Cationicity Enhancement on the Hydrophilic Face of Ctriporin Significantly Reduces Its Hemolytic Activity and Improves the Antimicrobial Activity against Antibiotic-Resistant ESKAPE Pathogens

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Analogue Design of Ctriporin
2.2. CM5 and CM6 Exhibited the Most Potent Antimicrobial Activity and the Lowest Hemolytic Activity
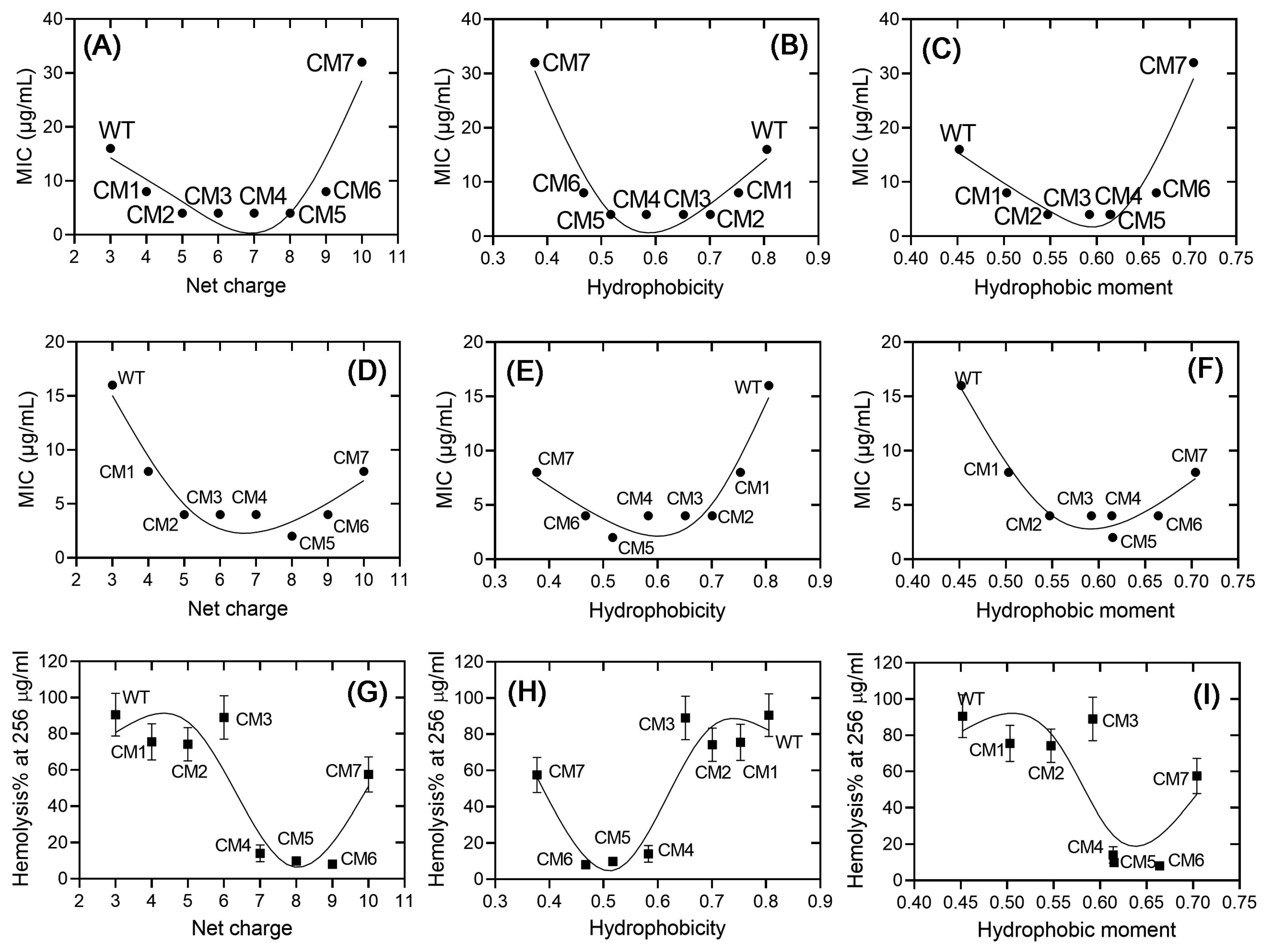
2.3. The Correlations between the Physiochemical Properties and the Activities of Ctriporin and Its Analogs
2.4. Antimicrobial Activities of CM5 and CM6 against Clinical Isolates of ESKAPE Pathogens
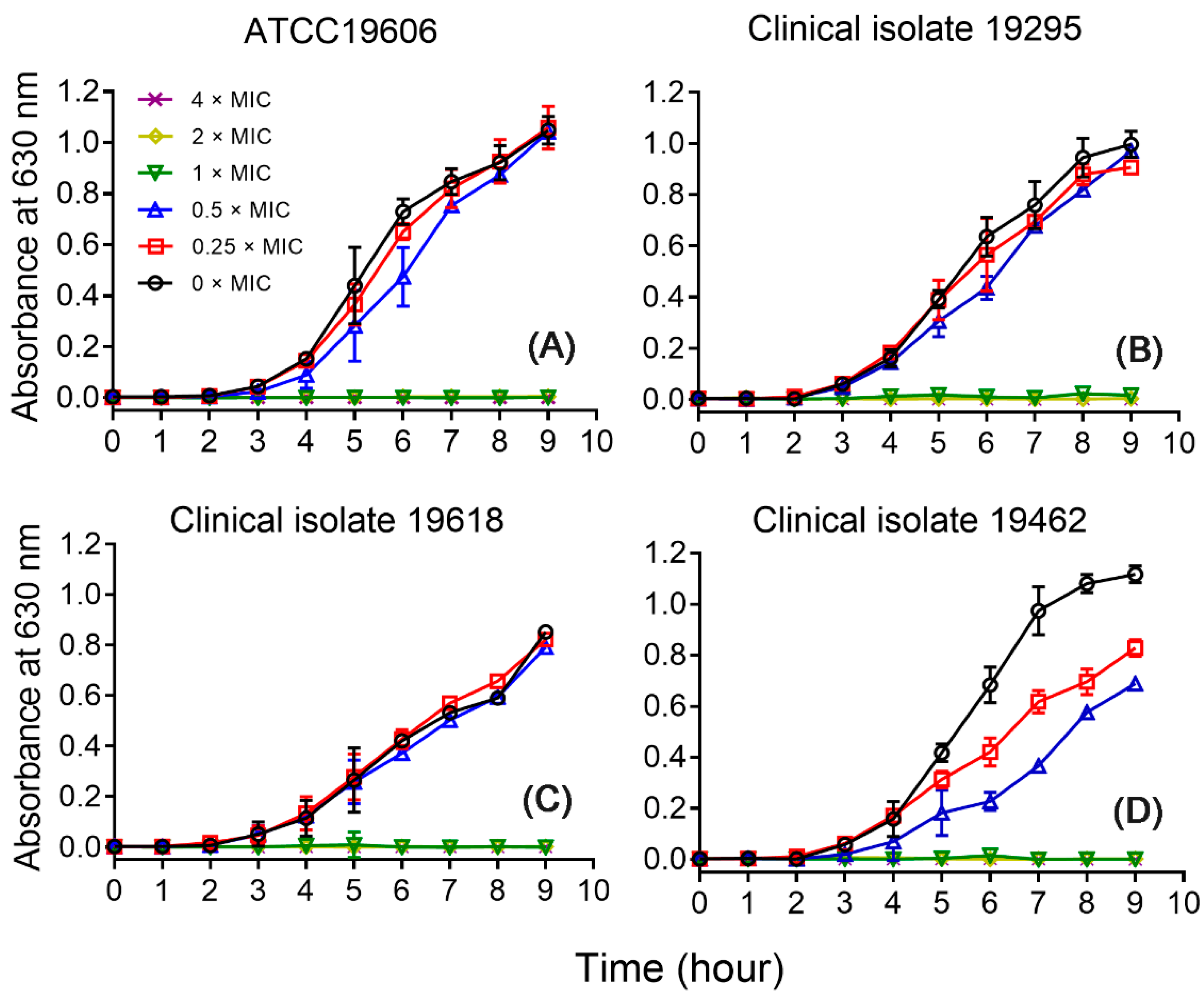
2.5. Growth Inhibitory Effects of CM5
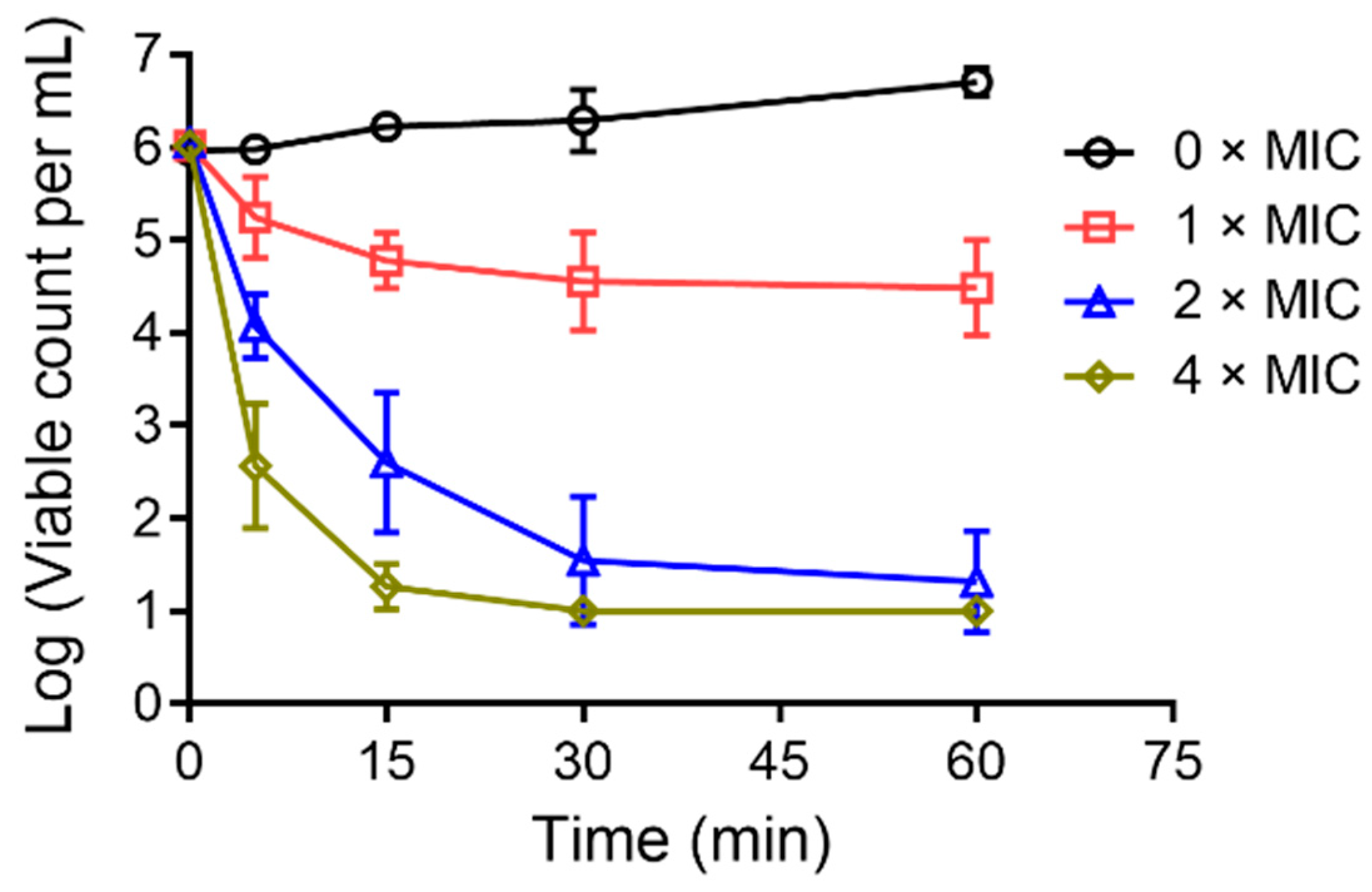
2.6. Bacterial-Killing Kinetics
2.7. CM5 Induces Dose-Dependent Membrane Disruptions of the Bacterial Cells
3. Discussion
4. Materials and Methods
4.1. Peptide Analysis and Synthesis
4.2. Circular Dichroism
4.3. Bacterial Strains
4.4. Minimum Inhibitory Concentration (MIC) Determination
4.5. Hemolytic Activity Determination
4.6. Growth Curve
4.7. Time-Killing Kinetics
4.8. Membrane Permeabilization Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti. Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Hu, F.; Zhu, D.; Wang, F.; Wang, M. Current status and trends of antibacterial resistance in China. Clin. Infect. Dis. 2018, 67 (Suppl. S2), S128–S134. [Google Scholar] [CrossRef]
- Oliveira, D.M.P.D.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walkera, M.J. Antimicrobial resistance in ESKAPE pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Harrison, P.L.; Abdel-Rahman, M.A.; Miller, K.; Strong, P.N. Antimicrobial peptides from scorpion venoms. Toxicon 2014, 88, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A. Diversity, evolution and medical applications of insect antimicrobial peptides. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150290. [Google Scholar] [CrossRef] [PubMed]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci. 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [PubMed]
- de Barros, E.; Goncalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Candido, E.S. Snake venom cathelicidins as natural antimicrobial peptides. Front. Pharmacol. 2019, 10, 1415. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, L.D.; Romanelli, A. Temporins: Multifunctional peptides from frog skin. Int. J. Mol. Sci. 2023, 24, 5426. [Google Scholar] [CrossRef] [PubMed]
- Rincón-Cortés, C.A.; Bayona-Rojas, M.A.; Reyes-Montaño, E.A.; Vega-Castro, N.A. Antimicrobial activity developed by scorpion venoms and its peptide component. Toxins 2022, 14, 740. [Google Scholar] [CrossRef]
- Ciumac, D.; Gong, H.; Hu, X.; Lu, J.R. Membrane targeting cationic antimicrobial peptides. J. Colloid Interface Sci. 2019, 537, 163–185. [Google Scholar] [CrossRef]
- Lee, T.H.; Hall, K.N.; Aguilar, M.I. Antimicrobial peptide structure and mechanism of action: A focus on the role of membrane structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Kmeck, A.; Tancer, R.J.; Ventura, C.R.; Wiedman, G.R. Synergies with and resistance to membrane-active peptides. Antibiotics 2020, 9, 620. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends Microbiol. 2019, 27, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez de la Vega, R.C.; Schwartz, E.F.; Possani, L.D. Mining on scorpion venom biodiversity. Toxicon 2010, 56, 1155–1161. [Google Scholar] [CrossRef]
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef]
- Fan, Z.; Cao, L.; He, Y.; Hu, J.; Di, Z.; Wu, Y.; Li, W.; Cao, Z. Ctriporin, a new anti-methicillin-resistant Staphylococcus aureus peptide from the venom of the scorpion Chaerilus tricostatus. Antimicrob. Agents Chemother. 2011, 55, 5220–5229. [Google Scholar] [CrossRef]
- Dai, C.; Ma, Y.; Zhao, Z.; Zhao, R.; Wang, Q.; Wu, Y.; Cao, Z.; Li, W. Mucroporin, the first cationic host defense peptide from the venom of Lychas mucronatus. Antimicrob. Agents Chemother. 2008, 52, 3967–3972. [Google Scholar] [CrossRef]
- Zhao, Z.H.; Ma, Y.B.; Dai, C.; Zhao, R.M.; Li, S.R.; Wu, Y.L.; Cao, Z.J.; Li, W.X. Imcroporin, a new cationic antimicrobial peptide from the venom of the scorpion Isometrus maculates. Antimicrob. Agents Chemother. 2009, 53, 3472–3477. [Google Scholar] [CrossRef]
- de Melo, E.T.; Estrela, A.B.; Santos, E.C.; Machado, P.R.; Farias, K.J.; Torres, T.M.; Carvalho, E.; Lima, J.P.; Silva-Junior, A.A.; Barbosa, E.G.; et al. Structural characterization of a novel peptide with antimicrobial activity from the venom gland of the scorpion Tityus stigmurus: Stigmurin. Peptides 2015, 68, 3–10. [Google Scholar] [CrossRef]
- Guo, X.; Ma, C.; Du, Q.; Wei, R.; Wang, L.; Zhou, M.; Chen, T.; Shaw, C. Two peptides, TsAP-1 and TsAP-2, from the venom of the Brazilian yellow scorpion, Tityus serrulatus: Evaluation of their antimicrobial and anticancer activities. Biochimie 2013, 95, 1784–1794. [Google Scholar] [CrossRef]
- Almaaytah, A.; Zhou, M.; Wang, L.; Chen, T.; Walker, B.; Shaw, C. Antimicrobial/cytolytic peptides from the venom of the North African scorpion, Androctonus amoreuxi: Biochemical and functional characterization of natural peptides and a single site-substituted analog. Peptides 2012, 35, 291–299. [Google Scholar] [CrossRef]
- Luo, F.; Zeng, X.C.; Hahin, R.; Cao, Z.J.; Liu, H.; Li, W.X. Genomic organization of four novel nondisulfide-bridged peptides from scorpion Mesobuthus martensii Karsch: Gaining insight into evolutionary mechanism. Peptides 2005, 26, 2427–2433. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, K.; Zhu, W.; Ye, X.; Ding, L.; Jiang, H.; Li, F.; Chen, Z.; Luo, X. Two new cationic α-helical peptides identified from the venom gland of Liocheles australasiae possess antimicrobial activity against methicillin-resistant staphylococci. Toxicon 2021, 196, 63–73. [Google Scholar] [CrossRef]
- Luo, X.; Ye, X.; Ding, L.; Zhu, W.; Zhao, Z.; Luo, D.; Liu, N.; Sun, L.; Chen, Z. Identification of the scorpion venom-derived antimicrobial peptide Hp1404 as a new antimicrobial agent against carbapenem-resistant Acinetobacter baumannii. Microb. Pathog. 2021, 157, 104960. [Google Scholar] [CrossRef] [PubMed]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific α-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.; Villegas, E.; Satake, H.; Possani, L.D.; Corzo, G. Amino acid substitutions in an α-helical antimicrobial arachnid peptide affect its chemical properties and biological activity towards pathogenic bacteria but improves its therapeutic index. Amino Acids 2011, 40, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.D.T.; Sothiselvam, S.; Lu, T.K.; de la Fuente-Nunez, C. Peptide design principles for antimicrobial applications. J. Mol. Biol. 2019, 431, 3547–3567. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Shao, W.M.; Chen, G.M. Autoencoder-based nonlinear Bayesian locally weighted regression for soft sensor development. ISA Trans. 2020, 103, 143–155. [Google Scholar] [CrossRef]
- Karl, A.S.; Steel, J.G.; Warr, G.B. Regression fitting megavoltage depth dose curves to determine material relative electron density in radiotherapy. Phys. Eng. Sci. Med. 2023, 46, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yang, F.; Li, F.; Li, Z.; Lang, Y.; Shen, B.; Wu, Y.; Li, W.; Harrison, P.L.; Strong, P.N.; et al. Therapeutic potential of a scorpion venom-derived antimicrobial peptide and its homologs against antibiotic-resistant gram-positive bacteria. Front. Microbiol. 2018, 9, 1159. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Lakshmaiah Narayana, J.; Lushnikova, T.; Wang, X.; Wang, G. Low cationicity is important for systemic in vivo efficacy of database-derived peptides against drug-resistant gram-positive pathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 13517–13522. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Zhang, L.S.; He, Y.H.; Liu, K.X.; Zhang, F.F.; Zhang, H.R.; Lu, Y.Q.; Yang, C.Y.; Wang, Z.P.; Fareed, M.S.; et al. An optimized analog of antimicrobial peptide Jelleine-1 shows enhanced antimicrobial activity against multidrug resistant and negligible toxicity in vitro and in vivo. Eur. J. Med. Chem. 2021, 219, 113433. [Google Scholar] [CrossRef] [PubMed]
- Liscano, Y.; Salamanca, C.H.; Vargas, L.; Cantor, S.; Laverde-Rojas, V.; Oñate-Garzón, J. Increases in hydrophilicity and charge on the polar face of alyteserin 1c helix change its selectivity towards gram-positive bacteria. Antibiotics 2019, 8, 238. [Google Scholar] [CrossRef]
- Mourtada, R.; Herce, H.D.; Yin, D.J.; Moroco, J.A.; Wales, T.E.; Engen, J.R.; Walensky, L.D. Design of stapled antimicrobial peptides that are stable, nontoxic and kill antibiotic-resistant bacteria in mice. Nat. Biotechnol. 2019, 37, 1186–1197. [Google Scholar] [CrossRef]
- Jiang, M.; Yang, X.; Wu, H.; Huang, Y.; Dou, J.; Zhou, C.; Ma, L. An active domain HF-18 derived from hagfish intestinal peptide effectively inhibited drug-resistant bacteria in vitro/vivo. Biochem. Pharmacol. 2020, 172, 113746. [Google Scholar] [CrossRef]
- Stone, T.A.; Cole, G.B.; Ravamehr-Lake, D.; Nguyen, H.Q.; Khan, F.; Sharpe, S.; Deber, C.M. Positive charge patterning and hydrophobicity of membrane-active antimicrobial peptides as determinants of activity, toxicity, and pharmacokinetic stability. J. Med. Chem. 2019, 62, 6276–6286. [Google Scholar] [CrossRef]
- Tan, Y.N.; Chen, X.L.; Ma, C.B.; Xi, X.P.; Wang, L.; Zhou, M.; Burrows, J.F.; Kwok, H.F.; Chen, T.B. Biological activities of cationicity-enhanced and hydrophobicity-optimized analogues of an antimicrobial peptide, dermaseptin-ps3, from the skin secretion of Phyllomedusa sauvagii. Toxins 2018, 10, 320. [Google Scholar] [CrossRef]
- Pedron, C.N.; Torres, M.T.; Lima, J.; Silva, P.I.; Silva, F.D.; Oliveira, V.X. Novel designed VmCT1 analogs with increased antimicrobial activity. Eur. J. Med. Chem. 2017, 126, 456–463. [Google Scholar] [CrossRef]
- Oliveira, C.S.; Torres, M.T.; Pedron, C.N.; Andrade, V.B.; Silva, P.I., Jr.; Silva, F.D.; de la Fuente-Nunez, C.; Oliveira, V.X., Jr. Synthetic peptide derived from scorpion venom displays minimal toxicity and anti-infective activity in an animal model. ACS Infect. Dis. 2021, 7, 2736–2745. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Ye, X.; Ding, L.; Zhu, W.; Yi, P.; Zhao, Z.; Gao, H.; Shu, Z.; Li, S.; Sang, M.; et al. Fine-tuning of alkaline residues on the hydrophilic face provides a non-toxic cationic alpha-helical antimicrobial peptide against antibiotic-resistant ESKAPE pathogens. Front. Microbiol. 2021, 12, 684591. [Google Scholar]
- Cao, L.; Dai, C.; Li, Z.; Fan, Z.; Song, Y.; Wu, Y.; Cao, Z.; Li, W. Antibacterial activity and mechanism of a scorpion venom peptide derivative in vitro and in vivo. PLoS ONE 2012, 7, e40135. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Vasil, A.I.; Hale, J.D.; Hancock, R.E.; Vasil, M.L.; Hodges, R.S. Effects of net charge and the number of positively charged residues on the biological activity of amphipathic α-helical cationic antimicrobial peptides. Biopolymers 2008, 90, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Li, Z.; Zhang, R.; Wu, Y.; Li, W.; Cao, Z. StCT2, a new antibacterial peptide characterized from the venom of the scorpion Scorpiops tibetanus. Peptides 2012, 36, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.C.; Zhou, L.; Shi, W.; Luo, X.; Zhang, L.; Nie, Y.; Wang, J.; Wu, S.; Cao, B.; Cao, H. Three new antimicrobial peptides from the scorpion Pandinus imperator. Peptides 2013, 45, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, X.; Meng, L.; Zhang, Q.; Cao, L.; Li, W.; Wu, Y.; Cao, Z. Hp1404, a new antimicrobial peptide from the scorpion Heterometrus petersii. PLoS ONE 2014, 9, e97539. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, I.C.C.; Silva, P.I., Jr.; Silva, F.D.; Araujo, I.; Alves, F.L.; Oliveira, C.S.; Oliveira, V.X., Jr. IsCT-based analogs intending better biological activity. J. Pept. Sci. 2019, 25, e3219. [Google Scholar] [CrossRef]
- Ding, L.; Hao, J.; Luo, X.; Zhu, W.; Wu, Z.; Qian, Y.; Hu, F.; Liu, T.; Ruan, X.; Li, S.; et al. The Kv1.3 channel-inhibitory toxin BF9 also displays anticoagulant activity via inhibition of factor XIa. Toxicon 2018, 152, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Saranathan, R.; Vasanth, V.; Vasanth, T.; Shabareesh, P.R.; Shashikala, P.; Devi, C.S.; Kalaivani, R.; Asir, J.; Sudhakar, P.; Prashanth, K. Emergence of carbapenem non-susceptible multidrug resistant Acinetobacter baumannii strains of clonal complexes 103(B) and 92(B) harboring OXA-type carbapenemases and metallo-β-lactamases in Southern India. Microbiol. Immunol. 2015, 59, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.M.; Edwards, M.A.; Li, J.; Yip, C.M.; Deber, C.M. Roles of hydrophobicity and charge distribution of cationic antimicrobial peptides in peptide-membrane interactions. J. Biol. Chem. 2012, 287, 7738–7745. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, G.; Meng, L.; Yu, W.; Xu, X.; Li, W.; Wu, Y.; Cao, Z. K1K8: An Hp1404-derived antibacterial peptide. Appl. Microbiol. Biotechnol. 2016, 100, 5069–5077. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence a | aa b | MW c | z d | <H> e | <μH> f |
|---|---|---|---|---|---|---|
| Ctriporin | FLWGLIPGAISAVTSLIKK | 19 | 2013.5 | 3 | 0.805 | 0.452 |
| CM1 | FLWKLIPGAISAVTSLIKK | 19 | 2084.6 | 4 | 0.753 | 0.503 |
| CM2 | FLWKLIPKAISAVTSLIKK | 19 | 2155.7 | 5 | 0.701 | 0.547 |
| CM3 | FLWKLIPKAIKAVTSLIKK | 19 | 2196.8 | 6 | 0.651 | 0.592 |
| CM4 | FLWKLIPKAIKKVTSLIKK | 19 | 2253.9 | 7 | 0.583 | 0.614 |
| CM5 | FLWKLIPKAIKKVKSLIKK | 19 | 2282.0 | 8 | 0.517 | 0.615 |
| CM6 | FLWKLIPKAIKKVKKLIKK | 19 | 2323.1 | 9 | 0.467 | 0.664 |
| CM7 | FLWKLIKKAIKKVKKLIKK | 19 | 2353.1 | 10 | 0.377 | 0.704 |
| S. aureus ATCC29213 | S. aureus ATCC25923 | E. faecium ATCC29212 | E. coli ATCC25922 | E. coli ATCC35218 | P. aeruginosa ATCC27853 | K. pneumoniae ATCC700603 | A. baumannii ATCC19606 | ||
|---|---|---|---|---|---|---|---|---|---|
| MICs μg/mL (μM) | Hemolysis% at 256 μg/mL | ||||||||
| Ctriporin | 16 (8.0) | 32 (15.9) | 64 (31.8) | 64 (31.8) | >64 (31.8) | >64 (31.8) | >64 (31.8) | 16 (8.0) | 90.6 ± 11.8 |
| CM1 | 8 (3.8) | 16 (7.7) | 16 (7.7) | 8 (3.9) | 32 (15.4) | >64 (30.7) | 64 (30.7) | 8 (3.8) | 75.6 ± 10.1 |
| CM2 | 4 (1.9) | 8 (3.7) | 4 (1.9) | 4 (1.9) | 8 (3.7) | 16 (7.4) | 16 (7.4) | 4 (1.9) | 74.3 ± 9.2 |
| CM3 | 4 (1.8) | 4 (1.8) | 4 (1.8) | 4 (1.8) | 4 (1.8) | 8 (3.6) | 8 (3.6) | 4 (1.8) | 89.1 ± 12.1 |
| CM4 | 4 (1.8) | 8 (3.6) | 16 (7.1) | 8 (3.6) | 16 (7.1) | 16 (7.1) | 32 (14.2) | 4 (1.8) | 14.0 ± 4.7 |
| CM5 | 4 (1.8) | 4 (1.8) | 8 (3.5) | 8 (3.5) | 4 (1.8) | 4 (1.8) | 8 (3.5) | 2 (0.9) | 9.8 ± 0.6 |
| CM6 | 8 (3.4) | 8 (3.4) | 8 (3.4) | 8 (3.4) | 8 (3.4) | 4 (1.7) | 8 (3.4) | 4 (1.7) | 8.1 ± 1.9 |
| CM7 | 32 (13.6) | 32 (13.6) | 32 (13.6) | 32 (13.6) | 16 (6.8) | 8 (3.4) | 32 (6.8) | 8 (3.4) | 57.5 ± 9.8 |
| Strains | Resistance | Ctriporin | CM5 | CM6 |
|---|---|---|---|---|
| MICs μg/mL (μM) | ||||
| S. aureus 9124 | MRSA a | 8 (4.0) | 8 (3.5) | 8 (3.4) |
| S. aureus 891 | MRSA | 16 (8.0) | 4 (1.8) | 8 (3.4) |
| S. epidermidis 6943 | MRSE b | 8 (4.0) | 4 (1.8) | 4 (1.7) |
| S. epidermidis 9092 | MRSE | 8 (4.0) | 4 (1.8) | 4 (1.7) |
| S. capitis 3255 | MRSC c | 8 (4.0) | 2 (0.9) | 2 (0.86) |
| E. faecium 898 | MDR d | 16 (8.0) | 4 (1.8) | 4 (1.7) |
| E. coli 2678 | ESBL e | 64 (31.8) | 8 (3.5) | 8 (3.4) |
| E. coli 2687 | ESBL | >64 (31.8) | 8 (3.5) | 8 (3.4) |
| P. aeruginosa 9014 | CRE f/MDR | >64 (31.8) | 8 (3.5) | 8 (3.4) |
| K. pneumoniae 9126 | CRE | >64 (31.8) | 16 (7.0) | 16 (6.9) |
| A. baumannii 19295 | CRE/MDR | 32 (15.9) | 2 (0.9) | 2 (0.86) |
| A. baumannii 19462 | CRE/MDR | 16 (8.0) | 2 (0.9) | 2 (0.86) |
| A. baumannii 19618 | CRE/MDR | 32 (15.9) | 2 (0.9) | 2 (0.86) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Deng, H.; Ding, L.; Ye, X.; Sun, F.; Qin, C.; Chen, Z. Cationicity Enhancement on the Hydrophilic Face of Ctriporin Significantly Reduces Its Hemolytic Activity and Improves the Antimicrobial Activity against Antibiotic-Resistant ESKAPE Pathogens. Toxins 2024, 16, 156. https://doi.org/10.3390/toxins16030156
Luo X, Deng H, Ding L, Ye X, Sun F, Qin C, Chen Z. Cationicity Enhancement on the Hydrophilic Face of Ctriporin Significantly Reduces Its Hemolytic Activity and Improves the Antimicrobial Activity against Antibiotic-Resistant ESKAPE Pathogens. Toxins. 2024; 16(3):156. https://doi.org/10.3390/toxins16030156
Chicago/Turabian StyleLuo, Xudong, Huan Deng, Li Ding, Xiangdong Ye, Fang Sun, Chenhu Qin, and Zongyun Chen. 2024. "Cationicity Enhancement on the Hydrophilic Face of Ctriporin Significantly Reduces Its Hemolytic Activity and Improves the Antimicrobial Activity against Antibiotic-Resistant ESKAPE Pathogens" Toxins 16, no. 3: 156. https://doi.org/10.3390/toxins16030156