wattam, A.R.; et al. Antimicrobial Resistance Prediction in PATRIC and RAST. Sci Rep. 2016, 6, 27930. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, 687–692. [Google Scholar] [CrossRef] [PubMed]
- VFDB. Available online: http://www.mgc.ac.cn/VFs/ (accessed on 8 February 2022).
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Merda, D.; Felten, A.; Vingadassalon, N.; Denayer, S.; Titouche, Y.; Decastelli, L.; Hickey, B.; Kourtis, C.; Daskalov, H.; Mistou, M.Y.; et al. NAuRA: Genomic Tool to Identify Staphylococcal Enterotoxins in Staphylococcus aureus Strains Responsible for FoodBorne Outbreaks. Front. Microbiol. 2020, 11, 1483. [Google Scholar] [CrossRef] [PubMed]
- SCCmecFinder. Available online: https://cge.cbs.dtu.dk/services/SCCmecFinder-1.2 (accessed on 8 January 2022).
- Kaya, H.; Hasman, H.; Larsen, J.; Stegger, M.; Johannesen, T.B.; Allesøe, R.L.; Lemvigh, C.K.; Aarestrup, F.M.; Lund, O.; Larsen, A.R. SCCmecFinder, a web-based tool for typing of Staphylococcal Cassette Chromosome mec in Staphylococcus aureus using whole-genome sequence data. mSphere 2018, 3, e00612-17. [Google Scholar] [CrossRef] [Green Version]
- Schubert, J.; Podkowik, M.; Bystroń, J.; Bania, J. Production of staphylococcal enterotoxins in microbial broth and milk by Staphylococcus aureus strains harboring seh gene. Int. J. Food Microbiol. 2016, 235, 36–45. [Google Scholar] [CrossRef]
- Schubert, J.; Podkowik, M.; Bystroń, J.; Bania, J. Production of staphylococcal enterotoxins D and R in milk and meat juice by Staphylococcus aureus strains. Foodborne Pathog. Dis. 2017, 14, 223–230. [Google Scholar] [CrossRef]
- Valihrach, L.; Alibayov, B.; Zdenkova, K.; Demnerova, K. Expression and production of staphylococcal enterotoxin C is substantially reduced in milk. Food Microbiol. 2014, 44, 54–59. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MSSA or MRSA | Origin | ST | se and sel Genes | mecA (SCCmec-Type) | tst | lukF-PV and lukS-PV |
|---|---|---|---|---|---|---|---|
| SA01 | MSSA | Potato in sauce | 15 | selwa, selxh | - | - | - |
| SA02 | MSSA | Cooked meat | 5 | egc1 (seg, sei, sem, sen, seo, ψent1-ψent2 *), selw *,b, selx1 | - | - | - |
| SA04 | MSSA | Cooked meat | 1 | sea1, seh, sek, seq, selwa, selx5 | - | - | - |
| SA07 | MSSA | Cooked meat | 7199 k | selwa, selxh | - | - | - |
| SA08 | MSSA | Cooked meat | 25 | egc5 (seg, sei, sem, sen, seo, selu2), selwd, selx9 i | - | - | - |
| SA10 | MSSA | Cooked meat | 101 | selwe, selxh | - | - | - |
| SA18 | MSSA | Pizza | 97 | sebv1, selw *,f, selxbov2 | - | - | - |
| SA20 | MSSA | Cooked meat | 8 | sed *,j, selj, ser, selw c, selx2 | - | - | - |
| SA24 | MSSA | Lentil soup | 7200 k | egc5 (seg, sei, sem, sen, seo, selu2), selw *,f, selx5 | - | - | - |
| SA46 | MRSA | Braised beef | 80 | seh *, selw g, selx h, sey | +(SCCmec-IV) | - | + |
| SA51 | MSSA | Fermented milk | 5 | sea2 i, egc1 (seg, sei, sem, sen, seo, ψent1-ψent2 *), selw *,b, selx1, sel30 | - | - | - |
| SA82 | MSSA | Sautéed beef with potato | 22 | egc5 (seg, sei, sem, sen, seo, selu2), selwd, selx6 | - | + | - |
| Strain | Resistance Profile a | Genetic Determinants |
|---|---|---|
| SA01 | P | blaZ |
| SA02 | OF, ERY, L, TE | gyrA-S84L, ermB, lmrS, tet(L), tet(38) |
| SA04 | P | blaZ |
| SA07 | P | blaZ |
| SA08 | P | blaZ |
| SA10 | P | blaZ |
| SA18 | P, KAN, TE | blaZ, aph(3′)-IIIa, ant(6)-I, tet(L), tet(38) |
| SA20 | P | blaZ |
| SA24 | P | blaZ |
| SA46 | P, OXA, FOX, KAN | mecA, aph(3′)-IIIa, ant(6)-I |
| SA51 | - b | blaZ |
| SA82 | P | blaZ |
| Strain | 0 h | 24 h | 48 h | |||
|---|---|---|---|---|---|---|
| Log cfu/mL | pH | Log cfu/mL | pH | Log cfu/mL | pH | |
| SA04 | 6.77 ± 0.58 | 7.61 | 9.22 ± 0.03 | 6.86 | 9.22 ± 0.12 | 7.53 |
| SA18 | 6.91 ± 0.07 | 7.61 | 9.34 ± 0.23 | 6.97 | 9.44 ± 0.28 | 7.57 |
| SA20 | 6.91 ± 0.29 | 7.61 | 8.97 ± 0.09 | 6.71 | 9.08 ± 0.13 | 7.29 |
| SA46 | 6.39 ± 0.26 | 7.61 | 9.29 ± 0.08 | 7.05 | 9.26 ± 0.08 | 7.61 |
| SA51 | 6.38 ± 0.28 | 7.61 | 9.15 ± 0.11 | 6.81 | 9.16 ± 0.07 | 7.23 |
| Strain | SEA | SEB | SED | SEH | SER | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h | |
| SA04 | 156.04 ± 36.58 | 345.02 ± 62.67 | - | - | - | - | 221.76 ± 38.15 | 239.48 ± 84.83 | - | - |
| SA18 | - | - | 2871.28 ± 811.09 | 14,739.17 ± 5077.70 | - | - | - | - | - | - |
| SA20 | - | - | - | - | 322.70 ± 41.17 | 398.94 ± 64.79 | - | - | 36,720.10 ± 6272.32 | 63,176.06 ± 14,422.93 |
| SA46 | - | - | - | - | - | - | 1.76 ± 2.26 | N.D. | - | - |
| SA51 | 64.54 ± 9.29 | 74.91 ± 7.36 | - | - | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanelli, F.; Chieffi, D.; Cho, G.-S.; Schubert, J.; Mekhloufi, O.A.; Bania, J.; Franz, C.M.A.P.; Fusco, V. First Genome-Based Characterisation and Staphylococcal Enterotoxin Production Ability of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus Strains Isolated from Ready-to-Eat Foods in Algiers (Algeria). Toxins 2022, 14, 731. https://doi.org/10.3390/toxins14110731
Fanelli F, Chieffi D, Cho G-S, Schubert J, Mekhloufi OA, Bania J, Franz CMAP, Fusco V. First Genome-Based Characterisation and Staphylococcal Enterotoxin Production Ability of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus Strains Isolated from Ready-to-Eat Foods in Algiers (Algeria). Toxins. 2022; 14(11):731. https://doi.org/10.3390/toxins14110731
Chicago/Turabian StyleFanelli, Francesca, Daniele Chieffi, Gyu-Sung Cho, Justyna Schubert, Omar Amine Mekhloufi, Jacek Bania, Charles M. A. P. Franz, and Vincenzina Fusco. 2022. "First Genome-Based Characterisation and Staphylococcal Enterotoxin Production Ability of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus Strains Isolated from Ready-to-Eat Foods in Algiers (Algeria)" Toxins 14, no. 11: 731. https://doi.org/10.3390/toxins14110731