Expression Analysis of Stress-Related Genes in Kernels of Different Maize (Zea mays L.) Inbred Lines with Different Resistance to Aflatoxin Contamination


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Gene Expression Analysis Using qPCR

{kind=link}
{kind=link}
{kind=link}
| Gene ID | Putative Annotation | B73 | Lo1016 | Mo17 | Mp313E | Tex6 | Lo964 | A638 | B73 * | Tex6 * |
|---|---|---|---|---|---|---|---|---|---|---|
| TC273692 | heat shock protein 21 | -- | -- | - | + | ++ | + | ++ | ×× | ++ |
| TC248621 | early drought induced protein | -- | -- | -- | ++ | ++ | ++ | + | + | + |
| TC247852 | abscisic acid inducible gene | -- | -- | -- | ++ | ++ | + | ++ | + | -- |
| TC260723 | putative salt-inducible protein kinase | - | - | - | + | + | + | + | ×× | - |
| TC249150 | proline-rich protein family-like | -- | -- | - | + | ++ | + | ++ | + | - |
| TC261320 | MAP kinase phosphatase | - | -- | - | + | + | + | ++ | ++ | - |
| TC248521 | lipid transfer protein | ++ | -- | -- | ++ | ++ | ++ | ++ | -- | + |
| TC259179 | γ-thionin | - | -- | + | ++ | ++ | ++ | + | ×× | + |
| TC270625 | glutathione reductase | - | -- | ×× | + | ++ | + | + | - | ×× |
| TC248890 | putative glutathione peroxidase | ++ | -- | -- | ++ | ++ | ++ | ++ | - | - |
| TC252272 | multi resistance protein | + | -- | -- | ++ | ++ | + | + | ×× | -- |
| TC260617 | putative MAP kinase | - | -- | ×× | + | + | + | + | ×× | ++ |
| TC261534 | putative hydroxyproline-rich glycoprotein DZ-HRGP | ×× | -- | + | ++ | + | + | ++ | + | ++ |
| TC261606 | leucine-rich repeat transmembrane protein kinase | ×× | -- | - | + | + | + | + | ++ | + |
| TC248912 | dehydrin DHN1 | + | ++ | ++ | ++ | -- | ++ | ×× | + | ++ |
| TC258769 | LEA14-A | ++ | ++ | ++ | ++ | -- | ++ | -- | ++ | ++ |
| TC269764 | glyoxalase I | - | -- | - | - | ++ | + | ++ | + | - |
| TC263499 | diamide resistance gene | - | -- | - | ×× | ++ | + | + | + | ++ |
| TC248251 | putative stress-related protein | -- | -- | -- | ++ | - | ++ | ++ | ++ | + |
| TC271560 | heat shock protein hsp22 precursor | - | -- | - | - | ++ | - | ++ | ++ | ++ |
| TC273584 | oxidation protection protein | -- | -- | - | ×× | + | + | ++ | + | ++ |
| TC248631 | unknown (myoD protein inhibitor) | - | -- | - | ×× | + | ×× | ++ | - | ++ |
| TC271775 | mitogen activated protein kinase | - | - | - | ×× | + | ×× | + | - | ++ |
| TC272055 | putative HSPC058 | - | - | - | - | + | + | ++ | ×× | - |
| TC250756 | polyphenol oxidase | - | - | - | -- | ++ | ++ | ++ | + | ++ |
| TC249851 | multidrug-resistance associated protein | - | -- | - | ×× | + | + | + | + | ++ |
| TC248721 | N/A | -- | -- | -- | ++ | - | - | ++ | ++ | + |
| TC260636 | leucine-rich repeat resistance protein-like protein | -- | - | - | + | + | - | + | - | + |
| AZM4-134720 | similar to water stress inducible protein | -- | -- | -- | ×× | ++ | - | ++ | -- | + |
| TC263714 | major facilitator superfamily antiporter | - | - | -- | + | ++ | + | ×× | -- | + |
| TC262308 | putative glycolate oxidase | -- | -- | - | ++ | + | - | ++ | - | + |
| TC259689 | cysteine protease | -- | -- | - | ++ | ++ | - | ++ | + | -- |
| TC261493 | thionin like protein | - | -- | -- | ×× | + | ++ | ++ | + | ×× |
| TC251520 | alpha globulin | - | -- | - | ×× | - | + | ++ | + | - |
| TC259802 | putative stress-induced protein | - | -- | - | + | + | + | ×× | - | -- |
| TC258326 | L-ascorbate peroxidase | - | -- | - | ×× | + | ++ | ++ | ++ | -- |
| TC271380 | probable trypsin inhibitor | -- | -- | -- | ++ | ++ | -- | ++ | -- | ×× |
| TC268744 | putative hydroxyproline-rich glycoprotein 1 | -- | -- | -- | ++ | ++ | ++ | -- | -- | + |
| TC250985 | unknown protein | -- | - | - | - | ×× | ++ | ++ | -- | + |
| TC269707 | r40g2 protein | -- | -- | - | + | ++ | - | + | + | -- |
| TC272484 | putative UVB-resistance protein | -- | -- | -- | - | ++ | + | ++ | - | + |
| TC261400 | receptor protein kinase PERK1-like protein | -- | -- | -- | ++ | ++ | -- | ++ | -- | ++ |
| TC251180 | hybrid proline-rich protein | ×× | ++ | ++ | -- | ++ | ++ | -- | - | + |
| TC258497 | metallothionein-like protein | - | -- | + | ×× | ++ | ×× | + | - | - |
| TC270445 | γ-zeathionin 1 | - | -- | ++ | -- | ++ | ++ | ++ | + | + |
| TC248731 | chitinase | -- | - | ×× | -- | ++ | ++ | ++ | + | + |
| CF630432 | bet v I allergen | - | - | ×× | + | ×× | + | + | - | + |
| TC269763 | subtilisin/chymotrypsin inhibitor | -- | + | ++ | ×× | -- | ++ | + | - | - |
| TC272650 | putative stress-inducible membrane pore protein | -- | -- | ×× | ++ | + | - | ++ | ×× | + |
| TC262243 | expressed protein | - | + | ++ | -- | ++ | + | - | -- | + |
| TC260600 | peroxiredoxin | -- | - | ++ | -- | ++ | ++ | ×× | ×× | - |
| TC248296 | nonspecific lipid-transfer protein precursor | -- | - | ++ | -- | ++ | ++ | ++ | + | ×× |
| TC271062 | NAM-related protein 1 | -- | -- | + | - | - | ++ | ++ | - | ++ |
| TC260324 | putative xylanase inhibitor protein | -- | ++ | ++ | -- | - | ++ | ++ | + | + |
| TC251880 | metallothionein | ×× | + | -- | ++ | ++ | ++ | - | + | -- |
| TC270149 | globulin-1S | -- | -- | ×× | ×× | + | ++ | ++ | + | - |
| TC253617 | putative serine/threonine-specific protein kinase | -- | - | + | + | - | + | ++ | -- | ++ |
| TC253449 | late embryogenesis abundant protein | -- | ++ | ++ | ++ | -- | ++ | ++ | + | ++ |
| TC261509 | putative aldose reductase | - | - | + | -- | -- | ++ | ++ | ++ | ++ |
| TC273536 | heat shock protein | + | - | - | -- | ++ | ++ | ×× | ++ | + |
| CF000577 | proline rich protein | -- | ×× | ++ | -- | -- | ++ | ++ | ++ | ++ |
| TC259915 | hageman factor inhibitor | + | -- | ×× | ++ | -- | + | + | + | -- |
| TC270070 | putative universal stress protein | ×× | -- | ×× | ×× | ×× | + | ++ | ++ | -- |
| TC258155 | putative glutathione S-transferase | -- | + | ++ | + | -- | ++ | ++ | - | ++ |
| TC270514 | hypothetical protein(aluminum-induced protein-like) | ++ | - | -- | + | - | - | ++ | - | + |
| TC248921 | putative peroxisome type ascorbate peroxidase | -- | + | ×× | ++ | + | ×× | ++ | - | ++ |
| TC249614 | Superoxide dismutase [Mn] | - | ×× | - | + | + | + | - | -- | ++ |
| TC258876 | hydroxyproline-rich glycoprotein precursor | ×× | - | + | + | -- | + | + | - | ++ |
| TC270339 | superoxide dismutase 2 | ++ | -- | ++ | ++ | -- | + | ++ | - | ++ |
| AI372267 | peroxidase | ++ | + | -- | - | ++ | -- | -- | ++ | - |
| TC259921 | antimicrobial peptide | ×× | - | + | -- | ++ | ++ | + | + | - |
| TC270782 | putative leucine-rich repeat transmembrane protein kinase | + | -- | -- | ++ | ++ | -- | ++ | -- | ×× |
| TC271639 | putative cytochrome P450 | ×× | ++ | -- | ++ | ++ | -- | ++ | -- | ++ |
| TC259396 | catalase | -- | - | + | ++ | ×× | + | ×× | -- | ++ |
| TC249070 | Superoxide dismutase [Cu-Zn] | + | - | -- | ++ | + | + | - | + | - |
| TC271423 | glutathione S-transferase | -- | ×× | -- | ++ | ++ | ++ | - | + | + |
| TC270868 | globulin 2 | -- | -- | ×× | - | + | ++ | ++ | ×× | -- |
| TC259180 | γ-zeathionin 2 | - | -- | + | + | ++ | ++ | - | - | -- |
| TC268733 | s-adenosylmethionine synthetase | - | - | ×× | + | ++ | ×× | - | -- | -- |
| TC270842 | cold shock protein | + | -- | - | + | ++ | ×× | ×× | + | ×× |
| TC260707 | salt-induced AAA-Type ATPase | -- | - | ×× | + | ×× | + | ++ | ++ | + |
| TC250578 | putative beta-1,3-glucanase | + | -- | -- | ++ | + | -- | + | -- | ++ |
| TC268849 | heat shock protein | ++ | -- | -- | -- | -- | - | ++ | ++ | ++ |
| AI372246 | lipoxygenase | + | -- | -- | -- | + | ++ | ++ | - | ++ |
| TC251457 | putative proline-rich protein | -- | -- | ++ | -- | -- | - | ++ | + | ++ |
| TC260910 | putative CC-NBS-LRR resistance protein MLA13 | -- | ++ | -- | ×× | ×× | ++ | ++ | ++ | ++ |
| TC264819 | putative antifungal zeamatin-like protein | -- | -- | ++ | - | - | -- | ++ | ++ | ++ |
| TC261221 | chalcone synthase | -- | -- | ×× | ×× | -- | + | ++ | -- | ++ |
| TC249478 | ascorbate peroxidase | ×× | -- | -- | ×× | ++ | ×× | ++ | ++ | ×× |
| TC259722 | cysteine protease component of protease-inhibitor complex | ++ | -- | ++ | -- | ++ | ++ | - | -- | ++ |
| TC202729 | ribosomal inactivating protein | ×× | -- | -- | + | ++ | - | ++ | -- | ×× |
| TC263586 | antifungal zeamatin-like protein | + | -- | -- | ++ | ++ | - | ++ | ++ | -- |
| AZM4-123774 | phenylalanine ammonia-lyase | + | - | + | - | ++ | - | + | + | -- |
| TC273961 | flavanone 3-hydroxylase | -- | -- | ++ | -- | + | - | ++ | -- | ++ |
3. Results
3.1. Aflatoxin Contamination in Different Maize Lines
| Genotypes | Aflatoxin (ppb) a | Significance | Reference b | |
|---|---|---|---|---|
| GTP2 | 924 ± 455 | a | ||
| B73 | 737 ± 45 | a | 1557 | 271 |
| Lo1016 | 663 ± 136 | ab | ||
| A638 | 560 ± 192 | ab | ||
| Mo17 | 507 ± 108 | ab | 950 | 152 |
| GTP27 | 494 ± 134 | ab | ||
| Mp313E | 434 ± 71 | b | 34 | |
| Lo964 | 332 ± 125 | bc | ||
| Tex6 | 291 ± 136 | c | 586 | 39 |
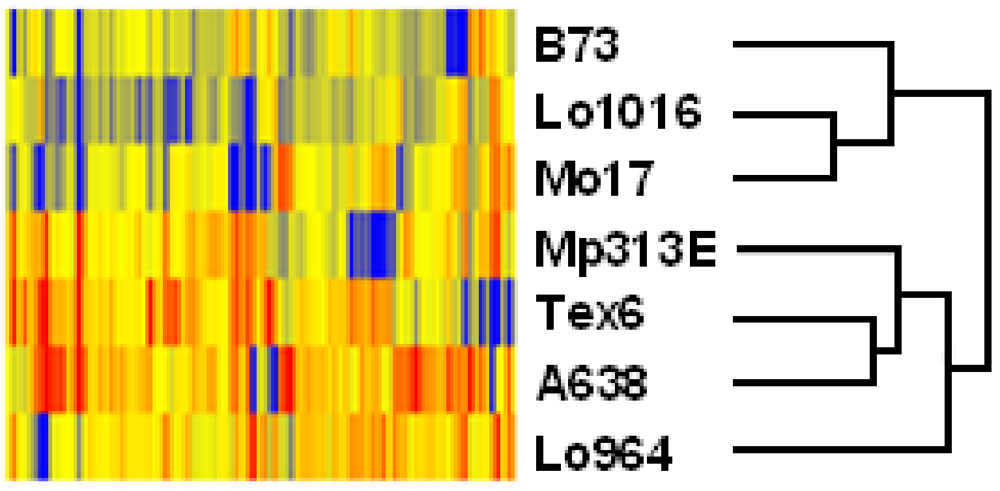
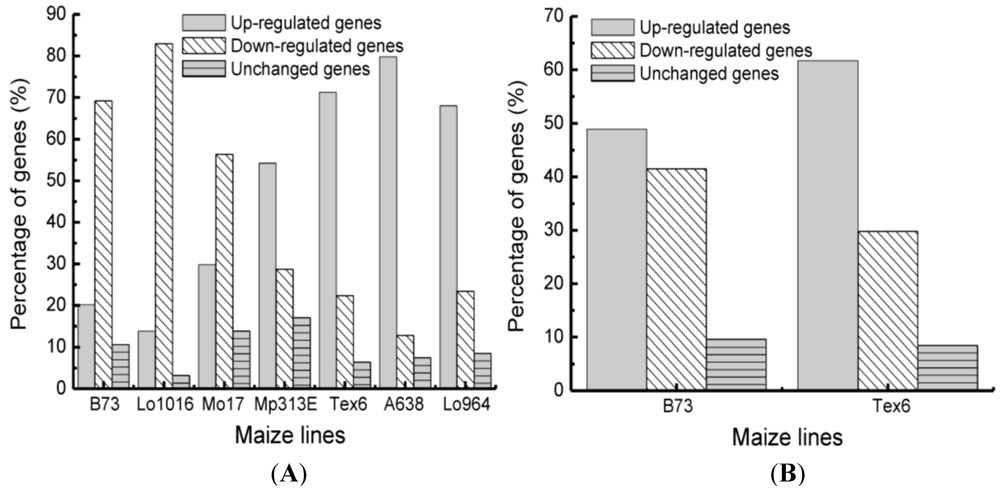
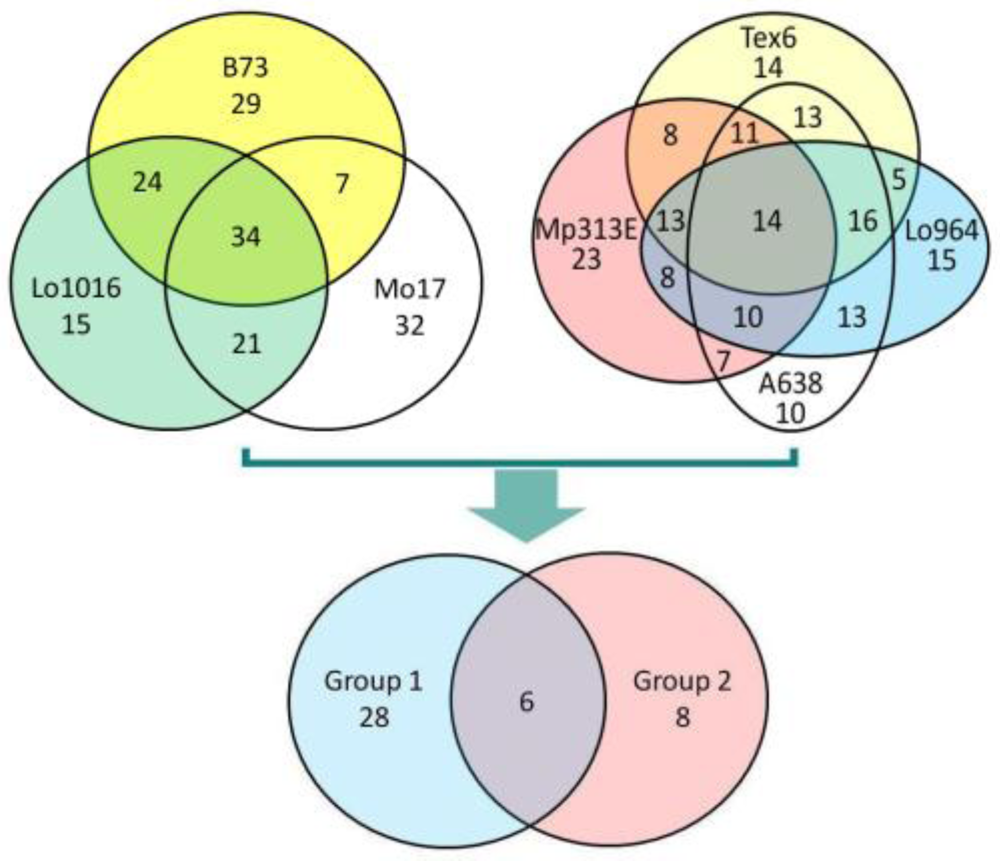
3.2. Differential Expression of Stress-Related Genes in Different Maize Lines



4. Discussion
Acknowledgement
References
- Luo, M.; Liu, J.; Lee, R.D.; Scully, B.T.; Guo, B.Z. Monitoring the expression of maize genes in developing kernels under drought stress using oligo-microarray. J. Integr. Plant Biol. 2010, 52, 1059–1074. [Google Scholar]
- Guo, B.Z.; Krakowsky, M.D.; Ni, X.; Scully, B.T.; Lee, R.D.; Coy, A.E.; Widstrom, N.W. Registration of Maize Inbred Line GT603. J. Plant Reg. 2011, 5, 211–214. [Google Scholar]
- Hamblin, A.M.; White, D.G. Inheritance of resistance to Aspergillus ear rot and aflatoxin production of corn from Tex6. Phytopathology 2000, 90, 292–296. [Google Scholar]
- Williams, W.P. Breeding for resistance to aflatoxin accumulation in maize. Mycotoxin Res. 2006, 22, 27–32. [Google Scholar]
- Cleveland, T.E.; Yu, J.; Bhatnagar, D.; Chen, Z.Y.; Brown, R.L.; Chang, P.K.; Cary, J.W. Progress in elucidating the molecular basis of the host plant—Aspergillus flavus interaction, a basis for devising strategies to reduce aflatoxin contamination in crops. J. Toxicol. Toxin Rev. 2004, 23, 345–380. [Google Scholar]
- Paul, C.; Naidoo, G.; Forbes, A.; Mikkilineni, V.; White, D.; Rocheford, T. Quantitative trait loci for low aflatoxin production in two related maize populations. Theor. Appl. Genet. 2003, 107, 263–270. [Google Scholar]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of a maize kernel stress-related protein and its effect on aflatoxin accumulation. Phytopathology 2004, 94, 938–945. [Google Scholar]
- Evers, D.; Ghislain, M.; Hausman, J.F.; Dommes, J. Differential gene expression in two potato lines differing in their resistance to Phytophthora infestans. J. Plant Physiol. 2003, 160, 709–712. [Google Scholar]
- Chen, Z.Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of unique or elevated levels of kernel proteins in aflatoxin-resistant maize genotypes through proteome analysis. Phytopathology 2002, 92, 1084–1094. [Google Scholar]
- Xu, D.; Duan, X.; Wang, B.; Hong, B.; Ho, T.H.D.; Wu, R. Expression of a late embryogenesis abundant protein gene HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol. 1996, 110, 249–257. [Google Scholar]
- Warburton, M.; Williams, P.; Hawkins, L.; Bridges, S.; Gresham, C.; Harper, J.; Ozkan, S.; Mylroie, E.; Shan, X. A public platform for the verification of the phenotypic effect of candidate genes for resistance to aflatoxin accumulation and A. flavus resistance in maize. Toxins 2011, 3. in press. [Google Scholar]
- Abbas, H.K.; Shier, W.T.; Cartwright, R.D. Effect of temperature rainfall and planting date on aflatoxin and fumonisin contamination in commercial Bt and non-Bt corn hybrids in Arkansas. Phytoprotection 2007, 88, 41–50. [Google Scholar]
- Luo, M.; Liu, J.; Lee, R.D.; Guo, B.Z. Characterization of gene expression profiles in developing kernels of maize (Zea mays L.) inbred Tex6. Plant Breed. 2008, 127, 569–578. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protocols 2008, 3, 1101–1108. [Google Scholar]
- Sanguineti, M.C.; Giuliani, M.M.; Govi, G.; Tuberosa, R.; Landi, P. Root and shoot traits of maize inbred lines grown in the field and in hydroponic culture and their relationship with root lodging. Maydica 1998, 43, 211–216. [Google Scholar]
- Carson, M.L.; Hooker, A.L. Inheritance of resistance to stalk rot of corn caused by Colletotricchum graminicola. Phytopathology 1981, 71, 1190–1196. [Google Scholar]
- Franco, O.; Murad, A.M.; Leite, J.R.; Mendes, P.A.M.; Prates, M.V.; Bloch, C., Jr. Identification of a cowpea γ-thionin with bactericidal activity. FEBS Lett. 2006, 273, 3489–3497. [Google Scholar]
- Guo, B.Z.; Chen, Z.-Y.; Brown, R.L.; Lax, A.R.; Cleveland, T.E.; Russin, J.S.; Mehta, A.D.; Selitrennikoff, C.P.; Widstrom, N.W. Germination induces accumulation of specific proteins and antifungal activities in corn kernels. Phytopathology 1997, 87, 1174–1178. [Google Scholar]
- Guo, B.Z.; Cleveland, T.E.; Brown, R.L.; Widstrom, N.W.; Lynch, R.E.; Russin, J.S. Distribution of antifungal proteins in maize kernel tissues using immunochemistry. J. Food Protect. 1999, 62, 295–299. [Google Scholar]
- Halterman, D.A.; Wei, F.; Wise, R.P. Powdery mildew-induced Mla mRNAs are alternatively spliced and contain multiple upstream open reading frames. Plant Physiol. 2003, 131, 558–567. [Google Scholar]
- Corbin, D.R.; Sauer, N.; Lamb, C.J. Differential regulation of a hydroxyproline-rich glycoprotein gene family in wounded and infected plants. Mol. Cell Biol. 1987, 7, 4337–4344. [Google Scholar]
- Sommer-Kundsen, J.; Bacic, A.; Clarke, A.E. Hydroxyproline-rich plant glycoprotein. Phytochemistry 1998, 47, 483–497. [Google Scholar]
- Briviba, K.; Kissner, R.; Koppenol, W.H.; Sies, H. Kinetic study of the reaction of glutathione peroxidase with peroxynitrite. Chem. Res. Toxicol. 1998, 11, 1398–1401. [Google Scholar]
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 2000, 97, 2940–2945. [Google Scholar]
- Mithöfer, A.; Schulze, B.; Boland, W. Biotic and heavy metal stress response in plants: Evidence for common signals. FEBS Lett. 2004, 566, 1–5. [Google Scholar]
- Teige, M.; Scheikl, E.; Eulgem, T.; Doczi, R.; Ichimura, K.; Shinozaki, K.; Dangl, J.L.; Hirt, H. The MKK2 pathway mediates cold and salt stress signaling in Arabidopsis. Mol. Cell 2004, 5, 141–152. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jiang, T.; Zhou, B.; Luo, M.; Abbas, H.K.; Kemerait, R.; Lee, R.D.; Scully, B.T.; Guo, B. Expression Analysis of Stress-Related Genes in Kernels of Different Maize (Zea mays L.) Inbred Lines with Different Resistance to Aflatoxin Contamination. Toxins 2011, 3, 538-550. https://doi.org/10.3390/toxins3060538
Jiang T, Zhou B, Luo M, Abbas HK, Kemerait R, Lee RD, Scully BT, Guo B. Expression Analysis of Stress-Related Genes in Kernels of Different Maize (Zea mays L.) Inbred Lines with Different Resistance to Aflatoxin Contamination. Toxins. 2011; 3(6):538-550. https://doi.org/10.3390/toxins3060538
Chicago/Turabian StyleJiang, Tingbo, Boru Zhou, Meng Luo, Hamed K. Abbas, Robert Kemerait, Robert Dewey Lee, Brian T. Scully, and Baozhu Guo. 2011. "Expression Analysis of Stress-Related Genes in Kernels of Different Maize (Zea mays L.) Inbred Lines with Different Resistance to Aflatoxin Contamination" Toxins 3, no. 6: 538-550. https://doi.org/10.3390/toxins3060538
APA StyleJiang, T., Zhou, B., Luo, M., Abbas, H. K., Kemerait, R., Lee, R. D., Scully, B. T., & Guo, B. (2011). Expression Analysis of Stress-Related Genes in Kernels of Different Maize (Zea mays L.) Inbred Lines with Different Resistance to Aflatoxin Contamination. Toxins, 3(6), 538-550. https://doi.org/10.3390/toxins3060538