Evaluation of Toxicant-Associated Fatty Liver Disease and Liver Neoplastic Progress in Sprague-Dawley Rats Treated with Low Doses of Aflatoxin B1 Alone or in Combination with Extremely Low Frequency Electromagnetic Fields

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
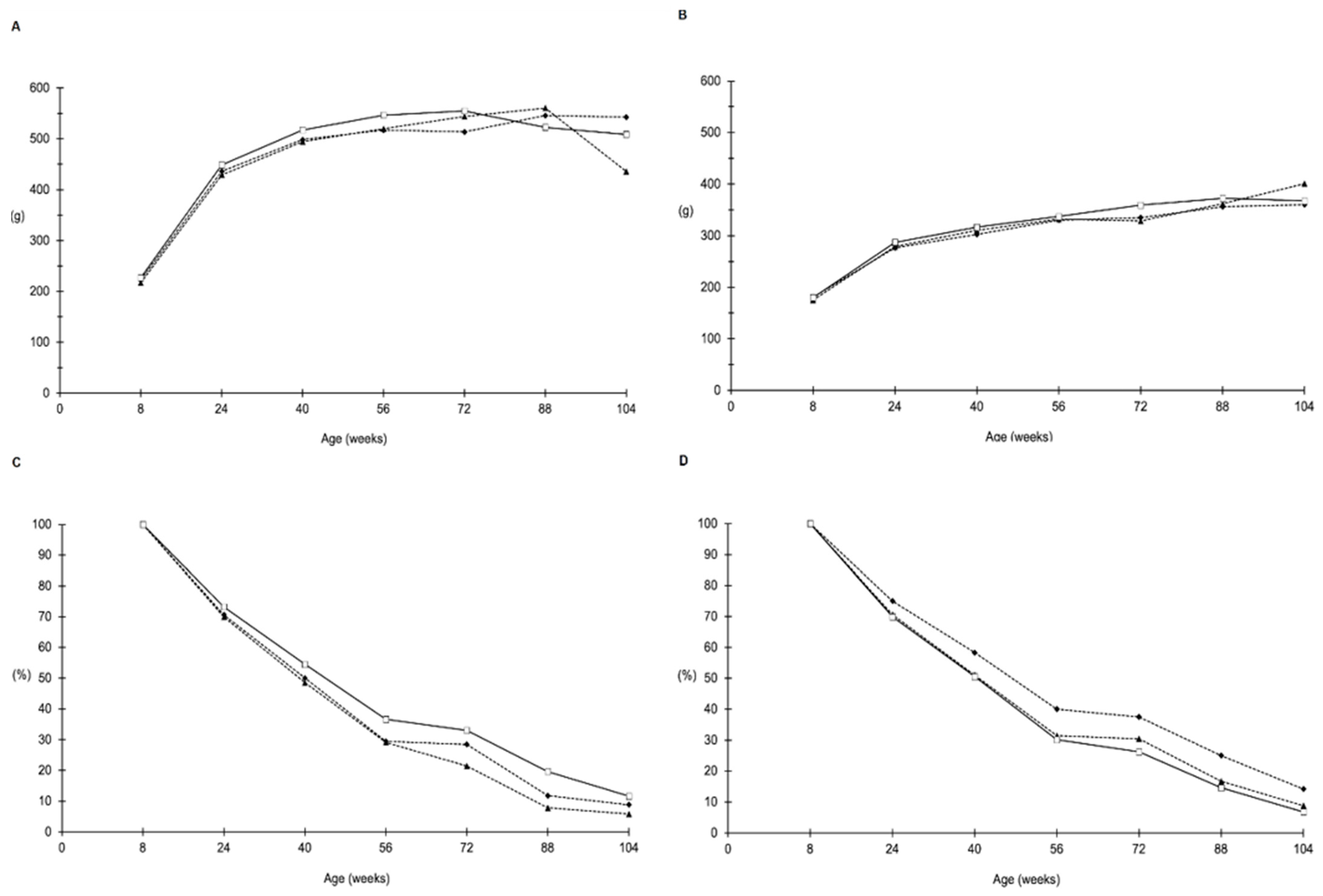
2.1. Food and Water Consumption, Body Weight and Survival
2.2. Necropsy
2.3. Synergistic Effects of AFB1 and ELFEMF in Male Rats
2.4. Synergistic Effects of AFB1 and ELFEMF in Female Rats
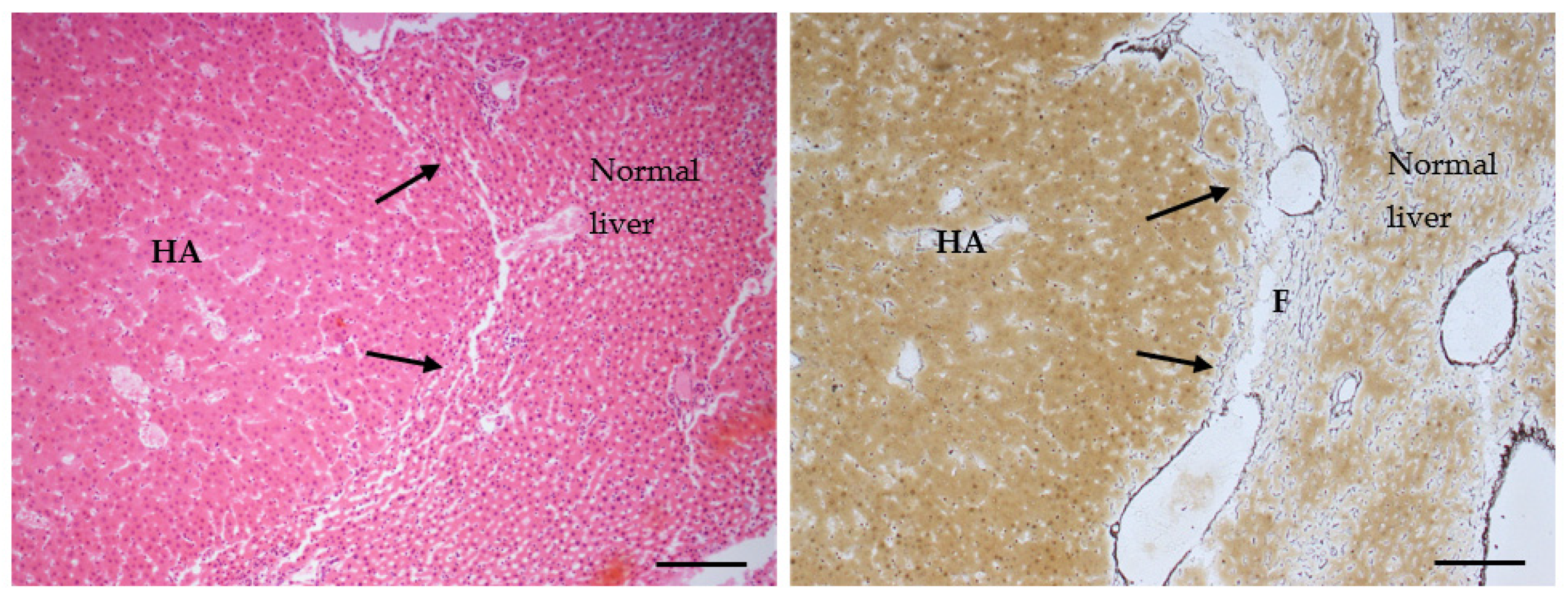
2.5. Pre-Neoplastic, Neoplastic and TAFLD Lesions in Liver of AFB1 Treated Male Rats
2.6. Pre-Neoplastic, Neoplastic and TAFLD Lesions in Liver of AFB1 Treated Female Rats
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Treatment and Conduct of the Experiment
5.2. Diet
5.3. Experimental Animals
5.4. Histopathology
5.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Farrell, G.; Schattenberg, J.M.; Leclercq, I.; Yeh, M.M.; Goldin, R.; Teoh, N.; Schuppan, D. Mouse Models of Nonalcoholic Steatohepatitis: Toward Optimization of Their Relevance to Human Nonalcoholic Steatohepatitis. Hepatology 2018, 69, 2241–2257. [Google Scholar] [CrossRef] [Green Version]
- Cave, M.; Falkner, K.C.; Ray, M.; Joshi-Barve, S.; Brock, G.; Khan, R.; Homme, M.B.; McClain, C.J. Toxicant-associated steatohepatitis in vinyl chloride workers. Hepatology 2009, 51, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Carithers, R.; McClain, C. Sleisenger & Fordtran’s Gastrointestinal and Liver Disease; Elsevier Inc.: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Clark, J.M. The epidemiology of nonalcoholic fatty liver disease in adults. J. Clin. Gastroenterol. 2006, 40, S5–S10. [Google Scholar] [CrossRef]
- Maltoni, C.; Lefemine, G.; Ciliberti, A.; Cotti, G.; Carretti, D. Carcinogenicity bioassays of vinyl chloride monomer: A model of risk assessment on an experimental basis. Environ. Health Perspect. 1981, 41, 3–29. [Google Scholar] [CrossRef]
- Wahlang, B.; Beier, J.I.; Clair, H.B.; Bellis-Jones, H.J.; Falkner, K.C.; McClain, C.J.; Cave, M.C. Toxicant-associated Steatohepatitis. Toxicol. Pathol. 2012, 41, 343–360. [Google Scholar] [CrossRef] [Green Version]
- Dey, D.K.; Kang, S.C. Aflatoxin B1 induces reactive oxygen species-dependent caspase-mediated apoptosis in normal human cells, inhibits Allium cepa root cell division, and triggers inflammatory response in zebrafish larvae. Sci. Total Environ. 2020, 737, 139704. [Google Scholar] [CrossRef]
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant Pathol. 2007, 8, 713–722. [Google Scholar] [CrossRef]
- IARC. Working Group on the Evaluation of Carcinogenic Risks to Humans International Agency for Research on Cancer—Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene; World Health Organization: Geneva, Switzerland, 2002; Volume 82. [Google Scholar]
- Kucukcakan, B.; Hayrulai-Musliu, Z. Challenging Role of Dietary Aflatoxin B1 Exposure and Hepatitis B Infection on Risk of Hepatocellular Carcinoma. Open Access Maced. J. Med Sci. 2015, 3, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Wild, C.P.; Miller, J.D.; Groopman, J.D. Mycotoxin Control in Low-And Middle-Income Countries. 2015. Available online: https://publications.iarc.fr/_publications/media/download/4264/f4032be5bbc99862a91d2677e6cc902a08faf7fa.pdf (accessed on 10 April 2022).
- Efsa Panel On Contaminants In The Food Chain (Contam); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; Del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.; et al. Risk assessment of aflatoxins in food. EFSA J. 2020, 18, e06040. [Google Scholar] [CrossRef]
- Soffritti, M.; McCONNELL, E.E. Liver Foci Formation during Aflatoxin B1Carcinogenesis in the Rat. Ann. N. Y. Acad. Sci. 1988, 534, 531–540. [Google Scholar] [CrossRef]
- Bua, L.; Tibaldi, E.; Falcioni, L.; Lauriola, M.; De Angelis, L.; Gnudi, F.; Manservigi, M.; Manservisi, F.; Manzoli, I.; Menghetti, I.; et al. Results of lifespan exposure to continuous and intermittent extremely low frequency electromagnetic fields (ELFEMF) administered alone to Sprague Dawley rats. Environ. Res. 2018, 164, 271–279. [Google Scholar] [CrossRef]
- Soffritti, M.; Tibaldi, E.; Padovani, M.; Hoel, D.G.; Giuliani, L.; Bua, L.; Lauriola, M.; Falcioni, L.; Manservigi, M.; Manservisi, F.; et al. Synergism between sinusoidal-50 Hz magnetic field and formaldehyde in triggering carcinogenic effects in male Sprague-Dawley rats. Am. J. Ind. Med. 2016, 59, 509–521. [Google Scholar] [CrossRef]
- Soffritti, M.; Tibaldi, E.; Padovani, M.; Hoel, D.G.; Giuliani, L.; Bua, L.; Lauriola, M.; Falcioni, L.; Manservigi, M.; Manservisi, F.; et al. Life-span exposure to sinusoidal-50 Hz magnetic field and acute low-dose γ radiation induce carcinogenic effects in Sprague-Dawley rats. Int. J. Radiat. Biol. 2016, 92, 202–214. [Google Scholar] [CrossRef]
- Zhang, L.; Ye, Y.; An, Y.; Tian, Y.; Wang, Y.; Tang, H. Systems Responses of Rats to Aflatoxin B1 Exposure Revealed with Metabonomic Changes in Multiple Biological Matrices. J. Proteome Res. 2010, 10, 614–623. [Google Scholar] [CrossRef]
- Towner, R.A.; Hashimoto, H.; Summers, P.M. Non-invasive in vivo magnetic resonance imaging assessment of acute aflatoxin B1 hepatotoxicity in rats. Biochim. Biophys. Acta (BBA) Gen. Subj. 2000, 1475, 314–320. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Li, X.; Wang, Z. Role of xenobiotics in the induction and progression of fatty liver disease. Toxicol. Res. 2018, 7, 664–680. [Google Scholar] [CrossRef] [Green Version]
- Treviño, L.S.; Katz, T.A. Endocrine Disruptors and Developmental Origins of Nonalcoholic Fatty Liver Disease. Endocrinology 2017, 159, 20–31. [Google Scholar] [CrossRef]
- Storvik, M.; Huuskonen, P.; Kyllönen, T.; Lehtonen, S.; El-Nezami, H.; Auriola, S.; Pasanen, M. Aflatoxin B1–a potential endocrine disruptor–up-regulates CYP19A1 in JEG-3 cells. Toxicol. Lett. 2011, 202, 161–167. [Google Scholar] [CrossRef]
- Huuskonen, P.; Myllynen, P.; Storvik, M.; Pasanen, M. The effects of aflatoxin B1 on transporters and steroid metabolizing enzymes in JEG-3 cells. Toxicol. Lett. 2013, 218, 200–206. [Google Scholar] [CrossRef]
- Yip, K.Y.; Wan, M.L.Y.; Wong, A.S.T.; Korach, K.S.; El-Nezami, H. Combined low-dose zearalenone and aflatoxin B1 on cell growth and cell-cycle progression in breast cancer MCF-7 cells. Toxicol. Lett. 2017, 281, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Shi, W.; Lv, P.; Meng, W.; Mao, G.; Gong, C.; Chen, Y.; Wei, Y.; He, X.; Zhao, J.; et al. Critical role of caveolin-1 in aflatoxin B1-induced hepatotoxicity via the regulation of oxidation and autophagy. Cell Death Dis. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, A.T.; Hirooka, E.Y.; e Silva, P.L.A.; Bracarense, A.P.F.R.L.; Flaiban, K.K.M.D.C.; Akagi, C.Y.; Kawamura, O.; da Costa, M.C.; Itano, E.N. Impact of a Single Oral Acute Dose of Aflatoxin B1 on Liver Function/Cytokines and the Lymphoproliferative Response in C57Bl/6 Mice. Toxins 2017, 9, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, D.K.; Chang, S.N.; Kang, S.C. The inflammation response and risk associated with aflatoxin B1 contamination was minimized by insect peptide CopA3 treatment and act towards the beneficial health outcomes. Environ. Pollut. 2021, 268, 115713. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P.; James, O.F. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef]
- Yilmaz, Y. Is non-alcoholic fatty liver disease a spectrum, or are steatosis and non-alcoholic steatohepatitis distinct conditions? Aliment. Pharmacol. Therap. 2012, 36, 815–823. [Google Scholar] [CrossRef]
- Takaki, A.; Kawai, D.; Yamamoto, K. Multiple Hits, Including Oxidative Stress, as Pathogenesis and Treatment Target in Non-Alcoholic Steatohepatitis (NASH). Int. J. Mol. Sci. 2013, 14, 20704–20728. [Google Scholar] [CrossRef] [Green Version]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Falkner, K.C.; Hill, B.G.; Sansbury, B.; McClain, C.J.; Cave, M.C. Mitochondrial toxicity of chloroacetaldehyde in HepG2 cells. In Proceedings of the Hepatology of the 61st Annual Meeting of the American Association for the Study of Liver Diseases: The Liver Meeting, Boston, MA, USA, 29 October–2 November 2010; Volume 52, pp. 455A–456A. [Google Scholar]
- Bannasch, P.; Haertel, T.; Su, Q. Significance of hepatic preneoplasia in risk identification and early detection of neoplasia. Toxicol. Pathol. 2003, 31, 134–139. [Google Scholar]
- Qian, G.; Wang, F.; Tang, L.; Massey, M.E.; Mitchell, N.J.; Su, J.; Williams, J.H.; Phillips, T.D.; Wang, J.-S. Integrative Toxicopathological Evaluation of Aflatoxin B1 Exposure in F344 Rats. Toxicol. Pathol. 2013, 41, 1093–1105. [Google Scholar] [CrossRef] [Green Version]
- Kamdem, L.K.; Meineke, I.; Gödtel-Armbrust, U.; Brockmöller, J.; Wojnowski, L. Dominant Contribution of P450 3A4 to the Hepatic Carcinogenic Activation of Aflatoxin B1. Chem. Res. Toxicol. 2006, 19, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Raney, K.D.; Shimada, T.; Kim, D.H.; Groopman, J.D.; Harris, T.M.; Guengerich, F.P. Oxidation of aflatoxins and sterigmatocystin by human liver microsomes: Significance of aflatoxin Q1 as a detoxication product of aflatoxin B1. Chem. Res. Toxicol. 1992, 5, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y. The Toxic Effects of Aflatoxin B1: An update. In Aflatoxin B1 Occurrence, Detection and Toxicological Effects; IntechOpen: London, UK, 2019. [Google Scholar]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological Properties and Their Involvement in Cancer Development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef] [Green Version]
- Chappell, G.; Pogribny, I.P.; Guyton, K.Z.; Rusyn, I. Epigenetic alterations induced by genotoxic occupational and environmental human chemical carcinogens: A systematic literature review. Mutat. Res. Mutat. Res. 2016, 768, 27–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, J.; Esteller, M. Cancer epigenomics: Beyond genomics. Curr. Opin. Genet. Dev. 2012, 22, 50–55. [Google Scholar] [CrossRef]
- Thoolen, B.; Maronpot, R.R.; Harada, T.; Nyska, A.; Rousseaux, C.; Nolte, T.; Malarkey, D.E.; Kaufmann, W.; Küttler, K.; Deschl, U.; et al. Proliferative and Nonproliferative Lesions of the Rat and Mouse Hepatobiliary System. Toxicol. Pathol. 2010, 38, 5S–81S. [Google Scholar] [CrossRef]
- Heyens, L.J.M.; Busschots, D.; Koek, G.H.; Robaeys, G.; Francque, S. Liver Fibrosis in Non-alcoholic Fatty Liver Disease: From Liver Biopsy to Non-invasive Biomarkers in Diagnosis and Treatment. Front. Med. 2021, 8, 476. [Google Scholar] [CrossRef]
- Zhong, F.; Zhou, X.; Xu, J.; Gao, L. Rodent Models of Nonalcoholic Fatty Liver Disease. Digestion 2020, 101, 522–535. [Google Scholar] [CrossRef]
- Krishna, M. Role of special stains in diagnostic liver pathology. Clin. Liver Dis. 2013, 2, S8–S10. [Google Scholar] [CrossRef]
- OECD. Guidance Notes for Analysis and Evaluation of Chronic Toxicity and Carcinogenicity Studies. 2002. Available online: https://one.oecd.org/document/ENV/JM/MONO(2002)19/en/pdf (accessed on 10 April 2022).
- OECD. Draft Guidance Document on the Design and Conduct of Chronic Toxicity and Carcinogenicity Studies, Series on Testing and Assessment No. 116. 2009. Available online: https://www.oecd.org/chemicalsafety/testing/44076587.pdf (accessed on 10 April 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | M | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group (Dose) | Group I (Controls) | Group II (AFB1) | Group III (AFB1 + S 50 Hz ELFEMF) | ||||||
| No | 10 | 10 | 10 | ||||||
| n | % | n | % | p-Value II vs. Ctrl | n | % | p-Value III vs. Ctrl | p-Value III vs. II | |
| Inflammation | 0 | 0.0 | 2 | 20.0 | 0.474 | 0 | 0.0 | - | 0.474 |
| Steatosis | 1 | 10.0 | 3 | 30.0 | 0.582 | 4 | 40.0 | 0.303 | 1.000 |
| Fibrosis | 0 | 0.0 | 2 | 20.0 | 0.474 | 1 | 10.0 | 1.000 | 1.000 |
| Foci | 0 | 0.0 | 3 | 30.0 | 0.211 | 1 | 10.0 | 1.000 | 0.582 |
| Hepatocellular Adenoma | 0 | 0.0 | 0 | 0.0 | - | 0 | 0.0 | - | - |
| Hepatocellular carcinoma (HCC) | 0 | 0.0 | 0 | 0.0 | - | 0 | 0.0 | - | - |
| Sex | M | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group (Dose) | Group I (Controls) | Group II (AFB1) | Group III (AFB1 + S 50 Hz ELFEMF) | ||||||
| No | 31 | 18 | 17 | ||||||
| n | % | n | % | p-Value II vs. Ctrl | n | % | p-Value III vs. Ctrl | p-Value III vs. II | |
| Inflammation intraparenchymal | 4 | 12.9 | 4 | 22.2 | 0.443 | 5 | 29.4 | 0.247 | 0.711 |
| Inflammation total | 5 | 16.1 | 4 | 22.2 | 0.708 | 5 | 29.4 | 0.295 | 0.711 |
| Steatosis | 5 | 16.1 | 3 | 16.7 | 1.000 | 6 | 35.3 | 0.163 | 0.264 |
| Hepatocyte necrosis | 5 | 16.1 | 6 | 33.3 | 0.286 | 6 | 35.3 | 0.163 | 1.000 |
| Foci | 1 | 3.2 | 3 | 16.7 | 0.134 | 4 | 23.5 | 0.047 * | 0.691 |
| Hepatocellular Adenoma | 0 | 0.0 | 0 | 0.0 | - | 1 | 5.9 | 0.354 | 0.486 |
| Sex | M | |||||||
|---|---|---|---|---|---|---|---|---|
| Group (Dose) | BT1CEM Controls | Group I (Controls) | Group II + III (AFB1; S-50 Hz ELFEMF + AFB1) | |||||
| No | 500 | 31 | 35 | |||||
| n | % | n | % | n | % | p-Value II + III vs. BT1CEM Ctrl | p-Value II + III vs. I | |
| Inflammation intraparenchymal | 81 | 16.2 | 4 | 12.9 | 9 | 25.7 | 0.146 | 0.192 |
| Inflammation total | 89 | 17.8 | 5 | 16.1 | 9 | 25.7 | 0.258 | 0.382 |
| Steatosis | 58 | 11.6 | 5 | 16.1 | 9 | 25.7 | 0.015 # | 0.342 |
| Fibrosis | 6 | 1.2 | 0 | 0.0 | 2 | 5.7 | 0.091 | 0.494 |
| Hepatocyte hyperplasia regenerative | 3 | 0.6 | 0 | 0.0 | 1 | 2.9 | 0.134 | 1.000 |
| Hepatocyte hypertrophy focal | 4 | 0.8 | 0 | 0.0 | 2 | 5.7 | 0.053 | 0.494 |
| Oval cell hyperplasia | 10 | 2.0 | 0 | 0.0 | 1 | 2.9 | 0.528 | 1.000 |
| Hepatocyte necrosis | 91 | 18.2 | 5 | 16.1 | 12 | 34.3 | 0.020 # | 0.158 |
| Basophilic focus | 0 | 0.0 | 0 | 0.0 | 1 | 2.9 | 0.065 | 1.000 |
| Clear cells focus | 2 | 0.4 | 0 | 0.0 | 4 | 11.4 | 0.000 # | 0.116 |
| Eosinophilic focus | 4 | 0.8 | 1 | 3.2 | 4 | 11.4 | 0.001 # | 0.360 |
| Foci total | 6 | 1.2 | 1 | 3.2 | 7 | 20.0 | 0.000 # | 0.058 |
| Hepatocellular Adenoma | 2 | 0.4 | 0 | 0.0 | 1 | 2.9 | 0.060 | 1.000 |
| Sex | F | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group (Dose) | Group I (Controls) | Group II (AFB1) | Group III (AFB1 + S 50 Hz ELFEMF) | ||||||
| No | 10 | 10 | 10 | ||||||
| n | % | n | % | p-Value II vs. Ctrl | n | % | p-Value III vs. Ctrl | p-Value III vs. II | |
| Inflammation total | 1 | 10.0 | 6 | 60.0 | 0.057 | 4 | 40.0 | 0.303 | 0.656 |
| Steatosis total | 2 | 20.0 | 1 | 10.0 | 1.000 | 4 | 40.0 | 0.628 | 0.303 |
| Fibrosis | 0 | 0.0 | 0 | 0.0 | - | 2 | 20.0 | 0.474 | 0.474 |
| Foci total | 0 | 0.0 | 3 | 30.0 | 0.211 | 3 | 30.0 | 0.211 | 1.000 |
| Hepatocellular Adenoma | 0 | 0.0 | 0 | 0.0 | - | 2 | 20.0 | 0.474 | 0.474 |
| Hepatocellular Carcinoma (HCC) | 0 | 0.0 | 0 | 0.0 | - | 1 | 10.0 | 1.000 | 1.000 |
| Sex | F | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group (Dose) | Group I (Controls) | Group II (AFB1) | Group III (AFB1 + S 50 Hz ELFEMF) | ||||||
| No | 23 | 21 | 39 | ||||||
| n | % | n | % | p-Value II vs. Ctrl | n | % | p-Value III vs. Ctrl | p-Value III vs. II | |
| Inflammation intraparenchymal | 4 | 17.4 | 11 | 52.4 | 0.025 * | 10 | 25.6 | 0.541 | 0.038 ° |
| Inflammation total | 5 | 21.7 | 12 | 57.1 | 0.029 * | 12 | 44.4 | 0.561 | 0.047 ° |
| Steatosis total | 3 | 13.0 | 4 | 19.0 | 0.693 | 11 | 28.2 | 0.218 | 0.541 |
| Hepatocyte necrosis | 2 | 8.7 | 6 | 28.6 | 0.126 | 10 | 25.6 | 1.000 | 0.182 |
| Eosinophilic focus | 0 | 0.0 | 6 | 28.6 | 0.008 * | 8 | 20.5 | 0.021 * | 0.532 |
| Foci total | 0 | 0.0 | 6 | 28.6 | 0.008 * | 9 | 23.1 | 0.020 * | 0.757 |
| Hepatocellular Adenoma | 0 | 0.0 | 0 | 0.0 | - | 5 | 12.8 | 0.152 | 0.148 |
| Sex | F | |||||||
|---|---|---|---|---|---|---|---|---|
| Group (Dose) | BT1CEM Controls | Group I (Control) | Group II + III (AFB1; S 50 Hz ELFEMF + AFB1) | |||||
| No | 501 | 23 | 60 | |||||
| n | % | n | % | n | % | p-Value II + III vs. BT1CEM Ctrl | p-Value II + III vs. I | |
| Inflammation intraparenchymal | 178 | 35.6 | 4 | 17.4 | 21 | 35.0 | 0.935 | 0.181 |
| Inflammation total | 184 | 36.7 | 5 | 21.7 | 24 | 40.0 | 0.620 | 0.133 |
| Steatosis diffuse | 40 | 8.0 | 2 | 8.7 | 7 | 20.0 | 0.324 | 1.000 |
| Steatosis focal | 14 | 2.8 | 1 | 4.3 | 8 | 22.9 | 0.001 # | 0.433 |
| Steatosis total | 54 | 10.8 | 3 | 13.0 | 15 | 42.9 | 0.003 # | 0.373 |
| Fibrosis | 10 | 2.0 | 1 | 4.3 | 2 | 3.3 | 0.374 | 1.000 |
| Cholangiofibrosis focal | 15 | 3.0 | 1 | 4.3 | 5 | 8.3 | 0.052 | 1.000 |
| Cystic dilation biliary duct | 3 | 0.6 | 1 | 4.3 | 5 | 8.3 | 0.001 # | 1.000 |
| Hepatocyte hyperplasia regenerative | 2 | 0.4 | 0 | 0.0 | 3 | 5.0 | 0.010 # | 0.557 |
| Hepatocyte hypertrophy focal | 2 | 0.4 | 0 | 0.0 | 8 | 13.3 | 0.000 # | 0.099 |
| Oval cell hyperplasia | 12 | 2.4 | 0 | 0.0 | 8 | 13.3 | 0.000 # | 0.099 |
| Hepatocyte necrosis diffuse | 83 | 16.6 | 1 | 4.3 | 11 | 31.4 | 0.716 | 0.165 |
| Hepatocyte necrosis focal | 15 | 3.0 | 1 | 4.3 | 5 | 8.3 | 0.052 | 1.000 |
| Hepatocyte necrosis | 98 | 19.6 | 2 | 8.7 | 16 | 26.7 | 0.233 | 0.134 |
| Basophilic focus | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | - | - |
| Clear cells focus | 3 | 0.6 | 0 | 0.0 | 2 | 3.3 | 0.091 | 1.000 |
| Eosinophilic focus | 1 | 0.2 | 0 | 0.0 | 14 | 23.3 | 0.000 # | 0.008 * |
| Foci total | 4 | 0.8 | 0 | 0.0 | 15 | 25.0 | 0.000 # | 0.008 * |
| Hepatocellular Adenoma (one) | 3 | 0.6 | 0 | 0.0 | 4 | 6.7 | 0.003 # | 0.572 |
| Hepatocellular Adenoma (multiple) | 0 | 0.0 | 0 | 0.0 | 1 | 1.7 | 0.107 | 1.000 |
| Hepatocellular adenoma | 3 | 0.6 | 0 | 0.0 | 5 | 8.3 | 0.001 # | 0.316 |
| Groups | Animals | Treatment | ||
|---|---|---|---|---|
| Sex | No | Aflatoxin B1 a | S-50 Hz MF (µT) c | |
| I | M | 112 | 0 b | 0 |
| F | 103 | |||
| M + F | 215 | |||
| II | M | 103 | 70 µg/rat | 0 |
| F | 102 | |||
| M + F | 205 | |||
| III | M | 102 | 70 µg/rat | 1000 C |
| F | 120 | |||
| M + F | 222 | |||
| Total | M | 317 | ||
| F | 325 | |||
| M + F | 642 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vornoli, A.; Tibaldi, E.; Gnudi, F.; Sgargi, D.; Manservisi, F.; Belpoggi, F.; Tovoli, F.; Mandrioli, D. Evaluation of Toxicant-Associated Fatty Liver Disease and Liver Neoplastic Progress in Sprague-Dawley Rats Treated with Low Doses of Aflatoxin B1 Alone or in Combination with Extremely Low Frequency Electromagnetic Fields. Toxins 2022, 14, 325. https://doi.org/10.3390/toxins14050325
Vornoli A, Tibaldi E, Gnudi F, Sgargi D, Manservisi F, Belpoggi F, Tovoli F, Mandrioli D. Evaluation of Toxicant-Associated Fatty Liver Disease and Liver Neoplastic Progress in Sprague-Dawley Rats Treated with Low Doses of Aflatoxin B1 Alone or in Combination with Extremely Low Frequency Electromagnetic Fields. Toxins. 2022; 14(5):325. https://doi.org/10.3390/toxins14050325
Chicago/Turabian StyleVornoli, Andrea, Eva Tibaldi, Federica Gnudi, Daria Sgargi, Fabiana Manservisi, Fiorella Belpoggi, Francesco Tovoli, and Daniele Mandrioli. 2022. "Evaluation of Toxicant-Associated Fatty Liver Disease and Liver Neoplastic Progress in Sprague-Dawley Rats Treated with Low Doses of Aflatoxin B1 Alone or in Combination with Extremely Low Frequency Electromagnetic Fields" Toxins 14, no. 5: 325. https://doi.org/10.3390/toxins14050325