The Need for Next-Generation Antivenom for Snakebite Envenomation in India
by
 ,
,
Muralidharan Vanuopadath
† ,
,
Karthika Rajan
†,
Aswathy Alangode
†,
Sudarslal Sadasivan Nair
and
Bipin Gopalakrishnan Nair
* School of Biotechnology, Amrita Vishwa Vidyapeetham, Kollam 690 525, Kerala, India
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Toxins 2023, 15(8), 510; https://doi.org/10.3390/toxins15080510
Submission received: 20 May 2023
/
Revised: 5 July 2023
/
Accepted: 11 July 2023
/
Published: 18 August 2023
(This article belongs to the Special Issue Snake Venom-Omics and Next Generation Antivenom)
Abstract
:The limitations posed by currently available antivenoms have emphasized the need for alternative treatments to counteract snakebite envenomation. Even though exact epidemiological data are lacking, reports have indicated that most global snakebite deaths are reported in India. Among the many problems associated with snakebite envenomation, issues related to the availability of safer and more efficient antivenoms are of primary concern. Since India has the highest number of global snakebite deaths, efforts should be made to reduce the burden associated with snakebite envenoming. Alternative methods, including aptamers, camel antivenoms, phage display techniques for generating high-affinity antibodies and antibody fragments, small-molecule inhibitors, and natural products, are currently being investigated for their effectiveness. These alternative methods have shown promise in vitro, but their in vivo effectiveness should also be evaluated. In this review, the issues associated with Indian polyvalent antivenoms in neutralizing venom components from geographically distant species are discussed in detail. In a nutshell, this review gives an overview of the current drawbacks of using animal-derived antivenoms and several alternative strategies that are currently being widely explored.
Keywords:
snake venomics; antivenomics; immunological cross-reactivity; Indian polyvalent antivenom; next-generation antivenom; aptamers; phage display; natural products; small-molecule inhibitorsKey Contribution: This review highlights the importance of developing next-generation antivenoms for tackling snakebite envenomation. In addition, since India records the highest number of snakebite cases, this review examines the problems associated with Indian polyvalent antivenoms.
1. Introduction
Snakebite envenoming (SBE) is a significant threat to the people living in the world’s tropical regions [1]. Globally, 5.4 million snakebites are reported yearly, resulting in 1.8 to 2.7 million envenomation cases, 8100 to 138,000 deaths, and three times as many permanent disabilities and amputations [2]. A recent study indicated that approximately 64,000 deaths are reported yearly due to snakebites in India, accounting for more than 50% of the global deaths reported due to snakebites [3]. Hence, due to the degree of severity, lack of proper treatment, increasing mortality rates, and high treatment costs, in June 2017, the World Health Organization included snakebite envenoming under the neglected tropical disease category [4]. By doing so, the WHO aims to achieve a 50% reduction in morbidity and mortality rates associated with snakebite envenomation by 2030. Several strategic plans were outlined by the WHO’s expert committee on snakebite envenomation. Some of these plans are based on control and prevention, improving ecological and epidemiological data, providing better treatment for envenomed patients, and addressing antivenom issues [2]. The ‘big four’ venomous snakes that are responsible for most snakebites and associated morbidity and mortality cases in India include Bungarus caeruleus (Common krait), Daboia russelii (Russell’s viper), Echis carinatus (Indian saw-scaled viper), and Naja naja (Indian cobra) [3]. Table 1 shows India’s medically important snake species and the corresponding availability of antivenoms used to treat SBE caused by these snakes.
2. Variation in Snake Venom Composition and Antivenom Efficacy
Snake venom is a complex cocktail of several inorganic and organic constituents, including carbohydrates, lipids, proteins, and metal ions. The major class of protein families present in snake venoms are distributed among the four families are as follows: phospholipase A2 (PLA2s), snake venom metalloproteases (SVMPs), snake venom serine proteases (SVSPs), and 3- finger toxins (3-FTxs) [6]. These proteins are commonly found across snake species but in varying proportion depending on the age, sex, region, and diet of the snake species [7]. Various clinical manifestations in snakebite patients are due to the action of these toxins. Targeting these proteins and neutralizing them will serve as a better treatment option for venom-associated pathologies. The pharmacological effects induced after a snakebite may result from the synergistic effects of all these components [6,8]. Though the snake venom proteome of the most medically important snake species has been studied, a detailed investigation of the influence of various factors such as diet, body size, gender, and geographical and habitat location has not been conducted [9,10,11,12]. Investigating and compiling all of this information might be useful in delineating the venom proteome and the impact of these in influencing the efficacy of Indian polyvalent antivenoms. Due to profitability issues, most companies stopped the production of antivenoms, which has severely affected the availability of antivenoms [13]. Hence, the only option is to optimize the existing antivenoms to treat snakebite envenomation [14,15,16,17]. A conventional treatment strategy available for snakebites is the intravenous administration of antivenom [18]. Antivenom production involves injecting snake venom into an animal host, usually a horse, which generates antibodies against the venom [19,20]. Subsequently, these antibodies are isolated from the animal and used as antidotes against snakebites. Although these are effective in neutralizing systemic toxins, the effective neutralization of local effects is limited [21,22,23], leading to morbidities associated with envenomation. This could partially be due to the onset of inflammatory mediator release immediately following the snakebite [24]. Snake venom toxins have antigenic determinants shared among phylogenetically distant snake species. Assessing the cross-reactivity of snake venom to polyvalent antivenoms is needed to understand the effectiveness of existing antivenoms. These antivenoms can also be used in testing venoms that are not included in the existing immunization mixture, which could eventually shed light on the clinical application and importance of antivenom specificity [15,16,25]. Paraspecific neutralization or the cross-neutralization of antivenoms is often seen in closely related venomous snake species that share homologous venom antigenic epitopes, even if they are from different geographical locations [26,27,28,29,30]. This might be helpful in the reduced usage of venom from more snakes for generating antivenom. Nevertheless, these must be tested in suitable animal models and validated clinically before being made commercially available. In vivo and in vitro preclinical studies can be performed to determine the effectiveness of a new antivenom formulation. In addition, the manufacturers need regulatory approval to generate pan-India or region-specific antivenoms. In all of these scenarios, the antivenom manufacturers must perform sufficient preclinical studies using a reference venom mixture of medically important snake species from a specific country or geographical location [13]. This helps to evaluate the efficacy of antivenoms, which is essential because reports show that venom variation plays a crucial role in determining the clinical efficacy of antivenoms [31,32,33,34]. As seen in Table 2, there are eight antivenom manufacturing companies in India. However, it is worth noting that three are not included in the WHO Snakebite Information and Data Platform [5]. Table 3 shows numerous reports on checking the cross-neutralization and preclinical efficacy of various Indian polyvalent antivenoms using venom from Indian snakes. As indicated earlier, though three of India’s antivenom manufacturers are not listed in the WHO’s database, many in vitro preclinical studies have been performed using their antivenoms (Table 3).
3. Preclinical Studies of ‘Big Four’ Indian Snakes Using Indian Polyvalent Antivenoms
The neutralization efficacies of the antivenoms manufactured by different vendors are tested using in vivo animal models before they enter the market. However, as recommended by the WHO, in vitro preclinical studies need to be performed to evaluate the neutralization and binding capacities of these antivenoms with respect to individual snake species [13]. Interestingly, most studies use the venom from the big four snakes from various geographical locations to check various antivenom immunological responses. Out of the big four snakes, Indian cobra venom and Russell’s viper venom (Table 3) obtained from different geographical locations have been extensively studied. Venom proteome profiling of all these species indicated that the abundance and distribution of snake venom protein families vary across different geographical locations. It was also interesting to see that venom proteome variation also affects antivenom efficacy (Table 3). For example, though different groups have studied Naja naja from south India, the antivenom studies using the same venom indicated that their cross-neutralization potential varies [34,35,36,37]. This shed light on the need to generate antivenom from a geographical perspective. Nevertheless, several other factors, such as antivenom production and purification strategies, might also contribute to antivenom’s clinical effectiveness. Additionally, though eight antivenoms are commercially available (Table 2), none of the published reports have checked the preclinical evaluation of all these antivenoms in a single study. Also, it is interesting to note that most studies use antivenom from Bharat Serums, Premium serums & vaccines, and VINS Bioproducts.
4. Lethal Envenoming by ‘Non-Big Four’ Snakes in India and the Antivenom Concern
Indian polyvalent antivenoms are generated by immunizing horses using the venom pooled from the ‘big four’ snakes collected by Irula snake catchers [38]. Most mortality cases are reportedly due to bites from ‘big four’ Indian snakes, and polyvalent antivenom is provided at the appropriate doses [38,39]. However, numerous in vitro and in vivo reports have substantiated the claim that the polyvalent antivenom often fails to recognize and bind to antigenic epitopes present even in the ‘big four’ species [37,39,40,41,42]. Apart from these, there are several other medically relevant species in India, including different species of kraits, cobras, pit vipers, saw-scaled vipers, and sea snakes [43]. Clinical reports also indicate that the number of envenomation cases and the complications from envenoming induced by these ‘non-big four’ snakes are also alarmingly increasing [44,45,46]. Clinical studies have shown that venom-induced consumption coagulopathy induced by Echis carinatus sochureki venom was not neutralized after providing Indian polyvalent antivenoms [47]. However, it is worth noting that specific antivenoms are unavailable for all of these snakes (Table 1). Reports have even indicated that Indian polyvalent antivenoms are ineffective in neutralizing the venom proteins present in some of these species [48]. Due to these concerns, the WHO recommends identifying and characterizing the venom proteome of medically important snake species from a particular geographical region and generating region-specific antivenoms after performing WHO-approved preclinical tests to check the neutralization efficacy of the generated antivenoms [13]. Table 3 shows that the antivenom neutralization studies are limited to the ‘big four’ snake species and that very few studies have considered the ‘non-big four’ snake species. In this regard, in addition to exploring venom proteome profiling, it is equally important to assess an antivenom’s immunological cross-reactivity towards all of the medically important snakes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 3.
Antivenom neutralization studies using Indian polyvalent antivenom and snake venom from different geographical locations in India.
Table 3.
Antivenom neutralization studies using Indian polyvalent antivenom and snake venom from different geographical locations in India.
| SI No. | Snake Species | Geographical Location of the Snake (s) Used for the Study | Antivenom (s) Used | Antivenom Neutralization Studies Performed | Observations/Inferences | Ref. |
|---|---|---|---|---|---|---|
| 1 | Naja naja | Northwestern (Rajasthan, Gujarat) | VINS | IAC | Antivenom efficacy varies according to the geographical location of the snake species. | [34] |
| 2 | Naja naja | Eastern India—(Burdwan District, West Bengal) Calcutta snake park, Kolkata | BSAV, PSAV | ELISA, WB, IAC | Both antivenoms showed poor immunological cross-reactivity to the low-molecular-mass proteins (<20 kDa) present in Naja naja venom. | [35] |
| 3 | Naja naja | Southern India—Tamil Nadu-Irula snake catcher’s society | BSAV, PSAV, VINS, and Virchow | ELISA, WB | The low molecular mass (<15 kDa) proteins showed poor immuno-recognition compared to the high- and mid-molecular-mass proteins. | [36] |
| 4 | Naja naja | Western India—Haffkine Institute, Mumbai | BSAV, PSAV, and Virchow | ELISA, WB, IAC, neutralization of enzyme activities and anti-coagulant activity | Poor recognition of proteins with low-molecular-mass (<20 kDa) toxins present in the cobra venom from Western parts of India. | [37] |
| 5 | Naja naja | Maharashtra (West India) | BSAV, PSAV, VINS, and Haffkine | ELISA, WB, LD50, ED50 | All antivenoms recognized the venom antigenic epitopes in the ‘big four’ snake species more efficiently than other snake species. | [48] |
| 6 | Naja naja | India (exact location not mentioned) | VINS, BSAV | LD50,ED50 | BSAV antivenom was very weak in recognizing venom from other krait and cobra species. However, the VPAV effectively neutralized venom from all Southeast Asian cobras, B. candidus, N. naja, and Ophiophagus hannah with varying potencies. | [49] |
| 7 | Naja naja | Western Ghats of India-Kerala | VINS, PSAV, Virchow | ELISA, WB | Antivenomics performed using VINS antivenom indicated that it detects and binds to low-molecular-mass proteins less effectively. | [50] |
| 8 | Naja naja | North—Punjab | BSAV, PSAV, Haffkine, and VINS | ELSA, WB, LD50, ED50 | This study showed that all the antivenoms failed to neutralize Naja naja venom from desert populations. This study reiterated the need for the development of a pan-India antivenom that is effective against all snake species. | [51] |
| South—(Tamil Nadu) | ||||||
| Southeast—(Andhra Pradesh) | ||||||
| East—(West Bengal) | ||||||
| Southwest—(Maharashtra) | ||||||
| Central—(Madhya Pradesh) | ||||||
| West—(Rajasthan) | ||||||
| 9 | Naja naja | Hindustan Park (Kolkotta, West Bengal) | Haffkines | LD50, Poteolytic and hemolytic inhibitory activities | This study suggests developing region-specific antivenoms for the effective management of snakebites. | [52] |
| Irula Snake Catchers (Chennai, Tamilnadu) | ||||||
| Haffkine Institute (Mumbai, Maharashtra) | ||||||
| 10 | Naja naja, Naja oxiana, and Naja kaouthia | Himachal Pradesh, West Bengal, Mizoram, Assam, Maharashtra, Tamil Nadu (Irula) and Arunachal Pradesh | VINS, PSAV | WB, IAC | Antivenomics indicated that low-molecular-mass proteins such as PLA2 and 3FTXs were recognized poorly by the antivenom. | [53] |
| 11 | Echis carinatus | Maharashtra (West India) | BSAV, PSAV, VINS, and Haffkine | ELISA, WB, LD50, ED50 | The detection and binding efficacies of antivenoms seems to vary among all of the snake venoms tested. | [48] |
| 12 | Echis carinatus | Tamil Nadu, Goa and Rajasthan | VINS | size-exclusion chromatography | Compared venom collected from Goa and Rajasthan; the E. carniatus venom collected from Tamil Nadu resulted in the formation of more venom–antivenom complexes, indicating binding efficacy. | [54] |
| 13 | Echis carinatus | Goa and Tamil Nadu | BSAV | IAC | Low-molecular-mass proteins, especially disintegrins, present in the venom showed poor binding to the antivenom tested. | [55] |
| 14 | Echis carinatus | Southern India—Tamil Nadu, Irula snake catcher’s society | BSAV, PSAV, Virchow | ELISA, WB, IAC, and pro-coagulant activity | The antivenoms poorly recognized the low-molecular-mass proteins (<20 kDa) present in E. carinatus venom. | [42] |
| 15 | Echis carinatus sochureki | Rajasthan (Northwest India) | BSAV, PSAV, VINS, and Haffkine | ELISA, WB, LD50, ED50 | The detection and binding efficacies of the antivenoms seems vary among all of the snake venoms tested. | [48] |
| 16 | Daboia russelii | Eastern India (Nadia and Burdwan District, West Bengal)—Calcutta Snake park | BSAV, PSAV, Virchow, and BE | ELISA, WB, IAC | All of the antivenoms failed to recognize low-molecular-mass proteins (<20 kDa). | [56] |
| 17 | Daboia russelii | Southern India-Tamil Nadu-Irula snake catcher’s society | BSAV, PSAV, Virchow, and BE | ELISA, WB, IAC, and neutralization of enzyme activities and pharmacological properties | Poor recognition of the low-molecular-mass protein(<20 kDa) of Naja naja venom from Western parts of India by all the antivenoms. | [57] |
| 18 | Daboia russelii | Southern India-Tamil Nadu-Irula snake catcher’s society | Haffkine, VINS, BE, and PSAV | ELISA, WB, LD50, ED50, and IAC | The immunological cross-reactivity was different towards all of the antivenoms. | [33] |
| 19 | Daboia russelii | Western India-Haffkine Institute, Mumbai | VINS and PSAV | ELISA, WB | Both the antivenoms exhibited poor cross-reactivity towards low-molecular-mass proteins (<18 kDa) in the crude venom. The study also demonstrated that monovalent antivenoms are better than polyvalent antivenoms. | [41] |
| 20 | Daboia russelii | North- Punjab | BSAV, PSAV, Haffkine, and VINS | ELISA, WB, LD50, ED50 | The antivenoms showed poor immunological cross-reactivity against all of the venoms used, indicating the need for pan-India effective antivenoms. | [58] |
| South-(Tamil Nadu) | ||||||
| Southeast-(Andhra Pradesh) | ||||||
| East-(West Bengal) | ||||||
| Southwest-(Maharashtra) | ||||||
| Central-(Madhya Pradesh) | ||||||
| 21 | Daboia russelii | Tamil Nadu region (South India) | VINS | ELISA, LD50, procoagulant activity and neutralization | Compared to high-molecular-mass venom proteins, the low-molecular-mass proteins were poorly recognized by the antivenom. | [59] |
| 22 | Bungarus caeruleus | Southern India-Tamil Nadu-Irula snake catcher’s society | BSAV, PSAV, and BE | ELISA, WB, IAC | Poor recognition of low-molecular-mass proteins (<15 kDa) such as three-finger toxins and phospholipase A2 by the antivenoms. | [40] |
| 23 | Bungarus caeruleus | South-eastern India, unspecified locales of India, supplied by Latoxan (France) | VINS, Neuro Polyvalent Antivenom (NBAV), and Bungarus candidus Monovalent Antivenom | ELISA, LD50, ED50 | All venoms showed better immuno-reactivity profiles towards VINS antivenom. Also, compared to venom from Pakistan and Sri Lanka, Indian venom was effectively neutralized by the antivenoms. | [60] |
| 24 | Bungarus sindanus | Bikaner, Rajasthan | Haffkine and PSAV | ELISA, WB, LD50, and ED50 | The antivenom effectively neutralized B. caeruleus venom, whereas B. sindanus and B. romulusi showed poor cross-reactivity profiles towards the antivenom. | [61] |
| Bungarus sindanus | Pune, Maharashtra | |||||
| Bungarus caeruleus | Pune, Maharashtra | |||||
| Bungarus romulusi | Bannerghatta, Karnataka | |||||
| 25 | Bungarus caeruleus | Punjab (North India) | BSAV, Haffkine, PSAV, and VINS | ELISA, WB, LD50, ED50 | All the antivenoms recognized the venom antigenic epitopes in the ‘big four’ snake species and showed varied immunological cross-reactivity towards venom from other species. | [48] |
| 26 | Bungarus sindanus | Rajasthan (Northwest India) | ||||
| 27 | Bungarus fasciatus | West Bengal (East India) | ||||
| 28 | Naja kaouthia | Arunachal Pradesh (Northeast India) | ||||
| 29 | Naja kaouthia | West Bengal (East India) | ||||
| 30 | Naja kaouthia | North East India (Assam—Guwahati and Jamurighat) | VINS | WB, IAC | The VINS polyvalent antivenom could not recognize the few three-finger toxins present in Naja kaouthia venom. | [62] |
| 31 | Naja kaouthia | East India (Kolkata, West Bengal, and Arunachal Pradesh) | BSAV, Haffkine, PSAV, VINS, and Thai monovalent N. kaouthia antivenom (QSMI) | LD50, ED50, ELISA | The study concluded that intraspecies venom variation affects antivenom efficacy. | [63] |
| 32 | Naja kaouthia | Eastern India-(Burdwan District, West Bengal)-Calcutta snake park, Kolkata | BSAV, PSAV | ELISA, WB, IAC | Both antivenoms showed poor immunological cross-reactivity profiles towards the low-molecular mass proteins (<20 kDa) present in N.kaouthia venom | [35] |
| 32 | Naja kaouthia | Assam | BSAV, PSAV, Virchow, VINS | ELISA, WB | The polyvalent antivenoms poorly recognized the low-molecular-mass proteins (<15 kDa) present in N.kaouthia venom from northeastern India. | [64] |
| 34 | Naja kaouthia | North-East India and Bangladesh | VINS, Haffkine, and BSAV | WB, ED50, LD50, IAC | Antivenoms showed better immunological cross-reactivity towards high-molecular-mass components. VINS antivenom poorly recognized low-molecular-mass proteins. | [32] |
| 35 | Trimeresurus malabaricus | Western Ghats of India-Kerala | VINS, PSAV, Virchow | ELISA, WB | Compared to Russell’s viper venom, all of the antivenoms showed poor immunological cross-reactivity towards Malabar pit viper venom proteins. | [65] |
Median Lethal Dose (LD50); Median Effective Dose (ED50); Western blotting (WB); Immunoaffinity chromatography (IAC); BSAV—Bharat Serums and Vaccines Ltd.; PSAV—Premium Serum and Vaccines Pvt., Ltd.; VINS—Vins Bioproduct Limited Virchow-Virchow Biotech Pvt. Ltd.; Haffkine—Haffkine Biopharmaceuticals Corporation Ltd.; BE—Biological E Limited.
5. General Strategies for Determining the Omes and Omics of Snake Venom
The venom proteome composition of medically important snake species and the antivenom efficacy can be determined through several strategies, a few of which are discussed below:
5.1. Snake Venom Proteomics
During the late twentieth century, the identity of venom proteins was established through conventional biochemical assays and analytical strategies [66]. One of the major drawbacks of these strategies was that they failed to determine the presence of non-enzymatic proteins in snake venom. The advent of electrospray ionization [67] and matrix-assisted laser desorption ionization mass spectrometry [68,69] ionization techniques have helped to delineate the venom proteome in a different way. Based on these ionization strategies, several mass spectrometers have been extensively used to characterize the venom proteome of several snake species, including the ‘big four’ snake species in India [12,48,65,70,71,72,73,74,75,76]. The venom proteome of several snake species, including the ‘big four’ snake species in India, has been explored using various mass spectrometry-based proteomics workflows [33,35,40,42,48,77]. Venom proteome profiling can be achieved through bottom-up and top-down strategies. The latter involves intact mass measurement, the estimation of disulfide linkages, and the identification of post-translational modifications. At the same time, bottom-up proteomics involves separating venom protein components using orthogonal separation strategies, including reversed-phase chromatography and SDS-PAGE analysis. The resolved proteins are then subjected to enzymatic digestion using trypsin, chymotrypsin, and V8 protease, and the raw data can be collected using various mass spectrometers [50]. Finally, the identity of the proteins can be established through data analysis using appropriate databases. Nevertheless, several limitations exist, including the non-availability of protein sequence information in public repositories. However, this information is crucial in determining the effectiveness of commercially available antivenoms.
5.2. Genomics and Venom Gland Transcriptomics
By using prediction algorithms and homology searches (using reference sequences), translated genes (exome) can be determined through whole genome sequencing [78]. This approach can also be helpful in estimating the molecular basis of adaptation and the evolution of various snake species [79,80]. The differences in venom composition and genes’ identity in coding venom–protein sequences can easily be established through genomics rather than transcriptomics alone [81,82]. Genome sequencing can also determine structural variations such as inversions, insertions, deletions, duplications, and rearrangements [83]. Studying the snake genome not only helps delineate the identity of venom-coding genes but also helps in designing antivenoms towards the antigenic epitopes identified. For example, the genome of several snake species, including the Indian cobra, has been published [79,80,82,84,85], and this information can be utilized for designing synthetic antivenoms. Genomic information might be useful in generating humanized recombinant antivenoms [82]. This might also be useful in generating species-specific antivenoms to counteract the life-threatening envenomation effects of these snake species.
5.3. Immunological Cross-Reactivity Studies and Antivenomics
The complex toxin arsenal of various snake species could be determined through venomics studies. The data obtained through snake venomics might be useful in improving antivenom production strategies through immunological cross-reactivity studies. This will also be helpful in determining the cross-reactivity and binding potential of antivenoms toward the homologous proteins present in various snake species [86]. Since antivenoms are the main form of treatment for snakebite envenomation, efficient and reliable strategies must be constructed to assess the therapeutic potential of antivenoms [87]. Several in vitro preclinical studies include end-point titration and avidity ELISA, immunoblotting, and immunoaffinity chromatography approaches (Table 3). Immunoblotting and end-point titration ELISA are being used widely to assess the cross-reactivity potential of antivenoms. Immunoblotting is a qualitative method, whereas, through ELISA, we can effectively quantify the amount of antivenom required to detect and bind to venom antigenic epitopes. However, the venom antigens that are specifically neutralized by the antivenom need to be explored; this is a major limitation of the previously mentioned approaches. To overcome this problem, several antivenomics strategies have been outlined to effectively determine antivenom-bound and unbound venom protein constituents. This immunoaffinity-based ‘antivenomics’ approach can give more accurate quantitative information on the binding and neutralization potential of antivenoms for the treatment of snakebite envenomation [86,88] (Figure 1).
6. Alternatives to Antivenom
6.1. Aptamers
Since there are a lot of concerns associated with antivenom including the storage, production, purification, and efficacy of antivenoms, finding alternative strategies is an urgent need. Due to this, many alternatives to antivenoms, including developing and designing toxin-specific oligonucleotide-based aptamers, are currently being considered [89]. Aptamers are single-stranded DNA or RNA oligonucleotides that have high specificity and affinity towards the target [90]. They are used for a wide variety of applications [91], and one such polynucleotide aptamer, Pegaptanib, which inhibits vascular endothelial growth factor, is typically used for treating age-related macular degeneration [92]. Using Systematic Evolution of Ligands by Exponential enrichment (SELEX) technology, aptamers are selected from a library of synthesized oligonucleotides [93]. Aptamers are preferred to antibodies because of their low immunogenicity, low cost, superior shelf-life, thermal stability, smaller size, biocompatibility, and easier production strategies [94]. Reports have shown that aptamers neutralize snake venom, cone snail, and scorpion toxins [90,95,96,97,98]. Most of the studies using aptamers are limited to purified snake venom proteins. However, further studies are required to prove the efficacy of these aptamers in neutralizing entire venom protein constituents. Table 4 shows a summary of aptamers designed for the treatment of various snake venom toxins. Our detailed review on the application of aptamers as anti-dotes or as diagnostic tools indicates that the studies on the use of aptamers in the literature at the time of writing were limited to only a few species. For example, most of the aptamers were designed to target low-molecular-mass toxins, especially the neurotoxins from various krait species [90,96,99,100]. ssDNA aptamers designed to fight against snake venom serine proteases such as ancrod and batroxobin from C. rhodostoma and B. atrox inhibited venom-induced coagulopathy through reducing the consumption of fibrinogen and plasma clotting activities [93]. Thus, the use of these technologies can be expanded in order to develop aptamers for other medically important snake species.
6.2. Camel Antivenoms
Another alternative strategy to conventional antivenom production is to generate single-domain antibodies from the camel H-chain (VHH) antibody [102,103]. Contemporary antivenoms are generally based on IgG-type antibodies and Fab and F(ab)2 fragments. All of these types of antibodies have a few drawbacks, including their limitations in penetrating tissues or blood to reach their target [104]. Moreover, the presence of Fc regions in whole IgG-type antivenoms might induce severe adverse effects, including anaphylaxis and serum sickness [105,106,107,108]. Since camelid immunoglobins are devoid of light chains, they tend to interact with the antigen only through available single variable (V) regions. Hence, these antibodies isolated from camels are known as VHH or nanobodies. Therefore, these types of antibodies can be easily screened and isolated even through phage display techniques [102,103]. Moreover, the camelid IgGs are known to be less immunogenic and highly thermostable, and they feebly activate the complement cascades [109]. Several studies have shown that camelid antibodies effectively neutralize scorpion [110,111,112], Australian paralysis tick [113], and snake venom [103,104,114,115,116]. In one study, the use of camelid antivenom was found to be effective in neutralizing the hemorrhage, coagulant, lethal, and local effects of Echis carinatus sochureki venom [115]. Similarly, hemorrhagic effects induced by the venom of Echis ocellatus [104] and α-cobratoxin (from Naja kaouthia venom)-induced lethal effects [103] were found to be neutralized effectively using camelid antivenoms. All of these reports indicate that, compared to equine antivenoms, camelid antivenoms may be an effective alternative strategy for combating snake venom-induced complications.
6.3. Phage Display
Phage display [117,118] is a powerful molecular biology technique used for the screening of peptides or antibodies against target antigen. Inserting the DNA of the desired protein or peptide into the gene of the bacteriophage coat protein allows one to display these molecules on the surface of the filamentous bacteriophage. Thus, a large library of phages displaying proteins or peptides of interest can be developed and used to interact with a target protein [117]. Several groups have investigated the feasibility of using phage display to develop recombinant antibodies or peptides (Table 5) against snake venom [119], which could lead to the discovery of therapeutics which are advantageous over conventional animal-derived antivenoms. Recently, an 8-mer peptide that exhibited binding to α-cobratoxin and subsequently the inhibition of nicotinic acetylcholine receptors in Xenopus oocytes by α-cobratoxin was identified using phage display and deep sequencing [120]. These peptide-based antivenoms could act as an alternative to animal-derived antivenoms, but their efficacy and in vivo pharmacokinetics should be thoroughly evaluated. A cocktail of peptides against the major venom toxins could be effective in binding and neutralizing venom components. Here, we have reviewed the most recent advancements in developing alternative strategies through phage display to generate improved antivenoms.
Contemporary treatments for envenomation use animal-derived polyclonal whole IgG molecules, which have several disadvantages. Hence, approaches using peptides and other small molecule-based antivenoms are considered as alternative strategies. As opposed to polyclonal [126] antibodies, monoclonal antibodies or antibody fragments have been shown to serve as better options in envenomation treatments. Comparisons between polyclonal chicken IgY and horse-derived antibodies and scFv (single-chain fragment variable) are also being made. A study showed that polyclonal IgY generated to fight against Naja naja atra venom in chicken was able to recognize the low-molecular-mass proteins present in Naja naja atra venom, but horse-derived antibodies recognized only proteins with a molecular mass greater than 22 kDa. They also generated antibody libraries for monoclonal scFv (single-chain variable fragment) antibodies against Naja naja atra venom, which recognized low-molecular-weight Naja naja atra proteins. In addition, they checked their effect in vivo and found that polyclonal IgY provided full protection, whereas the neutralization of Naja naja atra venom using monoclonal scFv antibodies was only partial in mice [121]. Additional tests were performed using polyclonal IgY antibodies to fight against Trimeresurus stejnegeri venom produced by immunizing chicken with T. stejnegeri venom, and scFv antibodies were generated using phage display. The authors also compared the binding affinities of both IgY and scFv antibodies and found that scFv antibodies could recognize T. stejnegeri venom proteins, also showing some cross-reactivity towards Trimeresurus mucrosquamatus proteins. IgY antibodies provided complete protection against T. stejnegeri venom in mice, whereas a combination of scFv antibodies reduced venom-induced mortality among mice [122].
Polyclonal antibodies have limited neutralization capacities due to the low immunogenicity of some snake venom toxins. A combination of polyclonal and human monoclonal antibodies has shown to be effective in circumventing this problem. A recent report has shown that phage display can be used to generate recombinant monoclonal antibodies to fight against snake venom from two different snakes. A study was performed to generate monoclonal antibodies using phage display by cross-panning against two different cobratoxin. The selected monoclonal antibodies showed broadly neutralizing potential against three different cobras: Naja nigricollis, Naja mossambica, and Naja melanoleuca. Hence, this study demonstrates the possibility of developing broadly neutralizing monoclonal antibodies by using a single antigen, but these antibodies could be capable of neutralizing homologous toxins of the original antigen used for phage display [123]. Recently, the light chain shuffling of antibodies, where the heavy chain of the antibody is retained but alternative light chains are explored, has been proposed as another method to generate better antivenoms. Recently, a study focused on developing a cross-reactive antibody by cross-panning the light chain-shuffled scFv antibody with two different toxins—α-cobratoxin and α-elapitoxin—from the venom of Dendroaspis polylepis. The selected scFv antibodies exhibited improved affinity and increased cross-neutralization in vitro and in vivo compared to the parent antibody against the α-neurotoxins of the elapid venoms of snakes belonging to the following genera: Dendroaspis, Ophiophagus, Bungarus, and Naja [124].
Unlike the full-length antibody molecule, ScFv is a smaller version of the antibody, where the variable light chain and variable heavy chain of the Fab fragment are retained. The VH and VL are connected by a peptide linker (Figure 2). There are several advantages of selecting scFv over IgGs, including the benefits related to size. The small size of scFv allows for the generation of these molecules in bacterial expression systems, while antibodies require mammalian systems. Additionally, they are small enough to be displayed on the phage; thus, they are easier to screen using the phage display method (Figure 3). Moreover, since phage display is an in vitro technique, animal immunization can be avoided. Faster clearance from the blood, better diffusion into tissues, and less immunogenicity due to the lack of Fc region are some of the most important advantages of using scFv. The Fc region can activate Fc receptor-expressing cells, thus resulting in a massive release of cytokines and leading to toxicity [127].
A recent study has shown that a monoclonal scFv antibody generated to fight against the cytotoxins of Naja atra venom was able to neutralize and prevent cytotoxicity in C2C12 myoblast cells [126]. Similarly, Cro-Bothrumabs is a human scFv fragment that recognizes venoms of both bothrops and crotalus. This is the first report involving a polyvalent antivenom fighting against Bothrops jararacassu and Crotalus durissus terificus. The antibody fragment was shown to be capable of neutralizing both of the venoms in both in vitro and in vivo experiments [125].
Thus, phage display is a promising technique that can be used to develop better antivenoms due to its specificity and lack of adverse effects compared to the techniques used to develop animal-derived antivenoms. However, to date, there is no published data pertaining to India available on the use of phage display for developing better antivenoms.
6.4. Small-Molecule Inhibitors
Due to the above-mentioned limitations of antivenoms, including the inability to neutralize low-molecular-weight toxins of venom, which can have deleterious effects, the repurposing of small-molecule inhibitors against these toxins has become a major area of research for the development of better treatment strategies for snake envenomation. There are various studies showing the inhibitory action of these small-molecule inhibitors on various toxins like phospholipase A2 [128] and metalloproteases in venom (Table 6). The most studied among them is varespladib (LY315920), which is a non-specific inhibitor of mammalian sPLA2 that has previously been used to treat acute coronary syndrome [129] but failed in a Phase II clinical trial. In 2016, Lewin et al. demonstrated that varespladib could inhibit PLA2 from the snake venoms of several snake species from six different continents [130]. Three other compounds—prinomastat, batimastat, and marimastat (Figure 4)—had a similar effect on the action of venom metalloproteases. These molecules were initially used to treat tumors but were shown to inhibit the metalloproteases of snake venom [131].
A recent study has shown that varespladib was able to inhibit the anticoagulant and procoagulant activities of venom. A study conducted by Xie et al. found that varespladib might also interfere with other toxins since, in their study, they observed that varespladib could partially abrogate the procoagulant activities of venom. Procoagulant activities are caused by the action of proteases [132]. Further studies are required to assess whether varespladib could affect other venom toxins also [133]. This could be an important finding since both PLA2 and snake venom metalloproteases are responsible for necrosis leading to tissue damage after envenomation, and these effects may not be neutralized effectively due to delays in the administration of antivenoms. In these scenarios, alternate inhibitors could be effective as a first-aid treatment prior to antivenom administration, as shown in previous studies [134,135]. It is still unknown whether varespladib can abrogate the effects of procoagulant toxins present in other snake venoms since the aforementioned study was performed only with the venom of a few snake species.
Table 6.
Studies on small-molecule inhibitors against snake venom.
| SI No. | Small-Molecule Inhibitor | Snake Species | Protein | Study Design | Ref. |
|---|---|---|---|---|---|
| 1 | Varespladib | Bothrops asper, Calloselasma rhodostoma, Deinagkistrodon acutus, Daboia russelii, Echis carinatus, Echis ocellatus, and Oxyuranus scutellatus. | Phospholipase A2 | In vitro | [133] |
| 2 | Varespladib | Daboia siamensis | Phospholipase A2 | In vitro | [136] |
| 3 | Batimastat | Crotalus atrox | Group I (PI) metalloprotease | In vitro and in silico | [137] |
| 4 | Marimastat | Crotalus atrox | Group I (PI) metalloprotease | In vitro and in silico | [137] |
| 5 | Varespladib | Naja ashei, Naja katiensis, and Naja nubiae | Phospholipase A2 | In vitro | [138] |
| 6 | Prinomastat | Naja ashei, Naja katiensis, and Naja nubiae | Phospholipase A2 | In vitro | [138] |
Similarly, another study assessed the effectiveness of antivenom alone and in conjunction with varespladib on the neurotoxicity induced by Chinese D. siamensis venom. The study was performed in vitro by isolating the chick biventer cervicis nerve muscle preparation in an organ bath to assess the effect of antivenom alone and in combination with varespladib on the neuromuscular blockade induced by the venom. The researchers found that neurotoxicity cannot be reversed by the monovalent antivenom. But the pre-incubation of venom with varespladib abrogated the neurotoxic and myotoxic activities of the venom.
However, when both venom and varespladib were given in combination, it did not result in an increased inhibitory effect on neurotoxicity or myotoxicity [136]. Additionally, other studies showing the inhibitory effect of varespladib on neurotoxic PLA2s have also been performed. In 2020, Gutierezz et al. showed that varespladib and methyl varespladib were able to rescue mice injected with venoms of Notechis scutatus, Crotalus durissus terrificus, Bungarus multicinctus, and Oxyuranus scutellatus, where pre-synaptically acting neurotoxic PLA2s with different quarternary structures were major toxins in these venoms [139]. Hence, this study also aimed to check whether varespladib was able to inhibit the PLA2s with a different quaternary structure. A recent study by Tan et al. in 2022 demonstrated the effect of varespladib on the neurotoxic activities of the five major kraits of Asia: B. caeruleus, B. sindanus, B. candidus, B. fasciatus, and B. multicinctus. These reports indicate the efficacy of varespladib in combating the toxic effect of venom PLA2 and thus could be used as an adjunct treatment for snake envenomation [140]. Thus, several in vitro and in vivo studies have been conducted to evaluate the preclinical efficacy of varespladib. Following this, a study protocol, BRAVO (Broad-spectrum Rapid Antidote: Varespladib Oral for snakebite), was recently designed to evaluate the clinical effects of varespladib-methyl. This is the first international clinical trial of a treatment for snakebite envenomation [141]. This study was designed to compare and evaluate the efficacy of varespladib-methyl plus standard of care with placebo plus standard of care in patients affected by snakebite envenomation.
Like varespladib, there are matrix metalloprotease inhibitors that could also prove to be promising against snake envenomation since they can inhibit snake venom metalloproteases. These molecules were initially developed for treating cancer, but they failed in clinical trials [142,143,144]. Recently, they were shown to inhibit the SVMP activity of western diamondback rattlesnake venom [137]. From all of these studies, it is likely that small-molecule inhibitors could be used as a form of complementary or adjuvant therapy in conjunction with antivenom administration. Furthermore, these molecules could be used as a first-aid treatment in a field setting where antivenom administration could be time-consuming due to a lack of availability regarding healthcare facilities. This could prevent delays in treating snakebite victims and thus avoid significant morbidity and mortality rates due to envenomation. Small-molecule inhibitors are usually targeted against a particular venom toxin, but since the venom components act in a synergistic way, a combination of these inhibitors might be appropriate to use as an adjuvant or first-aid treatment before antivenom administration.
From an Indian perspective, studying the effect of small-molecule inhibitors could prove promising. About half of the snakebite deaths occurring in India are due to envenomation by Russell’s viper, the venom of which has many toxins that affect the coagulation mechanism of the victim. PLA2 is the most abundant toxin of RV venom [57], and there are many isoforms of this toxin [31] in the venom of a single snake species. With a low molecular weight, small size, and, consequently, being less immunogenic, these toxins are not always effectively neutralized by the polyvalent antivenoms developed in India. Hence, alternate strategies like exploring the effect of the small-molecule inhibitors of these toxins should be developed to fight against limitations of antivenom.
6.5. Natural Products
Time is an essential factor governing the pathophysiology of envenomation, and most of the snakebite cases are reported from rural areas, where antivenom facilities are scarce and hospitals are usually out of reach [23]. In these scenario, using a traditional plant-based treatment as an alternative or supporting therapy to circumvent an antivenom’s limitations is of great importance.
Traditional medicines make use of plant extracts or derived compounds like secondary metabolites to treat snakebites. These products can be procured with ease, are relatively economical, and bring fewer administrational complications [24]. The therapeutic applications possessed by plants can be of two types: On one hand, the active components present in crude plant extracts can be used as inhibitors for snake venom toxins. On the other hand, the chemical structure of active constituents can be modified to target certain toxins in venom [145]. The pathways targeted by the venom toxins can be used as a target to develop inhibitors from plant products, and such an effort was shown in a study in which aqueous stem bark extracts of Mangifera indica were used to inhibit Group IA sPLA2 (phospholipase A2), which are enzymes that regulate the release of inflammatory mediators via the arachidonic acid pathway, thereby playing an important role in inflammation at the bite site [146]. The active constituents present in the plant extracts exert their action by interfering with the binding sites of the substrate or enzyme or by chelating the metal ions needed for enzymatic activity, inhibiting the enzyme [7]. The separation of the active components behind neutralization is one of the major limitations in developing the natural product-based inhibitors of snake venom. Several studies have been successful in isolating bioactive compounds from plant extracts. However, the validation of their toxicity profiles is a major hindrance to its development as a drug candidate [147].
Medicinally important plants can be screened to find inhibitors that act against different components present in snake venom and isolate the lead molecule responsible for the inhibitory effects [148]. Previous studies have demonstrated that various parts of plants can be used to treat snakebites and their associated symptoms. Leaves are commonly used to treat snakebites. Among the botanical family of plants, the most studied family regarding potential snake venom-inhibiting substances is the Fabaceae, followed by the Zingiberaceae, Salicaceae, and Asteraceae families [24]. Regarding secondary metabolites derived from plants with snake venom-inhibiting properties, studies have shown that phenolic compounds are the major class of compounds that contribute to snake venom neutralization [24]. Natural products can be designed to inhibit various toxin families present in snake venom (Table 7), as these are the class of proteins that act upon different physiological phenomena in the victim, leading to clinical manifestations such as local tissue damage [149,150].
A lot of plant species have reportedly shown anti-PLA2 activity. One of the main drawbacks of contemporary antivenoms is that the low-molecular-mass toxins in the venom are poorly recognized compared to high-molecular-mass components (Table 3). As these proteins are less immunogenic, antibodies designed to fight against these proteins are not efficiently generated, leading to low neutralization. One such low-molecular-mass protein is PLA2, which is responsible for inflammation and pain in bitten areas [151]. Plants used to treat inflammatory disorders could be screened for inhibitors of the enzyme phospholipase A2. One such inhibitor is Aristolochic acid, which is extracted from the plant root of the Aristolochia species and has been reported to inhibit PLA2 and L-amino acid oxidase (LAAO) components in snake venom [152,153]; however, due to the DNA-binding property of this molecule, it is considered to be carcinogenic. Hence, a modified semi-synthetic derivative of the molecule is used for inhibition [152]. One study reported the neutralization of the PLA2 enzyme from Cobra and Russell’s viper venom using a compound isolated from neem leaf extract, AIPLAI (Azadirachta indica PLA2 inhibitor) [154]. A later study demonstrated the use of neem plants for treating poisonous bites, including snakebites, by the Irula tribes of Walayar valley, South India, who practice herbal medicines using the indigenous plants found in the region to treat different ailments [155]. This information substantiates the previous research findings on AIPLAI. Further studies on the aqueous root extract of Mimosa pudica showed inhibitory activity towards the hyaluronidase and protease enzymes of Naja naja, Echis carinatus, and Daboia russelli venom [156]. The aqueous extract of Morus alba leaves has been shown to inhibit the systemic effects and local tissue damage induced by Russell’s viper venom. The extract acts against venom metalloproteinases (SVMP), wherein the metal ion chelation by the extract leads to the inactivation of the enzyme [157]. However, the active components responsible for these activities were not isolated. Many studies have been performed with compounds isolated from plants like secondary metabolites; one particular study involving a compound, quercetin-3-O-α-rhamnoside, from the Indian species Euphorbia hirta (Euphorbiaceae) significantly inhibited the PLA2 enzyme from N. naja by 93%. It also inhibited hyaluronidase and hemolytic activity [158]. Regarding plants endemic to India, a H. indicus (Apocynaceae)-derived compound, 2-hydroxy-4-methoxy benzoic acid, has shown inhibitory effects on N. kaouthia, O. hannah, D. russelii, and E. carinatus venom. The compound effectively inhibited D. russelii venom-induced inflammation and hemorrhage and coagulant effects in mouse models [159,160,161]. Apart from Flavonoids, Alkaloids, and secondary metabolites, a glycoprotein isolated from a Withania somnifera plant, WSG, has shown inhibitory potential towards cobra venom toxic PLA2 [162]. This is an example of mimicking the structure analogy of natural PLA2 inhibitors present in snake plasma. The glycoprotein WSG is similar to the α-chain of the phospholipase inhibitors(PLIs) found in snake plasma [163,164].
Considering the vast opportunities plant products offer, in silico studies using molecular docking could be employed to identify the bioactive components that help bind to the active site of the toxins in snake venom and study their interaction mechanisms. This could help in designing modified plant compounds with better binding ability and hence improved neutralization potential [165,166].
7. Conclusions
Since geographical venom variation contributes to antivenom efficacy, from a regional perspective, there is a pressing need to develop antivenom. Antivenom availability is limited in certain countries, including Sri Lanka, Bangladesh, Pakistan, and Nepal. Hence, these countries depend on Indian polyvalent antivenoms generated using the venom collected from the big four snakes of India. When numerous publications report the ineffectiveness of Indian antivenoms even towards Indian snakes, it is alarming not only for Indian citizens but also for those in neighboring countries that depend on Indian antivenoms to treat snakebites. Preclinical studies have indicated the inefficiency of Indian polyvalent antivenoms in detecting and binding to the venom epitopes present in the snake species of other countries, and few studies agree with the cross-neutralization of homologous venom proteins. However, their efficacy towards proteins of a heterologous nature is still a concern, and more studies need to be performed to validate the results of prior studies. Additionally, several reports have indicated that, in addition to venom from the ‘big four’ snakes, venom from other medically important snake species needs to be included in the horse immunization mixture when developing antivenoms. Since the morbidity and mortality rates are high, effective measures need to be outlined to reduce the burden imposed by this neglected disease. To achieve this, several alternatives have been proposed, including synthetic antibodies, repurposed drugs, and small-molecule inhibitors; however, all of these potential treatments need to be approved in clinical trials. Some of the major drawbacks of antivenoms include the fact they do not completely neutralize the local effects of envenomation, they have low neutralization capabilities with respect to low-molecular-weight proteins, and the fact that serum sickness towards the host antigen present in the antivenom can occur. These strategies can also be implemented along with antivenom treatment so that it will slow down some of the effects induced after SBE. Nevertheless, clinical trials should be performed to test the efficacy of administering these treatments in conjunction with antivenom. To conclude, since comprehensive expertise is not available under a single roof, thorough inter-institutional and intra-institutional collaboration within the country and outside India is needed to explore the potential of implementing these alternatives to counteract snakebite envenomation.
Author Contributions
Conceptualization, M.V. and B.G.N.; writing—original draft preparation, M.V., K.R. and A.A.; writing—review and editing, M.V., K.R., A.A., S.S.N. and B.G.N.; supervision and funding acquisition, B.G.N. All authors have read and agreed to the published version of the manuscript.
Funding
Muralidharan V. is supported by the Young Scientist Fellowship program provided by the Department of Health Research, Indian Council of Medical Research, Government of India, (grant number—File No.R.12014/31/2022-HR) and the APC was funded by Amrita Vishwa Vidyapeetham.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Mata Amritanadamayi Devi, for her constant support. The authors acknowledge the kind help they received from Vishnu Raj in collecting and compiling the tabulated information in Table 3.
Conflicts of Interest
The authors declare no conflict of interest.
References
- The Lancet. Snake-bite envenoming: A priority neglected tropical disease. Lancet 2017, 390, 2. [Google Scholar] [CrossRef] [PubMed]
- Snakebite Envenoming. Available online: https://www.who.int/news-room/fact-sheets/detail/snakebite-envenoming (accessed on 18 May 2023).
- Roberts, N.L.S.; Johnson, E.K.; Zeng, S.M.; Hamilton, E.B.; Abdoli, A.; Alahdab, F.; Alipour, V.; Ancuceanu, R.; Andrei, C.L.; Anvari, D.; et al. Global mortality of snakebite envenoming between 1990 and 2019. Nat. Commun. 2022, 13, 6160. [Google Scholar]
- Chippaux, J.P. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef]
- World Health Organisation-Snakebite Information and Data Platform. Available online: https://snbdatainfo.who.int/ (accessed on 18 May 2023).
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, M.S.; Hemshekhar, M.; Sunitha, K.; Thushara, R.M.; Jnaneshwari, S.; Kemparaju, K.; Girish, K.S. Snake venom induced local toxicities: Plant secondary metabolites as an auxiliary therapy. Mini Rev. Med. Chem. 2013, 13, 106–123. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; Viegas, M.F.; da Silva, S.L.; Soares, A.M.; Ramos, M.J.; Fernandes, P.A. The chemistry of snake venom and its medicinal potential. Nat. Rev. Chem. 2022, 6, 451–469. [Google Scholar] [CrossRef]
- Menezes, M.C.; Furtado, M.F.; Travaglia-Cardoso, S.R.; Camargo, A.C.; Serrano, S.M. Sex-based individual variation of snake venom proteome among eighteen Bothrops jararaca siblings. Toxicon 2006, 47, 304–312. [Google Scholar] [CrossRef]
- Daltry, J.C.; Wuster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef]
- Gibbs, H.L.; Mackessy, S.P. Functional basis of a molecular adaptation: Prey-specific toxic effects of venom from Sistrurus rattlesnakes. Toxicon 2009, 53, 672–679. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Leroy, J.; Mocino-Deloya, E.; Setser, K.; Bryson, R.W.; Saviola, A.J. Venom Ontogeny in the Mexican Lance-Headed Rattlesnake (Crotalus polystictus). Toxins 2018, 10, 271. [Google Scholar] [CrossRef]
- Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins. Available online: https://apps.who.int/iris/bitstream/handle/10665/255657/9789241210133-eng.pdf#page=217 (accessed on 18 May 2023).
- Brown, N.I. Consequences of neglect: Analysis of the sub-Saharan African snake antivenom market and the global context. PLoS Negl. Trop. Dis. 2012, 6, e1670. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Gutierrez, J.M.; Calvete, J.J.; Wuster, W.; Ratanabanangkoon, K.; Paiva, O.; Brown, N.I.; Casewell, N.R.; Harrison, R.A.; Rowley, P.D.; et al. Ending the drought: New strategies for improving the flow of affordable, effective antivenoms in Asia and Africa. J. Proteom. 2011, 74, 1735–1767. [Google Scholar] [CrossRef]
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Negl. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef]
- Habib, A.G.; Brown, N.I. The snakebite problem and antivenom crisis from a health-economic perspective. Toxicon 2018, 150, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Leon, G.; Lomonte, B.; Angulo, Y. Antivenoms for snakebite envenomings. Inflamm. Allergy Drug. Targets 2011, 10, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Bermudez-Mendez, E.; Fuglsang-Madsen, A.; Fons, S.; Lomonte, B.; Gutierrez, J.M.; Laustsen, A.H. Innovative Immunization Strategies for Antivenom Development. Toxins 2018, 10, 452. [Google Scholar] [CrossRef]
- Otero, R.; Fonnegra, R.; Jimenez, S.L.; Nunez, V.; Evans, N.; Alzate, S.P.; Garcia, M.E.; Saldarriaga, M.; Del Valle, G.; Osorio, R.G.; et al. Snakebites and ethnobotany in the northwest region of Colombia: Part I: Traditional use of plants. J. Ethnopharmacol. 2000, 71, 493–504. [Google Scholar] [CrossRef]
- Gimenes, S.N.C.; Sachett, J.A.G.; Colombini, M.; Freitas-de-Sousa, L.A.; Ibiapina, H.N.S.; Costa, A.G.; Santana, M.F.; Park, J.J.; Sherman, N.E.; Ferreira, L.C.L.; et al. Observation of Bothrops atrox Snake Envenoming Blister Formation from Five Patients: Pathophysiological Insights. Toxins 2021, 13, 800. [Google Scholar] [CrossRef]
- Puzari, U.; Fernandes, P.A.; Mukherjee, A.K. Pharmacological re-assessment of traditional medicinal plants-derived inhibitors as antidotes against snakebite envenoming: A critical review. J. Ethnopharmacol. 2022, 292, 115208. [Google Scholar] [CrossRef]
- Adriao, A.A.X.; Dos Santos, A.O.; de Lima, E.; Maciel, J.B.; Paz, W.H.P.; da Silva, F.M.A.; Pucca, M.B.; Moura-da-Silva, A.M.; Monteiro, W.M.; Sartim, M.A.; et al. Plant-Derived Toxin Inhibitors as Potential Candidates to Complement Antivenom Treatment in Snakebite Envenomations. Front. Immunol. 2022, 13, 842576. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef]
- Ainsworth, S.; Slagboom, J.; Alomran, N.; Pla, D.; Alhamdi, Y.; King, S.I.; Bolton, F.M.S.; Gutierrez, J.M.; Vonk, F.J.; Toh, C.H.; et al. The paraspecific neutralisation of snake venom induced coagulopathy by antivenoms. Commun. Biol. 2018, 1, 34. [Google Scholar] [CrossRef] [PubMed]
- Sandesha, V.D.; Darshan, B.; Tejas, C.; Girish, K.S.; Kempaiah, K. A comparative cross-reactivity and paraspecific neutralization study on Hypnale hypnale, Echis carinatus, and Daboia russelii monovalent and therapeutic polyvalent anti-venoms. PLoS Negl. Trop. Dis. 2022, 16, e0010292. [Google Scholar] [CrossRef] [PubMed]
- Oh, A.M.F.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Venom proteome of Bungarus sindanus (Sind krait) from Pakistan and in vivo cross-neutralization of toxicity using an Indian polyvalent antivenom. J. Proteom. 2019, 193, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Faisal, T.; Tan, K.Y.; Sim, S.M.; Quraishi, N.; Tan, N.H.; Tan, C.H. Proteomics, functional characterization and antivenom neutralization of the venom of Pakistani Russell’s viper (Daboia russelii) from the wild. J. Proteom. 2018, 183, 1–13. [Google Scholar] [CrossRef]
- Maduwage, K.; Silva, A.; O’Leary, M.A.; Hodgson, W.C.; Isbister, G.K. Efficacy of Indian polyvalent snake antivenoms against Sri Lankan snake venoms: Lethality studies or clinically focussed in vitro studies. Sci. Rep. 2016, 6, 26778. [Google Scholar] [CrossRef]
- Kalita, B.; Mackessy, S.P.; Mukherjee, A.K. Proteomic analysis reveals geographic variation in venom composition of Russell’s Viper in the Indian subcontinent: Implications for clinical manifestations post-envenomation and antivenom treatment. Expert. Rev. Proteom. 2018, 15, 837–849. [Google Scholar] [CrossRef]
- Deka, A.; Reza, M.A.; Faisal Hoque, K.M.; Deka, K.; Saha, S.; Doley, R. Comparative analysis of Naja kaouthia venom from North-East India and Bangladesh and its cross reactivity with Indian polyvalent antivenoms. Toxicon 2019, 164, 31–43. [Google Scholar] [CrossRef]
- Pla, D.; Sanz, L.; Quesada-Bernat, S.; Villalta, M.; Baal, J.; Chowdhury, M.A.W.; Leon, G.; Gutierrez, J.M.; Kuch, U.; Calvete, J.J. Phylovenomics of Daboia russelii across the Indian subcontinent. Bioactivities and comparative in vivo neutralization and in vitro third-generation antivenomics of antivenoms against venoms from India, Bangladesh and Sri Lanka. J. Proteom. 2019, 207, 103443. [Google Scholar] [CrossRef]
- Sintiprungrat, K.; Watcharatanyatip, K.; Senevirathne, W.D.; Chaisuriya, P.; Chokchaichamnankit, D.; Srisomsap, C.; Ratanabanangkoon, K. A comparative study of venomics of Naja naja from India and Sri Lanka, clinical manifestations and antivenomics of an Indian polyspecific antivenom. J. Proteom. 2016, 132, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Patra, A.; Kalita, B.; Mukherjee, A.K. Proteomics analysis to compare the venom composition between Naja naja and Naja kaouthia from the same geographical location of eastern India: Correlation with pathophysiology of envenomation and immunological cross-reactivity towards commercial polyantivenom. Expert Rev. Proteom. 2018, 15, 949–961. [Google Scholar]
- Chanda, A.; Mukherjee, A.K. Quantitative proteomics to reveal the composition of Southern India spectacled cobra (Naja naja) venom and its immunological cross-reactivity towards commercial antivenom. Int. J. Biol. Macromol. 2020, 160, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Kalita, B.; Patra, A.; Senevirathne, W.; Mukherjee, A.K. Proteomic analysis and antivenomics study of Western India Naja naja venom: Correlation between venom composition and clinical manifestations of cobra bite in this region. Expert Rev. Proteom. 2019, 16, 171–184. [Google Scholar] [CrossRef]
- Warrell, D.A.; Gutierrez, J.M.; Calvete, J.J.; Williams, D. New approaches & technologies of venomics to meet the challenge of human envenoming by snakebites in India. Indian J. Med. Res. 2013, 138, 38–59. [Google Scholar]
- Simpson, I.D.; Norris, R.L. Snakes of medical importance in India: Is the concept of the “Big 4” still relevant and useful? Wilderness Environ. Med. 2007, 18, 2–9. [Google Scholar] [CrossRef]
- Patra, A.; Chanda, A.; Mukherjee, A.K. Quantitative proteomic analysis of venom from Southern India common krait (Bungarus caeruleus) and identification of poorly immunogenic toxins by immune-profiling against commercial antivenom. Expert Rev. Proteom. 2019, 16, 457–469. [Google Scholar] [CrossRef]
- Kalita, B.; Patra, A.; Mukherjee, A.K. Unraveling the Proteome Composition and Immuno-profiling of Western India Russell’s Viper Venom for In-Depth Understanding of Its Pharmacological Properties, Clinical Manifestations, and Effective Antivenom Treatment. J. Proteome Res. 2017, 16, 583–598. [Google Scholar] [CrossRef]
- Patra, A.; Kalita, B.; Chanda, A.; Mukherjee, A.K. Proteomics and antivenomics of Echis carinatus carinatus venom: Correlation with pharmacological properties and pathophysiology of envenomation. Sci. Rep. 2017, 7, 17119. [Google Scholar] [CrossRef]
- Whitaker, R.; Martin, G. Diversity and Distribution of Medically Important Snakes of India. In Clinical Toxinology in Asia Pacific and Africa; Gopalakrishnakone, P., Faiz, A., Fernando, R., Gnanathasan, C., Habib, A., Yang, C.C., Eds.; Springer: Dordrecht, the Netherlands, 2015. [Google Scholar]
- Joseph, J.K.; Simpson, I.D.; Menon, N.C.; Jose, M.P.; Kulkarni, K.J.; Raghavendra, G.B.; Warrell, D.A. First authenticated cases of life-threatening envenoming by the hump-nosed pit viper (Hypnale hypnale) in India. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 85–90. [Google Scholar] [CrossRef]
- Kumar, K.S.; Narayanan, S.; Udayabhaskaran, V.; Thulaseedharan, N.K. Clinical and epidemiologic profile and predictors of outcome of poisonous snake bites—An analysis of 1500 cases from a tertiary care center in Malabar, North Kerala, India. Int. J. Gen. Med. 2018, 11, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Herath, N.; Wazil, A.; Kularatne, S.; Ratnatunga, N.; Weerakoon, K.; Badurdeen, S.; Rajakrishna, P.; Nanayakkara, N.; Dharmagunawardane, D. Thrombotic microangiopathy and acute kidney injury in hump-nosed viper (Hypnale species) envenoming: A descriptive study in Sri Lanka. Toxicon 2012, 60, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, M.; Yadav, P.; Mathur, R.; Midha, N.; Garg, M.K. Venom-Induced Consumption Coagulopathy Unresponsive to Antivenom After Echis carinatus sochureki Envenoming. Wilderness Environ. Med. 2021, 32, 221–225. [Google Scholar] [CrossRef]
- Senji Laxme, R.R.; Khochare, S.; de Souza, H.F.; Ahuja, B.; Suranse, V.; Martin, G.; Whitaker, R.; Sunagar, K. Beyond the ‘big four’: Venom profiling of the medically important yet neglected Indian snakes reveals disturbing antivenom deficiencies. PLoS Negl. Trop. Dis. 2019, 13, e0007899. [Google Scholar] [CrossRef] [PubMed]
- Leong, P.K.; Tan, N.H.; Fung, S.Y.; Sim, S.M. Cross neutralisation of Southeast Asian cobra and krait venoms by Indian polyvalent antivenoms. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Vanuopadath, M.; Raveendran, D.; Nair, B.G.; Nair, S.S. Venomics and antivenomics of Indian spectacled cobra (Naja naja) from the Western Ghats. Acta Trop. 2022, 228, 106324. [Google Scholar] [CrossRef]
- Senji Laxme, R.R.; Attarde, S.; Khochare, S.; Suranse, V.; Martin, G.; Casewell, N.R.; Whitaker, R.; Sunagar, K. Biogeographical venom variation in the Indian spectacled cobra (Naja naja) underscores the pressing need for pan-India efficacious snakebite therapy. PLoS Negl. Trop. Dis. 2021, 15, e0009150. [Google Scholar] [CrossRef]
- Shashidharamurthy, R.; Kemparaju, K. Region-specific neutralization of Indian cobra (Naja naja) venom by polyclonal antibody raised against the eastern regional venom: A comparative study of the venoms from three different geographical distributions. Int. Immunopharmacol. 2007, 7, 61–69. [Google Scholar] [CrossRef]
- Deka, A.; Bhatia, S.; Santra, V.; Bharti, O.K.; Lalremsanga, H.T.; Martin, G.; Wuster, W.; Owens, J.B.; Graham, S.; Doley, R.; et al. Multilevel Comparison of Indian Naja Venoms and Their Cross-Reactivity with Indian Polyvalent Antivenoms. Toxins 2023, 15, 258. [Google Scholar] [CrossRef]
- Bhatia, S.; Blotra, A.; Vasudevan, K. Evaluating Antivenom Efficacy against Echis carinatus Venoms-Screening for In Vitro Alternatives. Toxins 2022, 14, 481. [Google Scholar] [CrossRef]
- Bhatia, S.; Blotra, A.; Vasudevan, K. Immunorecognition capacity of Indian polyvalent antivenom against venom toxins from two populations of Echis carinatus. Toxicon 2021, 201, 148–154. [Google Scholar] [CrossRef]
- Kalita, B.; Patra, A.; Das, A.; Mukherjee, A.K. Proteomic Analysis and Immuno-Profiling of Eastern India Russell’s Viper (Daboia russelii) Venom: Correlation between RVV Composition and Clinical Manifestations Post RV Bite. J. Proteome Res. 2018, 17, 2819–2833. [Google Scholar] [CrossRef] [PubMed]
- Kalita, B.; Singh, S.; Patra, A.; Mukherjee, A.K. Quantitative proteomic analysis and antivenom study revealing that neurotoxic phospholipase A2 enzymes, the major toxin class of Russell’s viper venom from southern India, shows the least immuno-recognition and neutralization by commercial polyvalent antivenom. Int. J. Biol. Macromol. 2018, 118, 375–385. [Google Scholar] [PubMed]
- Senji Laxme, R.R.; Khochare, S.; Attarde, S.; Suranse, V.; Iyer, A.; Casewell, N.R.; Whitaker, R.; Martin, G.; Sunagar, K. Biogeographic venom variation in Russell’s viper (Daboia russelii) and the preclinical inefficacy of antivenom therapy in snakebite hotspots. PLoS Negl. Trop. Dis. 2021, 15, e0009247. [Google Scholar] [CrossRef]
- Faisal, T.; Tan, K.Y.; Tan, N.H.; Sim, S.M.; Gnanathasan, C.A.; Tan, C.H. Proteomics, toxicity and antivenom neutralization of Sri Lankan and Indian Russell’s viper (Daboia russelii) venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2021, 27, e20200177. [Google Scholar] [CrossRef]
- Oh, A.M.F.; Tan, C.H.; Ariaranee, G.C.; Quraishi, N.; Tan, N.H. Venomics of Bungarus caeruleus (Indian krait): Comparable venom profiles, variable immunoreactivities among specimens from Sri Lanka, India and Pakistan. J. Proteom. 2017, 164, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sunagar, K.; Khochare, S.; Senji Laxme, R.R.; Attarde, S.; Dam, P.; Suranse, V.; Khaire, A.; Martin, G.; Captain, A. A Wolf in Another Wolf’s Clothing: Post-Genomic Regulation Dictates Venom Profiles of Medically-Important Cryptic Kraits in India. Toxins 2021, 13, 69. [Google Scholar] [CrossRef]
- Deka, A.; Gogoi, A.; Das, D.; Purkayastha, J.; Doley, R. Proteomics of Naja kaouthia venom from North East India and assessment of Indian polyvalent antivenom by third generation antivenomics. J. Proteom. 2019, 207, 103463. [Google Scholar] [CrossRef]
- Rashmi, U.; Khochare, S.; Attarde, S.; Laxme, R.R.S.; Suranse, V.; Martin, G.; Sunagar, K. Remarkable intrapopulation venom variability in the monocellate cobra (Naja kaouthia) unveils neglected aspects of India’s snakebite problem. J. Proteom. 2021, 242, 104256. [Google Scholar] [CrossRef]
- Kakati, H.; Patra, A.; Kalita, B.; Chanda, A.; Rapole, S.; Mukherjee, A.K. A comparison of two different analytical workflows to determine the venom proteome composition of Naja kaouthia from North-East India and immunological profiling of venom against commercial antivenoms. Int. J. Biol. Macromol. 2022, 208, 275–287. [Google Scholar] [CrossRef]
- Vanuopadath, M.; Shaji, S.K.; Raveendran, D.; Nair, B.G.; Nair, S.S. Delineating the venom toxin arsenal of Malabar pit viper (Trimeresurus malabaricus) from the Western Ghats of India and evaluating its immunological cross-reactivity and in vitro cytotoxicity. Int. J. Biol. Macromol. 2020, 148, 1029–1045. [Google Scholar] [CrossRef] [PubMed]
- Chanda, A.; Mukherjee, A.K. Mass spectrometric analysis to unravel the venom proteome composition of Indian snakes: Opening new avenues in clinical research. Expert Rev. Proteom. 2020, 17, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Fenn, J.B.; Mann, M.; Meng, C.K.; Wong, S.F.; Whitehouse, C.M. Electrospray ionization for mass spectrometry of large biomolecules. Science 1989, 246, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Karas, M.; Hillenkamp, F. Laser desorption ionization of proteins with molecular masses exceeding 10,000 daltons. Anal. Chem. 1988, 60, 2299–2301. [Google Scholar] [CrossRef]
- Zaluzec, E.J.; Gage, D.A.; Watson, J.T. Matrix-assisted laser desorption ionization mass spectrometry: Applications in peptide and protein characterization. Protein Expr. Purif. 1995, 6, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Rey-Suarez, P.; Nunez, V.; Fernandez, J.; Lomonte, B. Integrative characterization of the venom of the coral snake Micrurus dumerilii (Elapidae) from Colombia: Proteome, toxicity, and cross-neutralization by antivenom. J. Proteom. 2016, 136, 262–273. [Google Scholar] [CrossRef]
- Degueldre, M.; Echterbille, J.; Smargiasso, N.; Damblon, C.; Gouin, C.; Mourier, G.; Gilles, N.; De Pauw, E.; Quinton, L. In-Depth Glyco-Peptidomics Approach Reveals Unexpected Diversity of Glycosylated Peptides and Atypical Post-Translational Modifications in Dendroaspis angusticeps Snake Venom. Int. J. Mol. Sci. 2017, 18, 2453. [Google Scholar] [CrossRef]
- Petras, D.; Heiss, P.; Sussmuth, R.D.; Calvete, J.J. Venom Proteomics of Indonesian King Cobra, Ophiophagus hannah: Integrating Top-Down and Bottom-Up Approaches. J. Proteome Res. 2015, 14, 2539–2556. [Google Scholar] [CrossRef]
- Rodrigues, C.R.; Teixeira-Ferreira, A.; Vargas, F.F.R.; Guerra-Duarte, C.; Costal-Oliveira, F.; Stransky, S.; Lopes-de-Souza, L.; Dutra, A.A.A.; Yarleque, A.; Bonilla, C.; et al. Proteomic profile, biological activities and antigenic analysis of the venom from Bothriopsis bilineata smaragdina (“loro machaco”), a pitviper snake from Peru. J. Proteom. 2018, 187, 171–181. [Google Scholar] [CrossRef]
- Wiezel, G.A.; Shibao, P.Y.T.; Cologna, C.T.; Morandi Filho, R.; Ueira-Vieira, C.; De Pauw, E.; Quinton, L.; Arantes, E.C. In-Depth Venome of the Brazilian Rattlesnake Crotalus durissus terrificus: An Integrative Approach Combining Its Venom Gland Transcriptome and Venom Proteome. J. Proteome Res. 2018, 17, 3941–3958. [Google Scholar] [CrossRef]
- Fox, J.W.; Ma, L.; Nelson, K.; Sherman, N.E.; Serrano, S.M. Comparison of indirect and direct approaches using ion-trap and Fourier transform ion cyclotron resonance mass spectrometry for exploring viperid venom proteomes. Toxicon 2006, 47, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Vanuopadath, M.; Sajeev, N.; Murali, A.R.; Sudish, N.; Kangosseri, N.; Sebastian, I.R.; Jain, N.D.; Pal, A.; Raveendran, D.; Nair, B.G.; et al. Mass spectrometry-assisted venom profiling of Hypnale hypnale found in the Western Ghats of India incorporating de novo sequencing approaches. Int. J. Biol. Macromol. 2018, 118, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, M.; McCleary, R.J.R.; Kesherwani, M.; Kini, R.M.; Velmurugan, D. Comparison of proteomic profiles of the venoms of two of the ‘Big Four’ snakes of India, the Indian cobra (Naja naja) and the common krait (Bungarus caeruleus), and analyses of their toxins. Toxicon 2017, 135, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Kerkkamp, H.M.; Kini, R.M.; Pospelov, A.S.; Vonk, F.J.; Henkel, C.V.; Richardson, M.K. Snake Genome Sequencing: Results and Future Prospects. Toxins 2016, 8, 360. [Google Scholar] [CrossRef]
- Castoe, T.A.; de Koning, A.P.; Hall, K.T.; Card, D.C.; Schield, D.R.; Fujita, M.K.; Ruggiero, R.P.; Degner, J.F.; Daza, J.M.; Gu, W.; et al. The Burmese python genome reveals the molecular basis for extreme adaptation in snakes. Proc. Natl. Acad. Sci. USA 2013, 110, 20645–20650. [Google Scholar] [CrossRef]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.; Kerkkamp, H.M.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef]
- Dowell, N.L.; Giorgianni, M.W.; Kassner, V.A.; Selegue, J.E.; Sanchez, E.E.; Carroll, S.B. The Deep Origin and Recent Loss of Venom Toxin Genes in Rattlesnakes. Curr. Biol. 2016, 26, 2434–2445. [Google Scholar] [CrossRef]
- Suryamohan, K.; Krishnankutty, S.P.; Guillory, J.; Jevit, M.; Schroder, M.S.; Wu, M.; Kuriakose, B.; Mathew, O.K.; Perumal, R.C.; Koludarov, I.; et al. The Indian cobra reference genome and transcriptome enables comprehensive identification of venom toxins. Nat. Genet. 2020, 52, 106–117. [Google Scholar]
- Tattini, L.; D’Aurizio, R.; Magi, A. Detection of Genomic Structural Variants from Next-Generation Sequencing Data. Front. Bioeng. Biotechnol. 2015, 3, 92. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Z.J.; Li, Q.Y.; Lian, J.M.; Zhou, Y.; Lu, B.Z.; Jin, L.J.; Qiu, P.X.; Zhang, P.; Zhu, W.B.; et al. Evolutionary trajectories of snake genes and genomes revealed by comparative analyses of five-pacer viper. Nat. Commun. 2016, 7, 13107. [Google Scholar] [CrossRef]
- Shibata, H.; Chijiwa, T.; Oda-Ueda, N.; Nakamura, H.; Yamaguchi, K.; Hattori, S.; Matsubara, K.; Matsuda, Y.; Yamashita, A.; Isomoto, A.; et al. The habu genome reveals accelerated evolution of venom protein genes. Sci. Rep. 2018, 8, 11300. [Google Scholar] [PubMed]
- Pla, D.; Gutierrez, J.M.; Calvete, J.J. Second generation snake antivenomics: Comparing immunoaffinity and immunodepletion protocols. Toxicon 2012, 60, 688–699. [Google Scholar] [PubMed]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Antivenom for Neuromuscular Paralysis Resulting From Snake Envenoming. Toxins 2017, 9, 143. [Google Scholar]
- Pla, D.; Rodriguez, Y.; Calvete, J.J. Third Generation Antivenomics: Pushing the Limits of the In Vitro Preclinical Assessment of Antivenoms. Toxins 2017, 9, 158. [Google Scholar] [CrossRef]
- Ascoet, S.; De Waard, M. Diagnostic and Therapeutic Value of Aptamers in Envenomation Cases. Int. J. Mol. Sci. 2020, 21, 3565. [Google Scholar] [CrossRef] [PubMed]
- Lauridsen, L.H.; Shamaileh, H.A.; Edwards, S.L.; Taran, E.; Veedu, R.N. Rapid one-step selection method for generating nucleic acid aptamers: Development of a DNA aptamer against alpha-bungarotoxin. PLoS ONE 2012, 7, e41702. [Google Scholar] [CrossRef] [PubMed]
- Nimjee, S.M.; Rusconi, C.P.; Sullenger, B.A. Aptamers: An emerging class of therapeutics. Annu. Rev. Med. 2005, 56, 555–583. [Google Scholar] [CrossRef]
- Gragoudas, E.S.; Adamis, A.P.; Cunningham, E.T., Jr.; Feinsod, M.; Guyer, D.R. Pegaptanib for neovascular age-related macular degeneration. N. Engl. J. Med. 2004, 351, 2805–2816. [Google Scholar] [CrossRef]
- Alomran, N.; Chinnappan, R.; Alsolaiss, J.; Casewell, N.R.; Zourob, M. Exploring the Utility of ssDNA Aptamers Directed against Snake Venom Toxins as New Therapeutics for Snakebite Envenoming. Toxins 2022, 14, 469. [Google Scholar] [CrossRef]
- El-Aziz, T.M.A.; Ravelet, C.; Molgo, J.; Fiore, E.; Pale, S.; Amar, M.; Al-Khoury, S.; Dejeu, J.; Fadl, M.; Ronjat, M.; et al. Efficient functional neutralization of lethal peptide toxins in vivo by oligonucleotides. Sci. Rep. 2017, 7, 7202. [Google Scholar] [CrossRef]
- Chen, Y.J.; Tsai, C.Y.; Hu, W.P.; Chang, L.S. DNA Aptamers against Taiwan Banded Krait alpha-Bungarotoxin Recognize Taiwan Cobra Cardiotoxins. Toxins 2016, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, A.; Anand, A.; Malhotra, A.; Khan, E.; Santra, V.; Kumar, A.; Sharma, T.K. Rational truncation of aptamer for cross-species application to detect krait envenomation. Sci. Rep. 2018, 8, 17795. [Google Scholar]
- Taiwe, G.S.; Montnach, J.; Nicolas, S.; De Waard, S.; Fiore, E.; Peyrin, E.; El-Aziz, T.M.A.; Amar, M.; Molgo, J.; Ronjat, M.; et al. Aptamer Efficacies for In Vitro and In Vivo Modulation of alphaC-Conotoxin PrXA Pharmacology. Molecules 2019, 24, 229. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Sola, M.; Jappe, E.C.; Oscoz, S.; Lauridsen, L.P.; Engmark, M. Biotechnological Trends in Spider and Scorpion Antivenom Development. Toxins 2016, 8, 226. [Google Scholar]
- Ye, F.; Zheng, Y.; Wang, X.; Tan, X.; Zhang, T.; Xin, W.; Wang, J.; Huang, Y.; Fan, Q. Recognition of Bungarus multicinctus venom by a DNA aptamer against beta-bungarotoxin. PLoS ONE 2014, 9, e105404. [Google Scholar]
- Anand, A.; Chatterjee, B.; Dhiman, A.; Goel, R.; Khan, E.; Malhotra, A.; Santra, V.; Salvi, N.; Khadilkar, M.V.; Bhatnagar, I.; et al. Complex target SELEX-based identification of DNA aptamers against Bungarus caeruleus venom for the detection of envenomation using a paper-based device. Biosens. Bioelectron. 2021, 193, 113523. [Google Scholar] [CrossRef]
- Devi, A.; Doley, R. Neutralization of Daboxin P activities by rationally designed aptamers. Toxicon 2021, 203, 93–103. [Google Scholar]
- Muyldermans, S. Single domain camel antibodies: Current status. J. Biotechnol. 2001, 74, 277–302. [Google Scholar]
- Richard, G.; Meyers, A.J.; McLean, M.D.; Arbabi-Ghahroudi, M.; MacKenzie, R.; Hall, J.C. In vivo neutralization of alpha-cobratoxin with high-affinity llama single-domain antibodies (VHHs) and a VHH-Fc antibody. PLoS ONE 2013, 8, e69495. [Google Scholar] [CrossRef]
- Harrison, R.A.; Hasson, S.S.; Harmsen, M.; Laing, G.D.; Conrath, K.; Theakston, R.D. Neutralisation of venom-induced haemorrhage by IgG from camels and llamas immunised with viper venom and also by endogenous, non-IgG components in camelid sera. Toxicon 2006, 47, 364–368. [Google Scholar] [CrossRef]
- Leon, G.; Lomonte, B.; Gutierrez, J.M. Anticomplementary activity of equine whole IgG antivenoms: Comparison of three fractionation protocols. Toxicon 2005, 45, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Otero, R.; Gutierrez, J.M.; Rojas, G.; Nunez, V.; Diaz, A.; Miranda, E.; Uribe, A.F.; Silva, J.F.; Ospina, J.G.; Medina, Y.; et al. A randomized blinded clinical trial of two antivenoms, prepared by caprylic acid or ammonium sulphate fractionation of IgG, in Bothrops and Porthidium snake bites in Colombia: Correlation between safety and biochemical characteristics of antivenoms. Toxicon 1999, 37, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Lalloo, D.G.; Theakston, R.D. Snake antivenoms. J. Toxicol. Clin. Toxicol. 2003, 41, 277–290; 317–327. [Google Scholar]
- Leon, G.; Monge, M.; Rojas, E.; Lomonte, B.; Gutierrez, J.M. Comparison between IgG and F(ab′)(2) polyvalent antivenoms: Neutralization of systemic effects induced by Bothrops asper venom in mice, extravasation to muscle tissue, and potential for induction of adverse reactions. Toxicon 2001, 39, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.A.; Owen, T.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid antibodies for antivenom development: Neutralisation of venom-induced pathology. Toxicon 2010, 56, 373–380. [Google Scholar] [CrossRef]
- Darvish, M.; Ebrahimi, S.A.; Shahbazzadeh, D.; Bagheri, K.P.; Behdani, M.; Shokrgozar, M.A. Camelid antivenom development and potential in vivo neutralization of Hottentotta saulcyi scorpion venom. Toxicon 2016, 113, 70–75. [Google Scholar] [PubMed]
- Meddeb-Mouelhi, F.; Bouhaouala-Zahar, B.; Benlasfar, Z.; Hammadi, M.; Mejri, T.; Moslah, M.; Karoui, H.; Khorchani, T.; El Ayeb, M. Immunized camel sera and derived immunoglobulin subclasses neutralizing Androctonus australis hector scorpion toxins. Toxicon 2003, 42, 785–791. [Google Scholar] [CrossRef]
- Darvish, M.; Behdani, M.; Shokrgozar, M.A.; Pooshang-Bagheri, K.; Shahbazzadeh, D. Development of protective agent against Hottentotta saulcyi venom using camelid single-domain antibody. Mol. Immunol. 2015, 68, 412–420. [Google Scholar] [CrossRef]
- Rodriguez-Valle, M.; McAlister, S.; Moolhuijzen, P.M.; Booth, M.; Agnew, K.; Ellenberger, C.; Knowles, A.G.; Vanhoff, K.; Bellgard, M.I.; Tabor, A.E. Immunomic Investigation of Holocyclotoxins to Produce the First Protective Anti-Venom Vaccine Against the Australian Paralysis Tick, Ixodes holocyclus. Front. Immunol. 2021, 12, 744795. [Google Scholar] [CrossRef]
- Cook, D.A.; Owen, T.; Wagstaff, S.C.; Kinne, J.; Wernery, U.; Harrison, R.A. Analysis of camelid IgG for antivenom development: Serological responses of venom-immunised camels to prepare either monospecific or polyspecific antivenoms for West Africa. Toxicon 2010, 56, 363–372. [Google Scholar] [CrossRef]
- Tanwar, P.D.; Ghorui, S.K.; Kochar, S.K.; Singh, R.; Patil, N.V. Production and preclinical assessment of camelid immunoglobulins against Echis sochureki venom from desert of Rajasthan, India. Toxicon 2017, 134, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khamehchian, S.; Zolfagharian, H.; Dounighi, N.M.; Tebianian, M.; Madani, R. Study on camel IgG purification: A new approach to prepare Naja Naja oxiana antivenom as passive immunization for therapy. Hum. Vaccin. Immunother. 2014, 10, 1633–1638. [Google Scholar] [CrossRef]
- Bazan, J.; Calkosinski, I.; Gamian, A. Phage display—A powerful technique for immunotherapy: 1. Introduction and potential of therapeutic applications. Hum. Vaccin. Immunother. 2012, 8, 1817–1828. [Google Scholar] [PubMed]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Roncolato, E.C.; Campos, L.B.; Pessenda, G.; Costa e Silva, L.; Furtado, G.P.; Barbosa, J.E. Phage display as a novel promising antivenom therapy: A review. Toxicon 2015, 93, 79–84. [Google Scholar] [PubMed]
- Lynagh, T.; Kiontke, S.; Meyhoff-Madsen, M.; Gless, B.H.; Johannesen, J.; Kattelmann, S.; Christiansen, A.; Dufva, M.; Laustsen, A.H.; Devkota, K.; et al. Peptide Inhibitors of the alpha-Cobratoxin-Nicotinic Acetylcholine Receptor Interaction. J. Med. Chem. 2020, 63, 13709–13718. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Leu, S.J.; Lee, Y.C.; Liu, C.I.; Lin, L.T.; Mwale, P.F.; Chiang, J.R.; Tsai, B.Y.; Chen, C.C.; Hung, C.S.; et al. Characterization of Chicken-Derived Single Chain Antibody Fragments against Venom of Naja Naja Atra. Toxins 2018, 10, 383. [Google Scholar] [CrossRef]
- Lee, C.H.; Liu, C.I.; Leu, S.J.; Lee, Y.C.; Chiang, J.R.; Chiang, L.C.; Mao, Y.C.; Tsai, B.Y.; Hung, C.S.; Chen, C.C.; et al. Chicken antibodies against venom proteins of Trimeresurus stejnegeri in Taiwan. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200056. [Google Scholar] [CrossRef]
- Ahmadi, S.; Pucca, M.B.; Jurgensen, J.A.; Janke, R.; Ledsgaard, L.; Schoof, E.M.; Sorensen, C.V.; Caliskan, F.; Laustsen, A.H. An in vitro methodology for discovering broadly-neutralizing monoclonal antibodies. Sci. Rep. 2020, 10, 10765. [Google Scholar] [CrossRef]
- Ledsgaard, L.; Wade, J.; Jenkins, T.P.; Boddum, K.; Oganesyan, I.; Harrison, J.A.; Villar, P.; Leah, R.A.; Zenobi, R.; Schoffelen, S.; et al. Discovery and optimization of a broadly-neutralizing human monoclonal antibody against long-chain alpha-neurotoxins from snakes. Nat. Commun. 2023, 14, 682. [Google Scholar]
- Silva, L.C.; Pucca, M.B.; Pessenda, G.; Campos, L.B.; Martinez, E.Z.; Cerni, F.A.; Barbosa, J.E. Discovery of human scFvs that cross-neutralize the toxic effects of B. jararacussu and C. d. terrificus venoms. Acta Trop. 2018, 177, 66–73. [Google Scholar]
- Liu, C.C.; Wu, C.J.; Chou, T.Y.; Liaw, G.W.; Hsiao, Y.C.; Chu, L.J.; Lee, C.H.; Wang, P.J.; Hsieh, C.H.; Chen, C.K.; et al. Development of a Monoclonal scFv against Cytotoxin to Neutralize Cytolytic Activity Induced by Naja atra Venom on Myoblast C2C12 Cells. Toxins 2022, 14, 459. [Google Scholar] [CrossRef] [PubMed]
- Holliger, P.; Hudson, P.J. Engineered antibody fragments and the rise of single domains. Nat. Biotechnol. 2005, 23, 1126–1136. [Google Scholar] [PubMed]
- Alangode, A.; Reick, M. Sodium oleate, arachidonate, and linoleate enhance fibrinogenolysis by Russell’s viper venom proteinases and inhibit FXIIIa; a role for phospholipase A2 in venom induced consumption coagulopathy. Toxicon 2020, 186, 83–93. [Google Scholar]
- Rosenson, R.S.; Hislop, C.; Elliott, M.; Stasiv, Y.; Goulder, M.; Waters, D. Effects of varespladib methyl on biomarkers and major cardiovascular events in acute coronary syndrome patients. J. Am. Coll. Cardiol. 2010, 56, 1079–1088. [Google Scholar] [CrossRef]
- Lewin, M.; Samuel, S.; Merkel, J.; Bickler, P. Varespladib (LY315920) Appears to Be a Potent, Broad-Spectrum, Inhibitor of Snake Venom Phospholipase A2 and a Possible Pre-Referral Treatment for Envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef]
- Menzies, S.K.; Clare, R.H.; Xie, C.; Westhorpe, A.; Hall, S.R.; Edge, R.J.; Alsolaiss, J.; Crittenden, E.; Marriott, A.E.; Harrison, R.A.; et al. In vitro and in vivo preclinical venom inhibition assays identify metalloproteinase inhibiting drugs as potential future treatments for snakebite envenoming by Dispholidus typus. Toxicon X 2022, 14, 100118. [Google Scholar]
- Isbister, G.K. Procoagulant snake toxins: Laboratory studies, diagnosis, and understanding snakebite coagulopathy. Semin. Thromb. Hemost. 2009, 35, 93–103. [Google Scholar]
- Xie, C.; Albulescu, L.O.; Still, K.B.M.; Slagboom, J.; Zhao, Y.; Jiang, Z.; Somsen, G.W.; Vonk, F.J.; Casewell, N.R.; Kool, J. Varespladib Inhibits the Phospholipase A2 and Coagulopathic Activities of Venom Components from Hemotoxic Snakes. Biomedicines 2020, 8, 165. [Google Scholar] [CrossRef]
- Escalante, T.; Franceschi, A.; Rucavado, A.; Gutierrez, J.M. Effectiveness of batimastat, a synthetic inhibitor of matrix metalloproteinases, in neutralizing local tissue damage induced by BaP1, a hemorrhagic metalloproteinase from the venom of the snake bothrops asper. Biochem. Pharmacol. 2000, 60, 269–274. [Google Scholar]
- Gutierrez, J.M.; Leon, G.; Rojas, G.; Lomonte, B.; Rucavado, A.; Chaves, F. Neutralization of local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon 1998, 36, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Lay, M.; Liang, Q.; Isbister, G.K.; Hodgson, W.C. In Vitro Efficacy of Antivenom and Varespladib in Neutralising Chinese Russell’s Viper (Daboia siamensis) Venom Toxicity. Toxins 2023, 15, 62. [Google Scholar] [CrossRef] [PubMed]
- Layfield, H.J.; Williams, H.F.; Ravishankar, D.; Mehmi, A.; Sonavane, M.; Salim, A.; Vaiyapuri, R.; Lakshminarayanan, K.; Vallance, T.M.; Bicknell, A.B.; et al. Repurposing Cancer Drugs Batimastat and Marimastat to Inhibit the Activity of a Group I Metalloprotease from the Venom of the Western Diamondback Rattlesnake, Crotalus atrox. Toxins 2020, 12, 309. [Google Scholar] [CrossRef]
- Chowdhury, A.; Lewin, M.R.; Zdenek, C.N.; Carter, R.; Fry, B.G. The Relative Efficacy of Chemically Diverse Small-Molecule Enzyme-Inhibitors Against Anticoagulant Activities of African Spitting Cobra (Naja Species) Venoms. Front. Immunol. 2021, 12, 752442. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Lewin, M.R.; Williams, D.J.; Lomonte, B. Varespladib (LY315920) and Methyl Varespladib (LY333013) Abrogate or Delay Lethality Induced by Presynaptically Acting Neurotoxic Snake Venoms. Toxins 2020, 12, 131. [Google Scholar] [CrossRef]
- Tan, C.H.; Lingam, T.M.C.; Tan, K.Y. Varespladib (LY315920) rescued mice from fatal neurotoxicity caused by venoms of five major Asiatic kraits (Bungarus spp.) in an experimental envenoming and rescue model. Acta Trop. 2022, 227, 106289. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.W.; Gerardo, C.J.; Samuel, S.P.; Kumar, S.; Kotehal, S.D.; Mukherjee, P.P.; Shirazi, F.M.; Akpunonu, P.D.; Bammigatti, C.; Bhalla, A.; et al. The BRAVO Clinical Study Protocol: Oral Varespladib for Inhibition of Secretory Phospholipase A2 in the Treatment of Snakebite Envenoming. Toxins 2022, 15, 22. [Google Scholar] [CrossRef] [PubMed]
- Winer, A.; Adams, S.; Mignatti, P. Matrix Metalloproteinase Inhibitors in Cancer Therapy: Turning Past Failures Into Future Successes. Mol. Cancer Ther. 2018, 17, 1147–1155. [Google Scholar] [CrossRef]
- Shalinsky, D.R.; Brekken, J.; Zou, H.; Kolis, S.; Wood, A.; Webber, S.; Appelt, K. Antitumor efficacy of AG3340 associated with maintenance of minimum effective plasma concentrations and not total daily dose, exposure or peak plasma concentrations. Investig. New. Drugs 1998, 16, 303–313. [Google Scholar] [CrossRef]
- Shalinsky, D.R.; Brekken, J.; Zou, H.; Bloom, L.A.; McDermott, C.D.; Zook, S.; Varki, N.M.; Appelt, K. Marked antiangiogenic and antitumor efficacy of AG3340 in chemoresistant human non-small cell lung cancer tumors: Single agent and combination chemotherapy studies. Clin. Cancer Res. 1999, 5, 1905–1917. [Google Scholar]
- Gomez-Betancur, I.; Gogineni, V.; Salazar-Ospina, A.; Leon, F. Perspective on the Therapeutics of Anti-Snake Venom. Molecules 2019, 24, 3276. [Google Scholar] [CrossRef] [PubMed]
- Dhananjaya, B.L.; Shivalingaiah, S. The anti-inflammatory activity of standard aqueous stem bark extract of Mangifera indica L. as evident in inhibition of Group IA sPLA2. An. Acad. Bras. Cienc. 2016, 88, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Alangode, A.; Rajan, K.; Nair, B.G. Snake antivenom: Challenges and alternate approaches. Biochem. Pharmacol. 2020, 181, 114135. [Google Scholar] [CrossRef]
- Alam, M.I.; Gomes, A. Snake venom neutralization by Indian medicinal plants (Vitex negundo and Emblica officinalis) root extracts. J. Ethnopharmacol. 2003, 86, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Lohse, B.; Lomonte, B.; Engmark, M.; Gutierrez, J.M. Selecting key toxins for focused development of elapid snake antivenoms and inhibitors guided by a Toxicity Score. Toxicon 2015, 104, 43–45. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wagstaff, S.C.; Wuster, W.; Cook, D.A.; Bolton, F.M.; King, S.I.; Pla, D.; Sanz, L.; Calvete, J.J.; Harrison, R.A. Medically important differences in snake venom composition are dictated by distinct postgenomic mechanisms. Proc. Natl. Acad. Sci. USA 2014, 111, 9205–9210. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Lomonte, B. Phospholipases A2: Unveiling the secrets of a functionally versatile group of snake venom toxins. Toxicon 2013, 62, 27–39. [Google Scholar] [CrossRef]
- Bhattacharjee, P.; Bera, I.; Chakraborty, S.; Ghoshal, N.; Bhattacharyya, D. Aristolochic acid and its derivatives as inhibitors of snake venom L-amino acid oxidase. Toxicon 2017, 138, 1–17. [Google Scholar] [CrossRef]
- Vishwanath, B.S.; Gowda, T.V. Interaction of aristolochic acid with Vipera russelli phospholipase A2: Its effect on enzymatic and pathological activities. Toxicon 1987, 25, 929–937. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Doley, R.; Saikia, D. Isolation of a snake venom phospholipase A2 (PLA2) inhibitor (AIPLAI) from leaves of Azadirachta indica (Neem): Mechanism of PLA2 inhibition by AIPLAI in vitro condition. Toxicon 2008, 51, 1548–1553. [Google Scholar] [CrossRef]
- Venkatachalapathi, A.; Sangeeth, T.; Ali, M.A.; Tamilselvi, S.S.; Paulsamy, S.; Al-Hemaidc, F.M.A. Ethnomedicinal assessment of Irula tribes of Walayar valley of Southern Western Ghats, India. Saudi J. Biol. Sci. 2018, 25, 760–775. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.S.; Mohanakumari, H.P.; Nagaraju, S.; Vishwanath, B.S.; Kemparaju, K. Hyaluronidase and protease activities from Indian snake venoms: Neutralization by Mimosa pudica root extract. Fitoterapia 2004, 75, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekara, K.T.; Nagaraju, S.; Nandini, S.U.; Kemparaju, K. Neutralization of local and systemic toxicity of Daboia russelii venom by Morus alba plant leaf extract. Phytother. Res. 2009, 23, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Gopi, K.; Anbarasu, K.; Renu, K.; Jayanthi, S.; Vishwanath, B.S.; Jayaraman, G. Quercetin-3-O-rhamnoside from Euphorbia hirta protects against snake Venom induced toxicity. Biochim. Biophys. Acta 2016, 1860, 1528–1540. [Google Scholar] [CrossRef]
- Alam, M.I.; Auddy, B.; Gomes, A. Isolation, purification and partial characterization of viper venom inhibiting factor from the root extract of the Indian medicinal plant sarsaparilla (Hemidesmus indicus R. Br.). Toxicon 1994, 32, 1551–1557. [Google Scholar] [CrossRef]
- Alam, M.I.; Gomes, A. Viper venom-induced inflammation and inhibition of free radical formation by pure compound (2-hydroxy-4-methoxy benzoic acid) isolated and purified from anantamul (Hemidesmus indicus R. BR) root extract. Toxicon 1998, 36, 207–215. [Google Scholar] [CrossRef]
- Alam, M.I.; Gomes, A. An experimental study on evaluation of chemical antagonists induced snake venom neutralization. Indian J. Med. Res. 1998, 107, 142–146. [Google Scholar]
- Machiah, D.K.; Gowda, T.V. Purification of a post-synaptic neurotoxic phospholipase A2 from Naja naja venom and its inhibition by a glycoprotein from Withania somnifera. Biochimie 2006, 88, 701–710. [Google Scholar] [CrossRef]
- Deepa, M.; Veerabasappa Gowda, T. Purification and characterization of a glycoprotein inhibitor of toxic phospholipase from Withania somnifera. Arch. Biochem. Biophys. 2002, 408, 42–50. [Google Scholar] [CrossRef]
- Lizano, S.; Domont, G.; Perales, J. Natural phospholipase A2 myotoxin inhibitor proteins from snakes, mammals and plants. Toxicon 2003, 42, 963–977. [Google Scholar] [CrossRef]
- Chandra, V.; Jasti, J.; Kaur, P.; Srinivasan, A.; Betzel, C.; Singh, T.P. Structural basis of phospholipase A2 inhibition for the synthesis of prostaglandins by the plant alkaloid aristolochic acid from a 1.7 A crystal structure. Biochemistry 2002, 41, 10914–10919. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R., II; Francisco, A.F.; Moreira-Dill, L.S.; Quintero, A.; Guimaraes, C.L.S.; Fernandes, C.A.H.; Takeda, A.A.S.; Zanchi, F.B.; Caldeira, C.A.S.; Pereira, P.S.; et al. Isolation and structural characterization of bioactive compound from Aristolochia sprucei aqueous extract with anti-myotoxic activity. Toxicon X 2020, 7, 100049. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Antivenomics workflow. (a) Incubation of the antivenom with the matrix (sepharose or agarose beads). (b) Addition of crude venom. (c) Elution of unbound venom proteins. (d) Elution of bound venom proteins. (e) Resolve the bound and unbound fractions through orthogonal separation strategies (upper panel—SDS-PAGE analysis; bottom panel—reversed-phase HPLC analysis). (f) The collected fractions are then subjected to mass spectrometry-based proteomics for the identification of antivenom-bound and unbound proteins.
Figure 1.
Antivenomics workflow. (a) Incubation of the antivenom with the matrix (sepharose or agarose beads). (b) Addition of crude venom. (c) Elution of unbound venom proteins. (d) Elution of bound venom proteins. (e) Resolve the bound and unbound fractions through orthogonal separation strategies (upper panel—SDS-PAGE analysis; bottom panel—reversed-phase HPLC analysis). (f) The collected fractions are then subjected to mass spectrometry-based proteomics for the identification of antivenom-bound and unbound proteins.

Figure 2.
Whole Immunoglobuling IgG and single-chain (scFv) antibody fragment. Adapted from https://blog.addgene.org/antibodies-101-single-chain-fragment-variables-scfvs (accessed on 20 May 2023).
Figure 2.
Whole Immunoglobuling IgG and single-chain (scFv) antibody fragment. Adapted from https://blog.addgene.org/antibodies-101-single-chain-fragment-variables-scfvs (accessed on 20 May 2023).
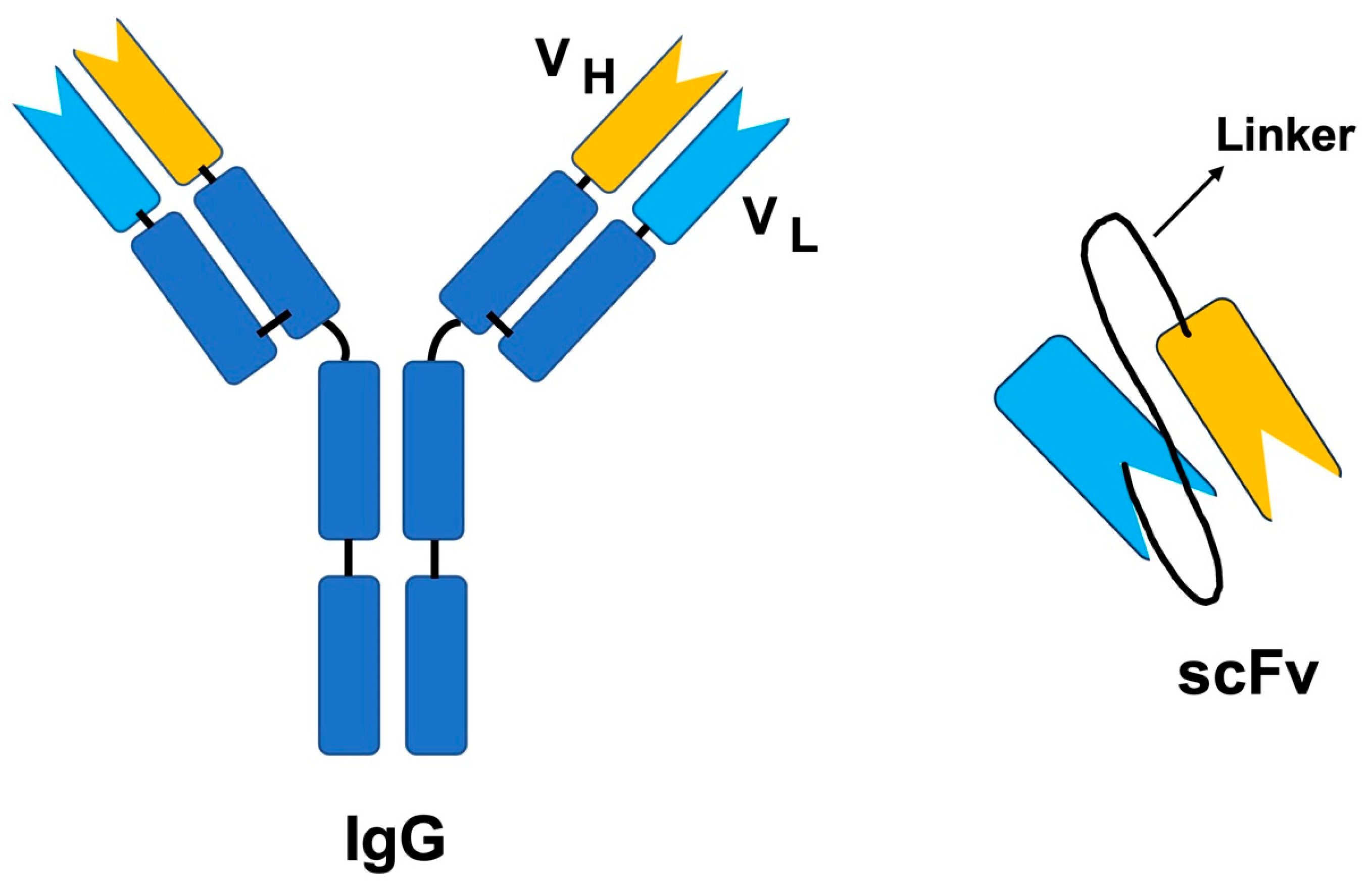
Figure 3.
Phage display with scFv on the phages are used to bind to the target antigen. High-binding phages are further amplifed to enrich these bound phages. Adapted from https://blog.addgene.org/antibodies-101-single-chain-fragment-variables-scfvs (accessed on 20 May 2023).
Figure 3.
Phage display with scFv on the phages are used to bind to the target antigen. High-binding phages are further amplifed to enrich these bound phages. Adapted from https://blog.addgene.org/antibodies-101-single-chain-fragment-variables-scfvs (accessed on 20 May 2023).

Figure 4.
The chemical structures of the small-molecule inhibitors: (A) varespladib, (B) batimastat, (C) marimastat, and (D) prinomastat. All chemical structures were drawn using ChemDraw® J S.
Figure 4.
The chemical structures of the small-molecule inhibitors: (A) varespladib, (B) batimastat, (C) marimastat, and (D) prinomastat. All chemical structures were drawn using ChemDraw® J S.
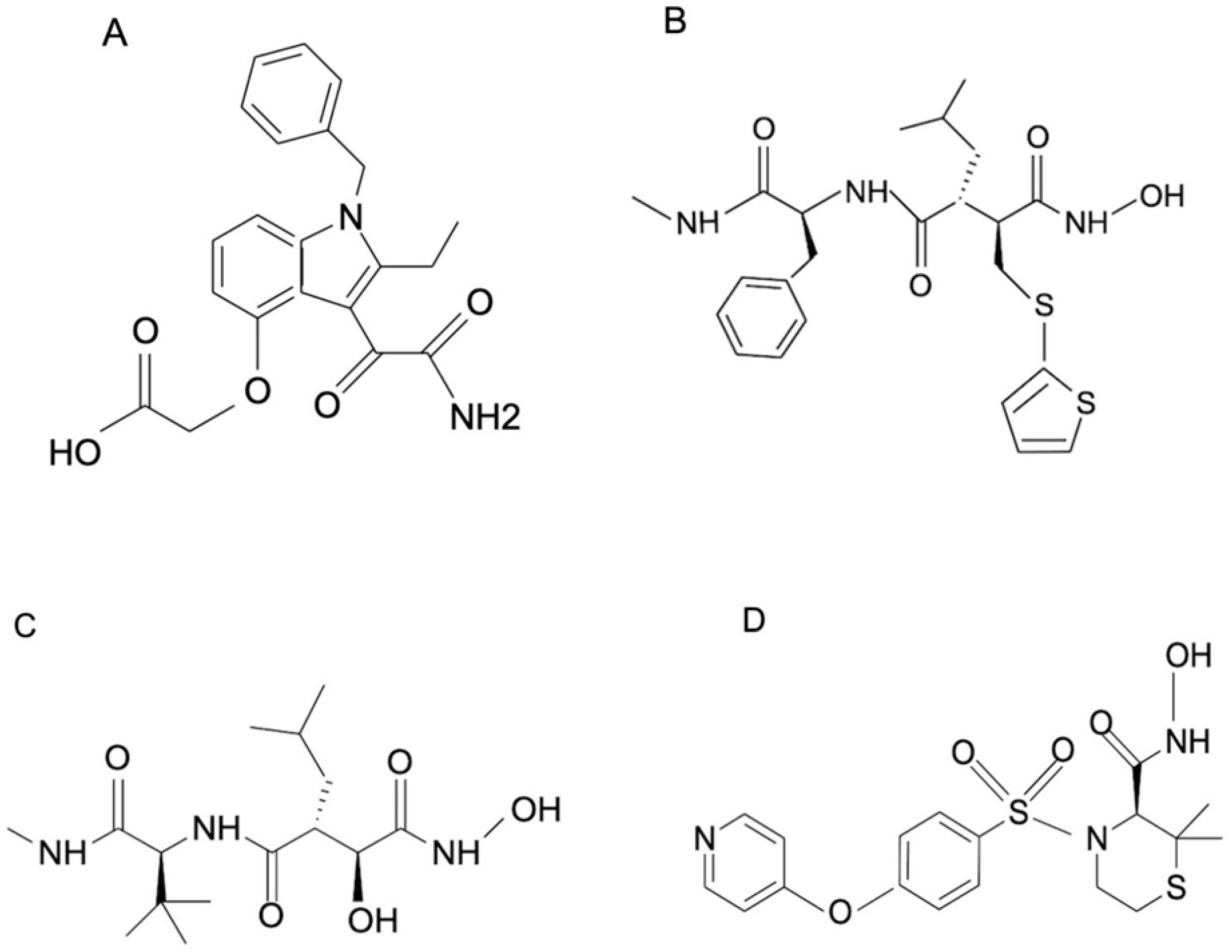
Table 1.
Medically important snake species of India and antivenom availability (Retrieved from the Snakebite Data Information portal—World Health Organization) [5]. Categories 1 and 2 indicate highly venomous snakes with the highest medical importance and secondary medical importance, respectively.
Table 1.
Medically important snake species of India and antivenom availability (Retrieved from the Snakebite Data Information portal—World Health Organization) [5]. Categories 1 and 2 indicate highly venomous snakes with the highest medical importance and secondary medical importance, respectively.
| SI NO. | Taxonomic Family | Species Name | Common Name | Geographical Distribution in India | Antivenom Available and Licensed in India | Nature of Available Antivenom (s) | Antivenom Manufacturing Countries |
|---|---|---|---|---|---|---|---|
| Category-1 | |||||||
| 1 | Elapidae | Bungarus caeruleus | Indian krait | Throughout | Yes | Polyvalent antivenom | India, Pakistan |
| 2 | Naja naja | Indian cobra | Throughout | Yes | Polyvalent antivenom | India, Pakistan | |
| 3 | Naja kaouthia | Monocellate cobra | Northeast | No | Monovalent and Polyvalent | Myanmar, Thailand, Vietnam | |
| 4 | Viperidae | Daboia russelii | Russell’s viper | Throughout | Yes | Polyvalent antivenom | India, Pakistan |
| 5 | Echis carinatus | Saw-scaled viper | Throughout | Yes | Polyvalent antivenom | India, Pakistan, Iran, Uzbekistan, Spain | |
| 6 | Hypnale hypnale | Hump-nosed pit viper | Southwest | No | NA | - | |
| Category-2 | |||||||
| 7 | Elapidae | Bungarus bungaroides | Northeastern hill krait | Northeast | No | NA | - |
| 8 | Bungarus fasciatus | Banded krait | Northeast | No | Polyvalent and monovalent antivenoms | Thailand, Indonesia | |
| 9 | Bungarus lividus | Lesser black krait | Northeast | No | NA | - | |
| 10 | Bungarus niger | Greater black krait | Northeast | No | NA | - | |
| 11 | Bungarus sindanus | Sind krait | Northwest | No | Polyvalent antivenom | Pakistan | |
| 12 | Bungarus walli | Wall’s krait | Northeast and Southwest | No | NA | - | |
| 13 | Naja oxiana | Central Asian cobra | North and Northwest | No | Polyvalent | Iran, Pakistan, Uzbekistan, Egypt | |
| 14 | Naja sagittifera | Andaman cobra | Andaman Islands | No | NA | - | |
| 15 | Ophiophagus hannah | King cobra | South, Northeast, Andaman Islands | No | Polyvalent and monovalent | Thailand | |
| 16 | Viperidae | Gloydius himalayanus | Himalayan pit viper | North | No | NA | - |
| 17 | Protobothrops jerdonii | Jerdon’s pit viper | Northeast | No | NA | - | |
| 18 | Protobothrops kaulbacki | Kaulback’s lance-headed pit viper | Northeast | No | NA | - | |
| 19 | Protobothrops mucrosquamatus | Brown-spotted pit viper | Northeast | No | Monovalent antivenom | China | |
| 20 | Trimeresurus gramineus | Common bamboo pit viper | South and East | No | NA | - | |
| 21 | Craspedocephalus malabaricus | Malabarian pit viper | Southwest | No | NA | - | |
| 22 | Macrovipera lebetina | Levantine viper | Northeast | No | |||
| 23 | Protobothrops himalayanus | NA | North | No | NA | - | |
| 24 | Trimeresurus andersonii | Andaman Pitviper | Andaman Islands | No | NA | - | |
| 25 | Trimeresurus erythrurus | Bamboo pitviper | East | No | Monovalent | Thailand | |
| 26 | Trimeresurus gumprechti | Gumprecht’s green pit viper | Northeast | No | NA | - | |
| 27 | Craspedocephalus macrolepis | large-scaled pit viper | South | No | NA | - | |
| 28 | Trimeresurus salazar | Salazar’s pit viper | Northeast | No | NA | - | |
| 29 | Trimeresurus septentrionalis | Nepal pitviper | North | No | NA | - | |
| 30 | Craspedocephalus strigatus | horseshoe pit viper | South | No | NA | - | |
| 31 | Trimeresurus yunnanensis | Yunnan bamboo pitviper | North | No | NA | - | |
NA—Not available.
Table 2.
List of antivenom manufacturers in India.
| SI No. | Name | Antivenom Manufacturer | Listed in WHO Database | Nature of Antivenom | Stated Efficacy |
|---|---|---|---|---|---|
| 1 | Polyvalent Snake Antivenin | Biological E Limited (Telangana, Hyderabad) | Yes | Polyvalent (raised against big four Indian snakes) | After reconstitution, each mL of Polyvalent snake venom antiserum neutralizes not less than: Indian Cobra venom—0.60 mg Common Krait venom—0.45 mg Russell’s viper venom—0.60 mg Saw scaled viper venom—0.45 mg (The stated efficacies of each of these antivenoms are indicated based on the LD50 and ED50 values obtained after performing In vivo studies) |
| 2 | Snake Venom Antiserum I.P. | VINS Bioproducts Ltd. (Telangana, Hyderabad) | Yes | ||
| 3 | Polyvalent Snake Antivenom | Bharat Serums & Vaccines (Mumbai, Maharashtra) | Yes | ||
| 4 | Snake antivenin I.P. | Haffkine Biopharmaceutical Corporation Ltd. (Mumbai, Maharashtra) | Yes | ||
| 5 | Virchow (V-ASV) | Virchow biotech private limited (Telangana, Hyderabad) | No | ||
| 6 | Snake Venom Antiserum I.P. | Premium Serums & Vaccines Pvt. Ltd. (Pune, Maharashtra) | No | ||
| 7 | Snake Venom Antiserum I.P. | Mediclone Biotech (Chennai, Tamil Nadu) | No | ||
| 8 | Polyvalent Anti Snake Venom Serum I.P. | King Institute of Preventative Medicine and Research (Chennai, Tamil Nadu) | Yes |
Table 4.
Studies on using aptamers to treat snakebite envenomation.
| SI No. | Species | Protein | Protein Family | Nature of Aptamer | Methodology Used for Selecting Aptamer | Efficacy/Efficiency | Reference |
|---|---|---|---|---|---|---|---|
| 1 | Daboia russelii | Daboxin P | Phospholipase A2 | Nucleic acid aptamer | Entropy fragment-based approach and seed and grow method | Showed PLA2 inhibitory and anticoagulant activities. | [101] |
| 2 | Bungarus multicinctus | β-bungarotoxin (β-BuTx) | Neurotoxin (three-finger toxin) | DNA aptamer | plate-SELEX | The designed aptamer βB-1 was specific to β-BuTx and could differentiate B. multicinctus venom among the other snake venoms tested. | [99] |
| 3 | Bungarus caeruleus | α-Toxin | Neurotoxin (three-finger toxin) | Truncated aptamer | Truncated aptamer | A truncated DNA aptamer, α-Tox-T2, generated to fight against the α-Toxin of Bungarus multicinctus was also able to detect Bungarus caeruleus venom. | [96] |
| 4 | Bungarus multicinctus | α-bungarotoxin | Neurotoxin (three-finger toxin) | DNA aptamer | Single-step selection on a glass coverslip using designed aptamers | Simple one-step selection could be applied for the rapid production of DNA and RNA aptamers. | [90] |
| 5 | N. atra | Cardiotoxins | Neurotoxin (three-finger toxin) | DNA aptamer | Neogene Biomedicals Corporation synthesized the labeled single-stranded DNA samples | The aptamers designed to fight against Bungarus multicinctus α-bungarotoxin inhibited cytotoxicity and membrane damage induced by Naja atra cardiotoxins. | [95] |
| 6 | Bungarus caeruleus | β-Bungarotoxin | Neurotoxin (three-finger toxin) | DNA aptamer | SELEX | The designed aptamer could discriminate B. caeruleus venom from Russell’s, Cobra, and Saw-scaled viper’s venom and was specific to β-Bungarotoxin. | [100] |
| 7 | C. rhodostoma and B. atrox | Ancrod and batroxobin | Snake venom serine protease | ssDNA aptamers | SELEX | The toxin-specific aptamers were found to exhibit in vitro cross-reactivity against the different isoforms present in various snake species. | [93] |
Table 5.
Studies on developing alternative treatment strategies using phage display.
| SI No. | Snake Species | Neutralizing Toxin/Protein (Antigen) | Phage Bound Molecule | Study Design | Reference |
|---|---|---|---|---|---|
| 1 | Naja kaouthia | α−cobratoxin | 8-mer peptide | In vitro | [120] |
| 2 | Naja naja atra | Naja naja atra proteins (NNA proteins) | Single-chain variable fragment (scFv) | In vitro and in vivo | [121] |
| 3 | Trimeresurus stejnegeri | Whole venom | scFv | In vitro and in vivo | [122] |
| 4 | Naja. nigricollis, Naja. mossambica, and Naja. melanoleuca | Whole venom | scFv | In vitro | [123] |
| 5 | Naja kaouthia | Whole venom | Monoclonal antibody | In vitro and in vivo | [124] |
| 6 | Bothrops jararacussu and Crotalus durissus terrificus | Whole venom | scFv | In vitro and in vivo | [125] |
Table 7.
Studies on natural product-based inhibitors against snake venom.
| SI No. | Species | Plant Extract/Compound | Protein Family | Nature of Plant Product | Neutralization Studies | Reference |
|---|---|---|---|---|---|---|
| 1 | Russell’s viper | Aristolochic acid | Phospholipase A2 L-amino acid Oxidase | Nitrophenanthrene carboxylic acids | In vitro cell based assays and in vivo mouse model | [101] |
| 2 | Daboia russelli, Naja naja, Naja kaouthia | Leaf extract of Azadirachta indica | Phospholipase A2 | Non- terpenoids | In vitro assays and in vivo mouse model | [99] |
| 3 | Daboia russelli, Naja naja, Echis carinatus | Root extract of Mimosa pudica | Hyaluronidase and Protease | Not determined | In vitro assays | [96] |
| 4 | Daboia russelli | Leaf extract of Morus alba | Snake venom metalloproteinase and hyaluronidase | Not determined | In vitro assays and in vivo mouse model | [90] |
| 5 | Naja naja | Quercetin-3-O-α-rhamnoside | Phospholipase A2, hyaluronidase and hemolytic activity | Flavonoid glycoside | In vitro assays and in vivo mouse model | [95] |
| 6 | Naja kaouthia, Ophiophagus hannah, Daboia russelii, and Echis carinatus | 2-hydroxy-4-methoxy benzoic acid | Phospholipase A2, snake venom metalloproteinase | Salicylates derivative | In vitro assays and in vivo mouse model | [100] |
| 7 | Naja naja | Withania sominifera Glycoprotein | Phospholipase A2 | Glycoprotein | In vitro assays and in vivo mouse model | [93] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vanuopadath, M.; Rajan, K.; Alangode, A.; Nair, S.S.; Nair, B.G. The Need for Next-Generation Antivenom for Snakebite Envenomation in India. Toxins 2023, 15, 510. https://doi.org/10.3390/toxins15080510
AMA Style
Vanuopadath M, Rajan K, Alangode A, Nair SS, Nair BG. The Need for Next-Generation Antivenom for Snakebite Envenomation in India. Toxins. 2023; 15(8):510. https://doi.org/10.3390/toxins15080510
Chicago/Turabian StyleVanuopadath, Muralidharan, Karthika Rajan, Aswathy Alangode, Sudarslal Sadasivan Nair, and Bipin Gopalakrishnan Nair. 2023. "The Need for Next-Generation Antivenom for Snakebite Envenomation in India" Toxins 15, no. 8: 510. https://doi.org/10.3390/toxins15080510
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.