NetB, a Pore-Forming Toxin from Necrotic Enteritis Strains of Clostridium perfringens

{kind=link}
{kind=link}
Abstract
:1. Introduction

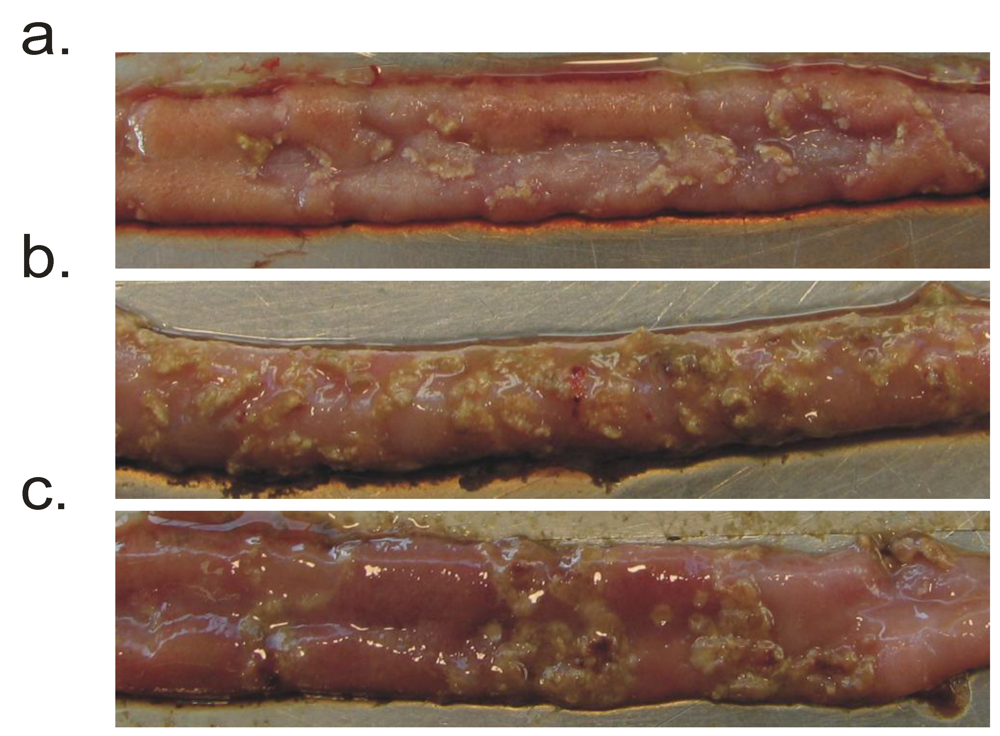
2. Identification and Analysis of NetB

3. Strain Distribution
4. Regulation of NetB Production and NetB Sequence Variation
5. NetB and the Development of a Necrotic Enteritis Vaccine
6. Conclusions
Acknowledgements
References
- Hatheway, C.L. Toxigenic clostridia. Clin. Microbiol. Rev. 1990, 3, 66–98. [Google Scholar]
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens: toxinotype and genotype. Trends Microbiol. 1999, 7, 104–110. [Google Scholar]
- Rood, J.I. Virulence genes of Clostridium perfringens. Annu. Rev. Microbiol. 1998, 52, 333–360. [Google Scholar]
- Smedley, J.G., III; Fisher, D.J.; Sayeed, S.; Chakrabarti, G.; McClane, B.A. The enteric toxins of Clostridium perfringens. Rev. Physiol. Biochem. Pharmacol. 2004, 152, 183–204. [Google Scholar] [PubMed]
- Songer, J.G. Clostridial enteric diseases of domestic animals. Clin. Microbiol. Rev. 1996, 9, 216–234. [Google Scholar]
- Brynestad, S.; Granum, P.E. Clostridium perfringens and foodborne infections. Int. J. Food Microbiol. 2002, 74, 195–202. [Google Scholar]
- Rood, J.I.; McClane, B.A. Clostridium perfringens: enterotoxaemic Diseases. In Molecular Medical Microbiology; Sussman, M., Ed.; Academic: London, UK, 2002; Volume 1, pp. 1117–1139. [Google Scholar]
- Rood, J.I. Clostridium perfringens and histotoxic disease. In The Prokaryotes: A Handbook on the Biology of Bacteria, 3rd; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2007; Volume 4, pp. 753–770. [Google Scholar]
- Van der Sluis, W. Clostridial enteritis—a syndrome. World Poultry 2000, 16, 56–57. [Google Scholar]
- Van der Sluis, W. Clostridial enteritis is an often underestimated problem. World Poultry 2000, 16, 42–43. [Google Scholar]
- Lovland, A.; Kaldhusdal, M. Severely impaired production performance in broiler flocks with high incidence of Clostridium perfringens-associated hepatitis. Avian Pathol. 2001, 30, 73–81. [Google Scholar]
- Kaldhusdal, M.; Hofshagen, M. Barley inclusion and avoparcin supplementation in broiler diets. 2. Clinical, pathological, and bacteriological findings in a mild form of necrotic enteritis. Poult. Sci. 1992, 71, 1145–1153. [Google Scholar] [PubMed]
- Helmboldt, C.F.; Bryant, E.S. The pathology of necrotic enteritis in domestic fowl. Avian Dis. 1971, 15, 775–780. [Google Scholar]
- Al-Sheikhly, F.; Truscott, R.B. The pathology of necrotic enteritis of chickens following infusion of broth cultures of Clostridium perfringens into the duodenum. Avian Dis. 1977, 21, 230–240. [Google Scholar]
- Al-Sheikhly, F.; Al-Saieg, A. Role of Coccidia in the occurrence of necrotic enteritis of chickens. Avian Dis. 1980, 24, 324–333. [Google Scholar]
- Gazdzinski, P.; Julian, R.J. Necrotic enteritis in turkeys. Avian Dis. 1992, 36, 792–798. [Google Scholar]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: an emerging threat for animal and public health. Avian Pathol. 2004, 33, 537–549. [Google Scholar]
- Broussard, C.T.; Hofacre, C.L.; Page, R.K.; Fletcher, O.J. Necrotic enteritis in cage-reared commercial layer pullets. Avian Dis. 1986, 30, 617–619. [Google Scholar]
- Awad, M.M.; Bryant, A.E.; Stevens, D.L.; Rood, J.I. Virulence studies on chromosomal alpha-toxin and theta-toxin mutants constructed by allelic exchange provide genetic evidence for the essential role of alpha-toxin in Clostridium perfringens-mediated gas gangrene. Mol. Microbiol. 1995, 15, 191–202. [Google Scholar]
- Al-Sheikhly, F.; Truscott, R.B. The pathology of necrotic enteritis of chickens following infusion of crude toxins of Clostridium perfringens into the duodenum. Avian Dis. 1977, 21, 241–255. [Google Scholar]
- Fukata, T.; Hadate, Y.; Baba, E.; Uemura, T.; Arakawa, A. Influence of Clostridium perfringens and its toxin in germ-free chickens. Res. Vet. Sci. 1988, 44, 68–70. [Google Scholar]
- Keyburn, A.L.; Sheedy, S.A.; Ford, M.E.; Williamson, M.M.; Awad, M.M.; Rood, J.I.; Moore, R.J. Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens. Infect. Immun. 2006, 74, 6496–6500. [Google Scholar]
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; Di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. 2008, 4, e26. [Google Scholar]
- Alouf, J.E. Pore-forming bacterial protein toxins: an overview. Curr. Top. Microbiol. Immunol. 2001, 257, 1–14. [Google Scholar]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar]
- Paton, J.C. The contribution of pneumolysin to the pathogenicity of Streptococcus pneumoniae. Trends Microbiol. 1996, 4, 103–106. [Google Scholar]
- Rubins, J.B.; Charboneau, D.; Fasching, C.; Berry, A.M.; Paton, J.C.; Alexander, J.E.; Andrew, P.W.; Mitchell, T.J.; Janoff, E.N. Distinct roles for pneumolysin's cytotoxic and complement activities in the pathogenesis of pneumococcal pneumonia. Am. J. Respir. Crit. Care Med. 1996, 153, 1339–1346. [Google Scholar]
- Ellemor, D.M.; Baird, R.N.; Awad, M.M.; Boyd, R.L.; Rood, J.I.; Emmins, J.J. Use of genetically manipulated strains of Clostridium perfringens reveals that both alpha-toxin and theta-toxin are required for vascular leukostasis to occur in experimental gas gangrene. Infect. Immun. 1999, 67, 4902–4907. [Google Scholar]
- Schmiel, D.H.; Miller, V.L. Bacterial phospholipases and pathogenesis. Microbes Infect. 1999, 1, 1103–1112. [Google Scholar]
- Titball, R.W. Membrane-damaging and cytotoxic phospholipases. In The Comprehensive Sourcebook of Bacterial Protein Toxins; Alouf, J.E., Freer, J.H., Eds.; Academic: London, UK, 1999; pp. 311–329. [Google Scholar]
- Kennedy, C.L.; Krejany, E.O.; Young, L.F.; O'Connor, J.R.; Awad, M.M.; Boyd, R.L.; Emmins, J.J.; Lyras, D.; Rood, J.I. The alpha-toxin of Clostridium septicum is essential for virulence. Mol. Microbiol. 2005, 57, 1357–1366. [Google Scholar]
- Prevost, G.; Mourey, L.; Colin, D.A.; Menestrina, G. Staphylococcal pore-forming toxins. Curr. Top. Microbiol. Immunol. 2001, 257, 53–83. [Google Scholar]
- Gouaux, E. alpha-Hemolysin from Staphylococcus aureus: an archetype of beta-barrel, channel-forming toxins. J. Struct. Biol. 1998, 121, 110–122. [Google Scholar]
- Tweten, R.K.; Christianson, K.K.; Iandolo, J.J. Transport and processing of staphylococcal alpha-toxin. J. Bacteriol. 1983, 156, 524–528. [Google Scholar]
- Gray, G.S.; Kehoe, M. Primary sequence of the alpha-toxin gene from Staphylococcus aureus wood 46. Infect. Immun. 1984, 46, 615–618. [Google Scholar]
- Shatursky, O.; Bayles, R.; Rogers, M.; Jost, B.H.; Songer, J.G.; Tweten, R.K. Clostridium perfringens beta-toxin forms potential-dependent, cation-selective channels in lipid bilayers. Infect. Immun. 2000, 68, 5546–5551. [Google Scholar]
- Sakurai, J.; Duncan, C.L. Some properties of beta-toxin produced by Clostridium perfringens type C. Infect. Immun. 1978, 21, 678–680. [Google Scholar]
- Vidal, J.E.; McClane, B.A.; Saputo, J.; Parker, J.; Uzal, F.A. Effects of Clostridium perfringens beta-toxin on the rabbit small intestine and colon. Infect. Immun. 2008, 76, 4396–4404. [Google Scholar]
- Katayama, S.; Dupuy, B.; Daube, G.; China, B.; Cole, S.T. Genome mapping of Clostridium perfringens strains with I-CeuI shows many virulence genes to be plasmid-borne. Mol. Gen. Genet. 1996, 251, 720–726. [Google Scholar]
- Hunter, S.E.; Brown, J.E.; Oyston, P.C.; Sakurai, J.; Titball, R.W. Molecular genetic analysis of beta-toxin of Clostridium perfringens reveals sequence homology with alpha-toxin, gamma-toxin, and leukocidin of Staphylococcus aureus. Infect. Immun. 1993, 61, 3958–3965. [Google Scholar]
- Steinthorsdottir, V.; Fridriksdottir, V.; Gunnarsson, E.; Andresson, O.S. Site-directed mutagenesis of Clostridium perfringens beta-toxin: expression of wild-type and mutant toxins in Bacillus subtilis. FEMS Microbiol. Lett. 1998, 158, 17–23. [Google Scholar]
- Nagahama, M.; Kihara, A.; Miyawaki, T.; Mukai, M.; Sakaguchi, Y.; Ochi, S.; Sakurai, J. Clostridium perfringens beta-toxin is sensitive to thiol-group modification but does not require a thiol group for lethal activity. Biochim. Biophys. Acta 1999, 1454, 97–105. [Google Scholar]
- Walker, B.; Bayley, H. Key residues for membrane binding, oligomerization, and pore forming activity of staphylococcal alpha-hemolysin identified by cysteine scanning mutagenesis and targeted chemical modification. J. Biol. Chem. 1995, 270, 23065–23071. [Google Scholar]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar]
- Nagahama, M.; Hayashi, S.; Morimitsu, S.; Sakurai, J. Biological activities and pore formation of Clostridium perfringens beta toxin in HL 60 cells. J. Biol. Chem. 2003, 278, 36934–36941. [Google Scholar]
- Manich, M.; Knapp, O.; Gibert, M.; Maier, E.; Jolivet-Reynaud, C.; Geny, B.; Benz, R.; Popoff, M.R. Clostridium perfringens delta toxin is sequence related to beta toxin, NetB, and Staphylococcus pore-forming toxins, but shows functional differences. PLoS One 2008, 3, e3764. [Google Scholar]
- Alouf, J.E.; Jolivet-Reynaud, C. Purification and characterization of Clostridium perfringens delta-toxin. Infect. Immun. 1981, 31, 536–546. [Google Scholar]
- Jolivet-Reynaud, C.; Cavaillon, J.M.; Alouf, J.E. Selective cytotoxicity of Clostridium perfringens delta toxin on rabbit leukocytes. Infect. Immun. 1982, 38, 860–864. [Google Scholar]
- Cavaillon, J.M.; Jolivet-Reynaud, C.; Fitting, C.; David, B.; Alouf, J.E. Ganglioside identification on human monocyte membrane with Clostridium perfringens delta-toxin. J. Leukoc. Biol. 1986, 40, 65–72. [Google Scholar]
- Jolivet-Reynaud, C.; Launay, J.M.; Alouf, J.E. Damaging effects of Clostridium perfringens delta toxin on blood platelets and their relevance to ganglioside GM2. Arch. Biochem. Biophys. 1988, 262, 59–66. [Google Scholar]
- Patel, A.H.; Nowlan, P.; Weavers, E.D.; Foster, T. Virulence of protein A-deficient and alpha-toxin-deficient mutants of Staphylococcus aureus isolated by allele replacement. Infect. Immun. 1987, 55, 3103–3110. [Google Scholar]
- Sayeed, S.; Uzal, F.A.; Fisher, D.J.; Saputo, J.; Vidal, J.E.; Chen, Y.; Gupta, P.; Rood, J.I.; McClane, B.A. Beta toxin is essential for the intestinal virulence of Clostridium perfringens type C disease isolate CN3685 in a rabbit ileal loop model. Mol. Microbiol. 2008, 67, 15–30. [Google Scholar]
- Bergeland, M.E. Pathogenesis and immunity of Clostridium perfringens type C enteritis in swine. J. Am. Vet. Med. Assoc. 1972, 160, 568–571. [Google Scholar]
- Fisher, D.J.; Fernandez-Miyakawa, M.E.; Sayeed, S.; Poon, R.; Adams, V.; Rood, J.I.; Uzal, F.A.; McClane, B.A. Dissecting the contributions of Clostridium perfringens type C toxins to lethality in the mouse intravenous injection model. Infect. Immun. 2006, 74, 5200–5210. [Google Scholar]
- Lepp, D.; Roxas, B.; Parreira, V.R.; Marri, P.R.; Rosey, E.L.; Gong, J.; Songer, J.G.; Vedantam, G.; Prescott, J.F. Identification of novel pathogenicity loci in Clostridium perfringens strains that cause avian necrotic enteritis. PLoS One 2010, 5, e10795. [Google Scholar]
- Engstrom, B.E.; Fermer, C.; Lindberg, A.; Saarinen, E.; Baverud, V.; Gunnarsson, A. Molecular typing of isolates of Clostridium perfringens from healthy and diseased poultry. Vet. Microbiol. 2003, 94, 225–235. [Google Scholar]
- Nauerby, B.; Pedersen, K.; Madsen, M. Analysis by pulsed-field gel electrophoresis of the genetic diversity among Clostridium perfringens isolates from chickens. Vet. Microbiol. 2003, 94, 257–266. [Google Scholar]
- Gholamiandekhordi, A.R.; Ducatelle, R.; Heyndrickx, M.; Haesebrouck, F.; Van Immerseel, F. Molecular and phenotypical characterization of Clostridium perfringens isolates from poultry flocks with different disease status. Vet. Microbiol. 2006, 113, 143–152. [Google Scholar]
- Chalmers, G.; Bruce, H.L.; Hunter, D.B.; Parreira, V.R.; Kulkarni, R.R.; Jiang, Y.F.; Prescott, J.F.; Boerlin, P. Multilocus sequence typing analysis of Clostridium perfringens isolates from necrotic enteritis outbreaks in broiler chicken populations. J. Clin. Microbiol. 2008, 46, 3957–3964. [Google Scholar]
- Chalmers, G.; Martin, S.W.; Hunter, D.B.; Prescott, J.F.; Weber, L.J.; Boerlin, P. Genetic diversity of Clostridium perfringens isolated from healthy broiler chickens at a commercial farm. Vet. Microbiol. 2008, 127, 116–127. [Google Scholar]
- Chalmers, G.; Martin, S.W.; Prescott, J.F.; Boerlin, P. Typing of Clostridium perfringens by multiple-locus variable number of tandem repeats analysis. Vet. Microbiol. 2008, 128, 126–135. [Google Scholar]
- Drigo, I.; Agnoletti, F.; Bacchin, C.; Bettini, F.; Cocchi, M.; Ferro, T.; Marcon, B.; Bano, L. Toxin genotyping of Clostridium perfringens field strains isolated from healthy and diseased chickens. Ital. J. Anim. Sci. 2008, 7, 397–400. [Google Scholar]
- Timbermont, L.; Lanckriet, A.; Gholamiandehkordi, A.R.; Pasmans, F.; Martel, A.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Origin of Clostridium perfringens isolates determines the ability to induce necrotic enteritis in broilers. Comp. Immunol. Microbiol. Infect. Dis. 2008, 32, 503–512. [Google Scholar]
- Johansson, A.; Aspan, A.; Kaldhusdal, M.; Engstrom, B.E. Genetic diversity and prevalence of netB in Clostridium perfringens isolated from a broiler flock affected by mild necrotic enteritis. Vet. Microbiol. 2009, 144, 87–92. [Google Scholar]
- Martin, T.G.; Smyth, J.A. Prevalence of netB among some clinical isolates of Clostridium perfringens from animals in the United States. Vet. Microbiol. 2009, 136, 202–205. [Google Scholar]
- Keyburn, A.L.; Yan, X.X.; Bannam, T.L.; Van Immerseel, F.; Rood, J.I.; Moore, R.J. Association between avian necrotic enteritis and Clostridium perfringens strains expressing NetB toxin. Vet. Res. 2010, 41, 21. [Google Scholar]
- Smyth, J.A.; Martin, T.G. Disease producing capability of netB positive isolates of C. perfringens recovered from normal chickens and a cow, and netB positive and negative isolates from chickens with necrotic enteritis. Vet. Microbiol. 2010. [Google Scholar]
- Abildgaard, L.; Sondergaard, T.E.; Engberg, R.M.; Schramm, A.; Hojberg, O. In vitro production of necrotic enteritis toxin B, NetB, by netB-positive and netB-negative Clostridium perfringens originating from healthy and diseased broiler chickens. Vet. Microbiol. 2010, 144, 231–235. [Google Scholar]
- Cooper, K.K.; Songer, J.G. Virulence of Clostridium perfringens in an experimental model of poultry necrotic enteritis. Vet. Microbiol. 2010, 142, 323–328. [Google Scholar]
- Nowell, V.J.; Poppe, C.; Parreira, V.R.; Jiang, Y.F.; Reid-Smith, R.; Prescott, J.F. Clostridium perfringens in retail chicken. Anaerobe 2010, 16, 314–315. [Google Scholar]
- Cheung, J.K.; Keyburn, A.L.; Carter, G.P.; Lanckriet, A.L.; Van Immerseel, F.; Moore, R.J.; Rood, J.I. The VirSR two-component signal transduction system regulates NetB toxin production in Clostridium perfringens. Infect. Immun. 2010, 78, 3064–3072. [Google Scholar]
- Ohtani, K.; Hirakawa, H.; Tashiro, K.; Yoshizawa, S.; Kuhara, S.; Shimizu, T. Identification of a two-component VirR/VirS regulon in Clostridium perfringens. Anaerobe 2010, 16, 258–264. [Google Scholar]
- Ohtani, K.; Yuan, Y.; Hassan, S.; Wang, R.; Wang, Y.; Shimizu, T. Virulence gene regulation by the agr system in Clostridium perfringens. J. Bacteriol. 2009, 191, 3919–3927. [Google Scholar]
- McDevitt, R.M.; Brooker, J.D.; Acamovic, T.; Sparks, N.H.C. Necrotic enteritis; a continuing challenge for the poultry industry. Worlds Poult. Sci. J. 2006, 62, 221–247. [Google Scholar]
- Lawrence, G.W.; Lehmann, D.; Anian, G.; Coakley, C.A.; Saleu, G.; Barker, M.J.; Davis, M.W. Impact of active immunisation against enteritis necroticans in Papua New Guinea. Lancet 1990, 336, 1165–1167. [Google Scholar]
- Uzal, F.A.; Kelly, W.R. Protection of goats against experimental enterotoxaemia by vaccination with Clostridium perfringens type D epsilon toxoid. Vet. Rec. 1998, 142, 722–725. [Google Scholar]
- Springer, S.; Selbitz, H.J. The control of necrotic enteritis in sucking piglets by means of a Clostridium perfringens toxoid vaccine. FEMS Immunol. Med. Microbiol. 1999, 24, 333–336. [Google Scholar]
- Heier, B.T.; Lovland, A.; Soleim, K.B.; Kaldhusdal, M.; Jarp, J. A field study of naturally occurring specific antibodies against Clostridium perfringens alpha toxin in Norwegian broiler flocks. Avian Dis. 2001, 45, 724–732. [Google Scholar]
- Lovland, A.; Kaldhusdal, M.; Redhead, K.; Skjerve, E.; Lillehaug, A. Maternal vaccination against subclinical necrotic enteritis in broilers. Avian Pathol. 2004, 33, 83–92. [Google Scholar]
- Kulkarni, R.R.; Parreira, V.R.; Sharif, S.; Prescott, J.F. Clostridium perfringens antigens recognized by broiler chickens immune to necrotic enteritis. Clin. Vaccine Immunol. 2006, 13, 1358–1362. [Google Scholar]
- Kulkarni, R.R.; Parreira, V.R.; Sharif, S.; Prescott, J.F. Immunization of broiler chickens against Clostridium perfringens-induced necrotic enteritis. Clin. Vaccine Immunol. 2007, 14, 1070–1077. [Google Scholar]
- Zekarias, B.; Mo, H.; Curtiss, R., III. Recombinant attenuated Salmonella enterica serovar typhimurium expressing the carboxy-terminal domain of alpha toxin from Clostridium perfringens induces protective responses against necrotic enteritis in chickens. Clin. Vaccine Immunol. 2008, 15, 805–816. [Google Scholar]
- Wardenburg, J.B.; Schneewind, O. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 2008, 205, 287–294. [Google Scholar]
- Ragle, B.E.; Wardenburg, J.B. Anti-alpha-hemolysin monoclonal antibodies mediate protection against Staphylococcus aureus pneumonia. Infect. Immun. 2009, 77, 2712–2718. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Keyburn, A.L.; Bannam, T.L.; Moore, R.J.; Rood, J.I. NetB, a Pore-Forming Toxin from Necrotic Enteritis Strains of Clostridium perfringens. Toxins 2010, 2, 1913-1927. https://doi.org/10.3390/toxins2071913
Keyburn AL, Bannam TL, Moore RJ, Rood JI. NetB, a Pore-Forming Toxin from Necrotic Enteritis Strains of Clostridium perfringens. Toxins. 2010; 2(7):1913-1927. https://doi.org/10.3390/toxins2071913
Chicago/Turabian StyleKeyburn, Anthony L., Trudi L. Bannam, Robert J. Moore, and Julian I. Rood. 2010. "NetB, a Pore-Forming Toxin from Necrotic Enteritis Strains of Clostridium perfringens" Toxins 2, no. 7: 1913-1927. https://doi.org/10.3390/toxins2071913
APA StyleKeyburn, A. L., Bannam, T. L., Moore, R. J., & Rood, J. I. (2010). NetB, a Pore-Forming Toxin from Necrotic Enteritis Strains of Clostridium perfringens. Toxins, 2(7), 1913-1927. https://doi.org/10.3390/toxins2071913