Enzyme-Linked Immunosorbent-Assay for Deoxynivalenol (DON)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Immunization
2.3. Specificity
2.4. Indirect Competitive ELISA Assay
2.5. Optimized Working Concentration
2.6. The Effect of Methanol on Detecting DON by ELISA
2.7. Preparation of Samples
2.8. Standard Curve
2.9. Recovery Test
2.10. Calculation of the Concentration of DON in Sample
- C: concentration of DON in the immunoplate well;
- V: volume of sample (mL);
- M: weight of sample (g);
- D: dilution ratio.
3. Results and Analysis
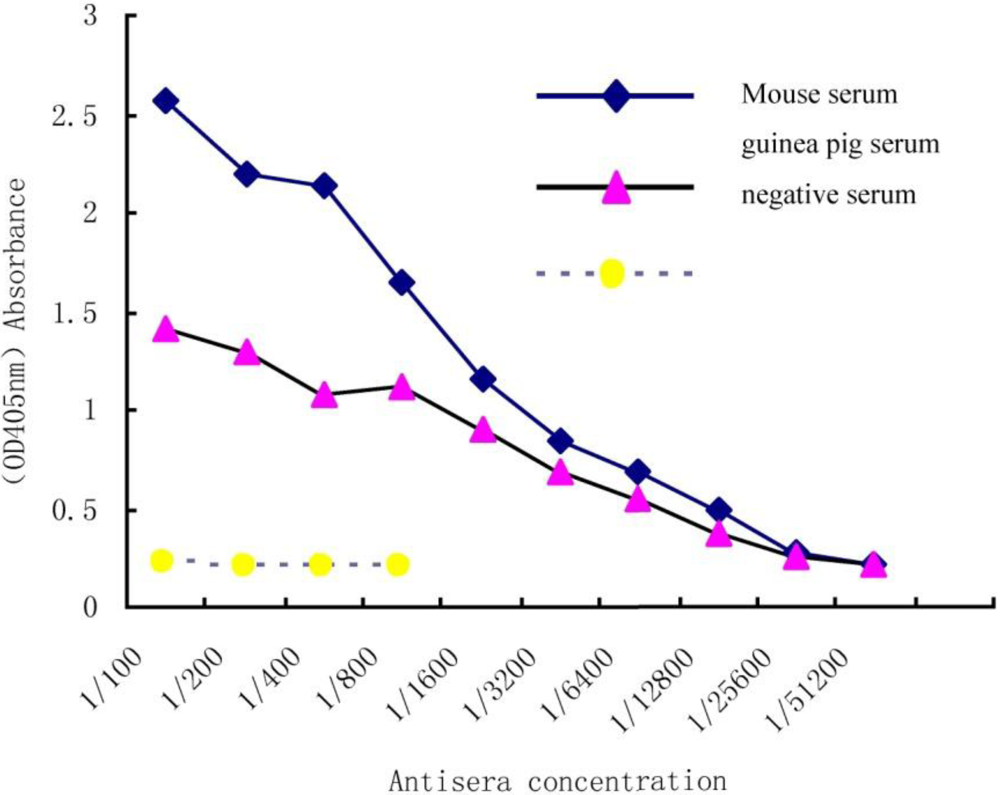
3.1. Titration of Antiserum

3.2. Optimized Working Concentration

{kind=link}
{kind=link}
{kind=link}
| Coated Antigen Concentration | Antisera Concentration | |||||
|---|---|---|---|---|---|---|
| 1/400 | 1/800 | 1/1600 | 1/3200 | 1/6400 | ||
| 1/500 | 2.620 | 2.447 | 2.269 | 2.040 | 1.685 | |
| 1/500 | 2.590 | 2.421 | 2.231 | 2.006 | 1.654 | |
| 1/800 | 1.309 | 1.102 | 1.001 | 0.773 | 0.579 | |
| 1/800 | 1.297 | 1.000 | 0.934 | 0.737 | 0.503 | |
| 1/1000 | 1.187 | 1.093 | 0.964 | 0.770 | 0.513 | |
| 1/1000 | 1.382 | 1.041 | 0.926 | 0.734 | 0.539 | |
| 1/1500 | 1.448 | 1.222 | 1.059 | 0.884 | 0.629 | |
| 1/1500 | 1.536 | 1.239 | 1.074 | 0.808 | 0.682 | |
| 1/2000 | 1.508 | 1.210 | 1.124 | 0.900 | 0.648 | |
| 1/2000 | 1.520 | 1.262 | 1.100 | 0.940 | 0.673 | |
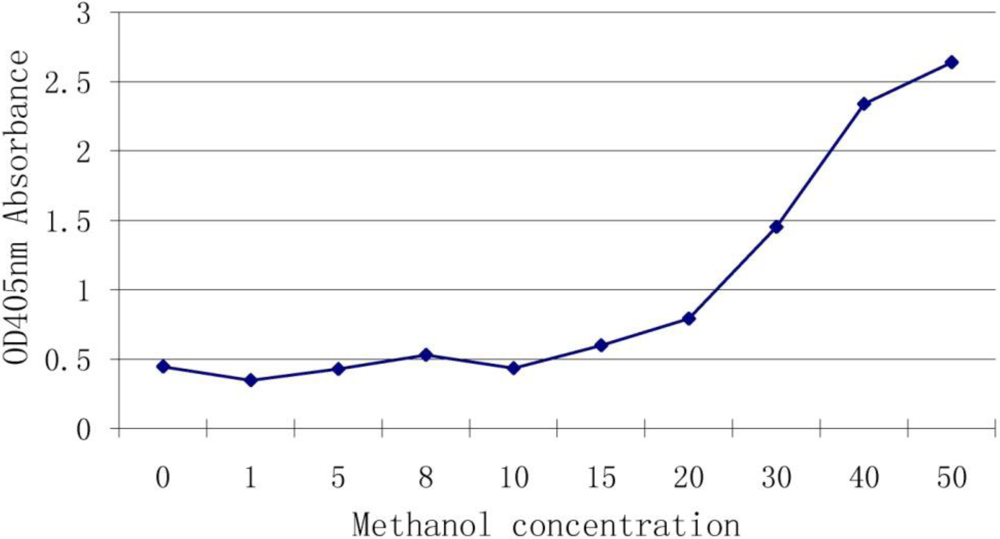
3.3. Effect of Methanol Concentration on the Assay

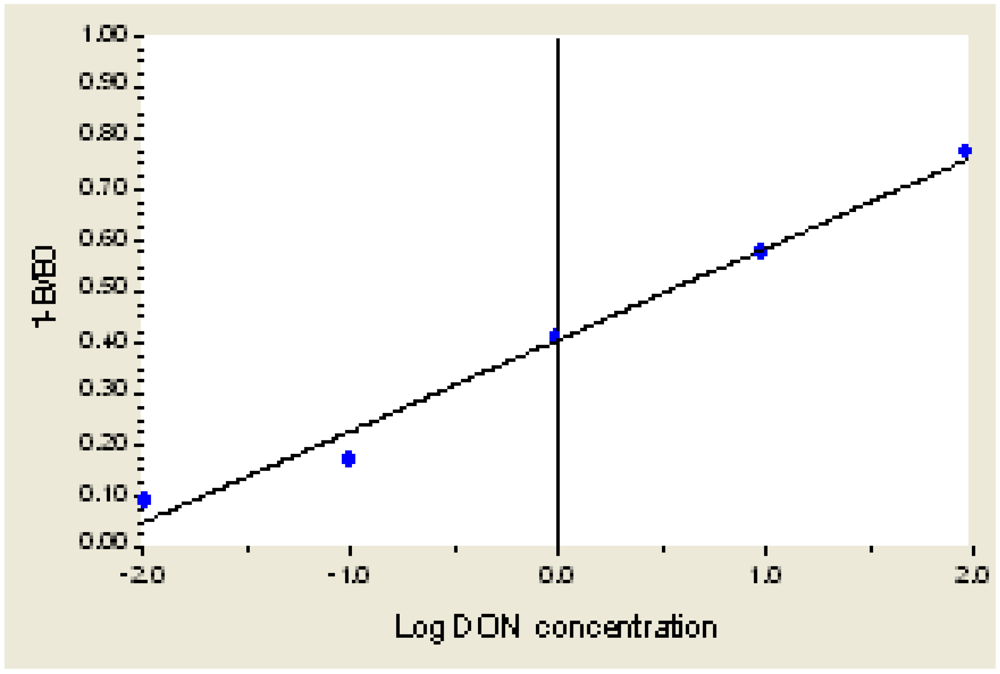
3.4. Standard Curve

3.5. Recovery Test
| Added DON (μg/g) | DON Detected (μg/g) | Recovery |
|---|---|---|
| 0.5 | 0.43 ± 0.02 | 86% |
| 10 | 9.3 ± 1.1 | 93% |
| 200 | 164 ± 35 | 82% |
3.6. Specificity
3.7. DON Detection in Sample
| Wheat Variety | FHB Resistance | DON (µg/g) |
|---|---|---|
| Sumai3 | Resistant | 3.98 |
| Wang shuibai | Resistant | 2.5 |
| Yangmai158 | Middle | 39.8 |
| Ningmai6 | Susceptible | 63.1 |
4. Discussion
4.1. Emulsification
4.2. Immune Animal and Method
4.3. Cross-Reactivity
4.4. Extraction of DON
4.5. FHB-Resistant Wheat Cultivars
5. Conclusions
Acknowledgements
References
- Council for Agricultural Science and Technology, Mycotoxins: Risks in Plants, Animals and Human Systems. Report No. 139; Council for Agricultural Science and Technology: Ames, IA, USA, January, 2003.
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. J.Mol. Sci. 2009, 10, 147–173. [Google Scholar]
- Ueno, Y.; Hosoya, M.; Morita, Y.; Ueno, I.; Tatsuno, T. Inhibition of the protein synthesis in rabbit reticulocyte by nivalenol, a toxic principle isolated from Fusarium nivale-growing rice. J. Biochem. 1968, 64, 479–485. [Google Scholar] [PubMed]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar]
- Guidance for Industry and FDA: Letter to State Agricultural Directors; State Feed Control Officials, and Food, Feed and Grain Trade Organizations. 1993. Available online: http://www.cfsan.fda.gov/_dms/graingui.html (accessed on 26 April 2006).
- Ueno, Y.; Sato, N.; Ishii, K.; Sakai, K.; Tsunoda, H. Biological and chemical detection of trichothecene mycotoxins of Fusarium species. Appl. Microbiol. 1973, 25, 699–704. [Google Scholar] [PubMed]
- Ueno, Y.; Nakajima, M.; Sakai, K.; Ishii, K.; Sato, N. Comparative toxicology of trichothec mycotoxins: Inhibition of protein synthesis in animal cells. J. Biochem. 1973, 74, 285–296. [Google Scholar]
- Council for Agricultural Science and Technology, Mycotoxins: Risks in Plants, Animals and Human Systems. Report No. 139; Council for Agricultural Science and Technology: Ames, IA, USA, January, 2003; p. 1999.
- Scott, P.M.; Kanhere, S.R. Comparison of column phases for separation of derivatized trichothecenes by capillary gas chromatography. J. Chromatogr. 1986, 368, 374–380. [Google Scholar]
- Acke, B.K.; Casper, H.H. Determination of Deoxynivalenol in wheat, barley, and malt by column cleanup and gas chromatography with electron capture detection. J. AOAC Int. 1996, 79, 472–475. [Google Scholar] [PubMed]
- Trucksess, M.W.; Page, S.W.; Wood, G.E.; Choth, T.H. Determination of Deoxynivalenol in white flour, whole wheat flour, and bran by solid-phase extraction/liquid chromatography: Interlaboratory study. J. AOAC Int. 1998, 81, 880–886. [Google Scholar] [PubMed]
- Jestoi, M.; Ritieni, A.; Rizzo, A. Analysis of the Fusarium mycotoxins fusaproliferin and trichothecenes in grains using gas chromatography-mass spectrometry. J. Agric. Food. Chem. 2004, 52, 1464–1469. [Google Scholar]
- Klotzel, M.; Gutsche, B.; Lauber, U.; Humpf, H.U. Determination of 12 type A and B trichothecenes in cereals by liquid chromatography-electrospray ionization tandem mass spectrometry. J. Agric. Food. Chem. 2005, 53, 8904–8910. [Google Scholar]
- Schneider, E.; Curtui, V.; Seidler, C.; Dietrich, R.; Usleber, E.; Märtlbauer, E. Rapid methods for deoxynivalenol and other trichothecenes. Toxicol. Lett. 2004, 153, 113–121. [Google Scholar]
- Barna-Vetro, I.; Gyongyosi, A.; Solt, L. Monoclonal antibody-based enzyme-linked immunosorbent assay of Fusarium T-2 and zearalenone toxins in cereals. Appl. Environ. Microbiol. 1994, 60, 729–731. [Google Scholar]
- Casale, W.L.; Pestka, J.J.; Hart, L.P. Enzyme-linked immunosorbent assay employing monoclonal antibody specific for Deoxynivalenol (vomitoxin) and several analogues. J. Agric. Food. Chem. 1988, 36, 663–668. [Google Scholar] [CrossRef]
- Abraham, G.E. Radioimmunoassay of steroids in biological fluids. J. Steroid Biochem. 1975, 6, 261–270. [Google Scholar]
- Park, J.J.; Chu, F.S. Assessment of immunochemical methods for the analysis of trichothecene mycotoxins in naturally occurring moldy corn. J. AOAC Int. 1996, 79, 465–471. [Google Scholar]
- Pestka, J.J.; Islam, Z.; Amuzie, C.J. Immunochemical assessment of Deoxynivalenol tissue distribution following oral exposure in the mouse. Toxicol. Lett. 2008, 178, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.G.; Hu, S.C.; Wei, P.H. Technology of Immunoassay; Nanjing University Press: Nanjing, China, 1998; pp. 279–284. (in Chinese) [Google Scholar]
- Chen, X.J.; Chen, M.Y.; Zhao, Y.J. Application of Immunological Techniques in Plant Science; China Agricultural University Press: Beijing, China, 1998. (in Chinese) [Google Scholar]
- Wang, Y.Z.; Miller, J.D. Toxin producing potential of Fusarium graminearum from China. MycoSystema 1994, 13, 229–234. (in Chinese). [Google Scholar]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Alexander, N.J.; McCormick, S.P.; Waalwijk, C.; Lee, T.; Proctor, R.H. The genetic basis for 3-ADON and 15-ADON trichothecene chemotypes in Fusarium. Fungal Genet Biol. 2011, 48, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Mesterhazy, A.; Krska, R.; Ruckenbauer, P. The ability to detoxify the mycotoxin deoxynivalenol colocalizes with a major quantitative trait locus for Fusarium head blight resistance in wheat. Mol. Plant Microbe Interact. 2005, 18, 1318–1324. [Google Scholar]
- Alexander, N.J.; McCormick, S.P.; Ziegenhorn, S.L. Phytotoxicity of selected trichothecenes using chlamydomonas reinhardtii as a model system. Nat. Toxins 1999, 7, 265–269. [Google Scholar] [CrossRef] [PubMed]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ji, F.; Li, H.; Xu, J.; Shi, J. Enzyme-Linked Immunosorbent-Assay for Deoxynivalenol (DON). Toxins 2011, 3, 968-978. https://doi.org/10.3390/toxins3080968
Ji F, Li H, Xu J, Shi J. Enzyme-Linked Immunosorbent-Assay for Deoxynivalenol (DON). Toxins. 2011; 3(8):968-978. https://doi.org/10.3390/toxins3080968
Chicago/Turabian StyleJi, Fang, Hua Li, Jianhong Xu, and Jianrong Shi. 2011. "Enzyme-Linked Immunosorbent-Assay for Deoxynivalenol (DON)" Toxins 3, no. 8: 968-978. https://doi.org/10.3390/toxins3080968
APA StyleJi, F., Li, H., Xu, J., & Shi, J. (2011). Enzyme-Linked Immunosorbent-Assay for Deoxynivalenol (DON). Toxins, 3(8), 968-978. https://doi.org/10.3390/toxins3080968