Disintegrins from Hematophagous Sources

Abstract
:1. Introduction
1.1. Disintegrins, Platelets and Thrombus Formation
1.2. Disintegrins, Angiogenesis and Cancer
1.3. Disintegrins from Hematophagous Animals

| Name/reference | Species | Mol wt | IC50 | R/S/P # | Tripeptide | Cell target | Integrin |
|---|---|---|---|---|---|---|---|
| Ticks | |||||||
| Variabilin [36] | D. variabilis | 5 | 157 nM | N/N/Y | RGD | Platelets | αIIbβ3 |
| ISL929/1373 [37] | Ixodes sp. | 10 | ? | Y/N/Y | ? | Neutrophils | αMβ2 ? |
| Monogrin [38] | A.monolakensis | 10 | 150 nM | Y/N/Y | RGD | Platelets | αIIβ3 |
| TAI [39]* | O. moubata | 15 | 8 nM | N/N/Y | ? | Platelets EC | α2β1, α1β1 |
| Disagregin [40] | O. moubata | 6 | 104 nM | N/N/Y | RED | Platelets | αIIbβ3 |
| Ixodegrin [41]** | Ixodes sp. | 7 | ? | N/N/N | RGD | Platelets? | αIIββ3 |
| Savignygrin [42] | O. savigny | 7 | 130 nM | N/N/Y | RGD | Platelets | αIIbβ3 |
| Horseflies | |||||||
| Tabinhibitin [28] | T. yao | 25 | < 40 nM | N/N/Y | RGD | Platelets | αIIbβ3 |
| Tablysin [29] | T. yao | 25 | 100 nM | Y/Y/Y | RGD | Platelets EC | αIIbβ3 αvβ3 |
| TabRTS [30] | T. yao | 25 | 50 nM | Y/N/Y | RTS | EC | α1β1 |
| Leeches | |||||||
| Decorsin [43] | M. decora | 4 | 500 nM | Y/Y/Y | RGD | Platelets | αIIbβ3 |
| Ornatin [44] | P. ornata | 5 | 130 nM | Y/N/Y | RGD | Platelets | αIIbβ3 |
| Worms | |||||||
| HPI [45] | A. caninum | 20 | ? | Y/N/N | KGD | Platelets? | αIIbβ3? |
| NIF [46] | A. caninum | 41 | <10 nM | Y/N/Y | ? | Neutrophil | αMβ2 |
1.3.1. Ticks
1.3.1.1. Variabilin
1.3.1.2. Disagregin
1.3.1.3. Savignygrin
1.3.1.4. Monogrin
1.3.1.5. Ixodegrin
1.3.1.6. Tick Antiplatelet Inhibitor (TAI)
1.3.1.7. ISL929/1373
1.3.2. Tabanids
1.3.2.1. Tabinhibitin
1.3.2.2. TabRTS
1.3.2.3. Tablysin-15
1.3.3. Leeches
1.3.3.1. Decorsin
1.3.3.2. Ornatin
1.3.4. Worms
1.3.4.1. Hookworm Platelet Inhibitor (HPI)
1.3.4.2. Neutrophil Inhibitory Factor (NIF)
1.4. Construction and Content: Identification of Putative Disintegrins
2. Discussion
2.1. Disintegrins from Bugs (Triatoma and Rhodnius sp.)
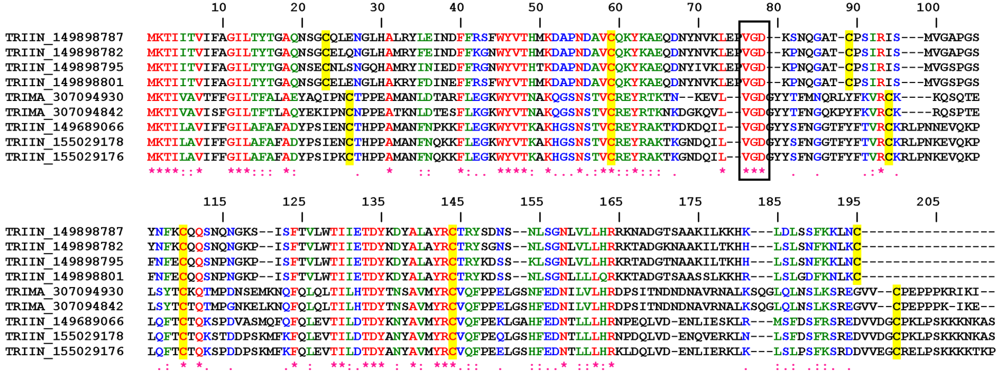
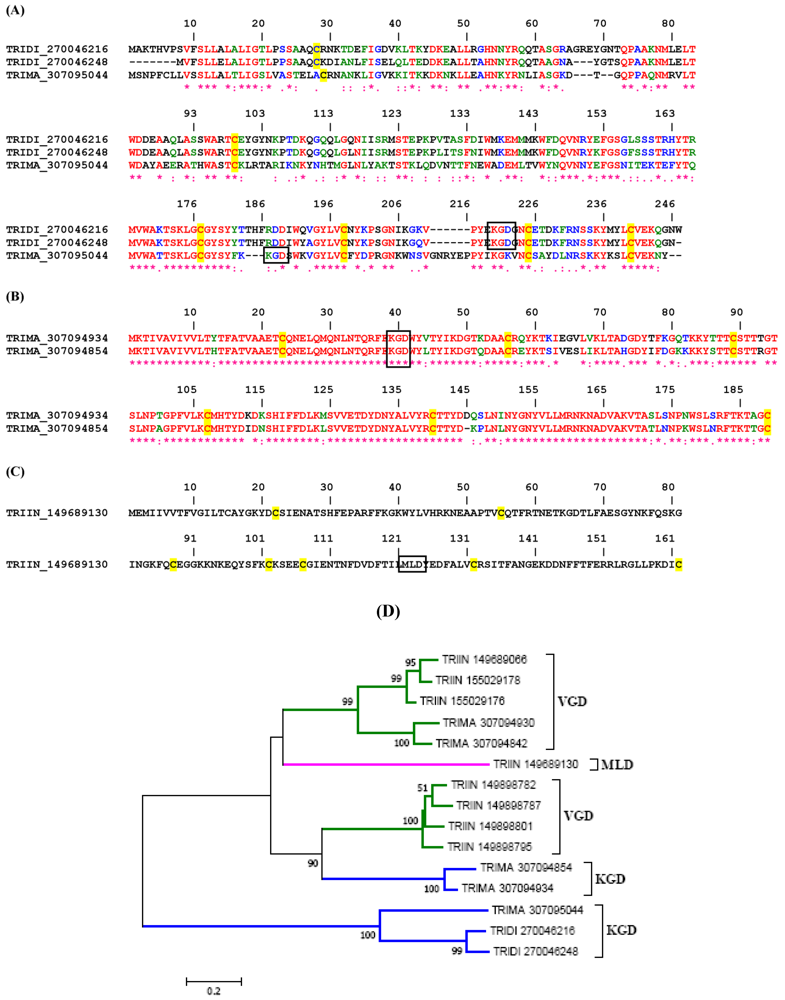
2.1.1. VGD Disintegrin Family
2.1.2. KGD, RGD and MLD Disintegrin Family
2.2. Disintegrins from Metastriate Ticks (Dermacentor, Rhipicephalus, and Amblyomma).
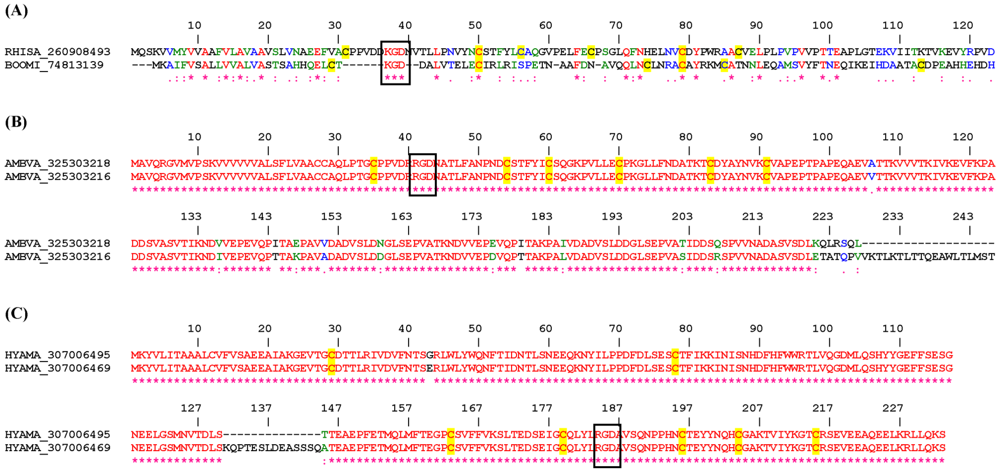
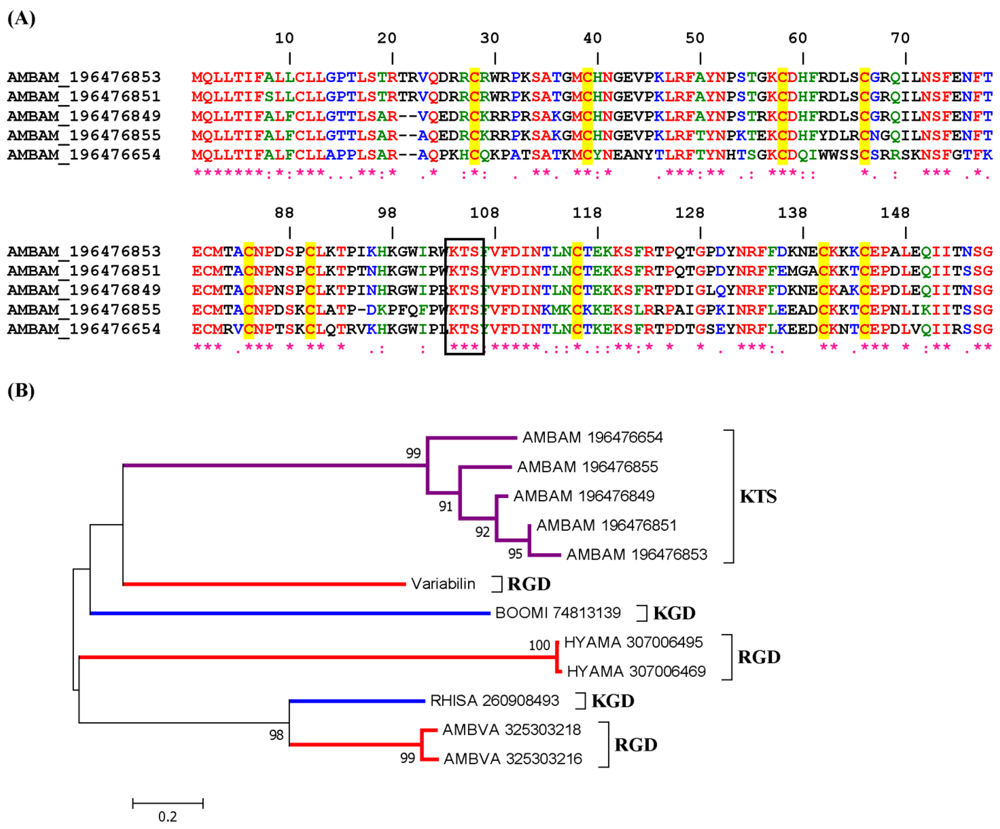
2.2.1. KGD and RGD Disintegrin Family


2.2.2. KTS/RTS Disintegrin Family
2.2.3. Duodegrins

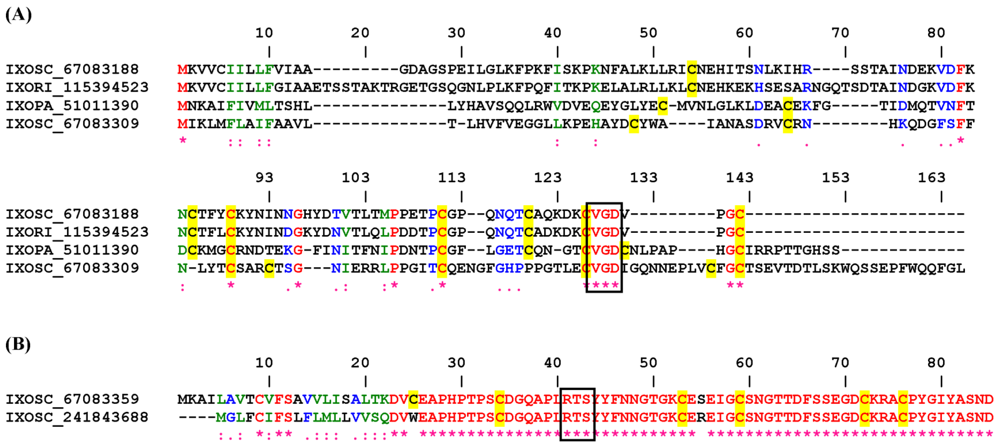
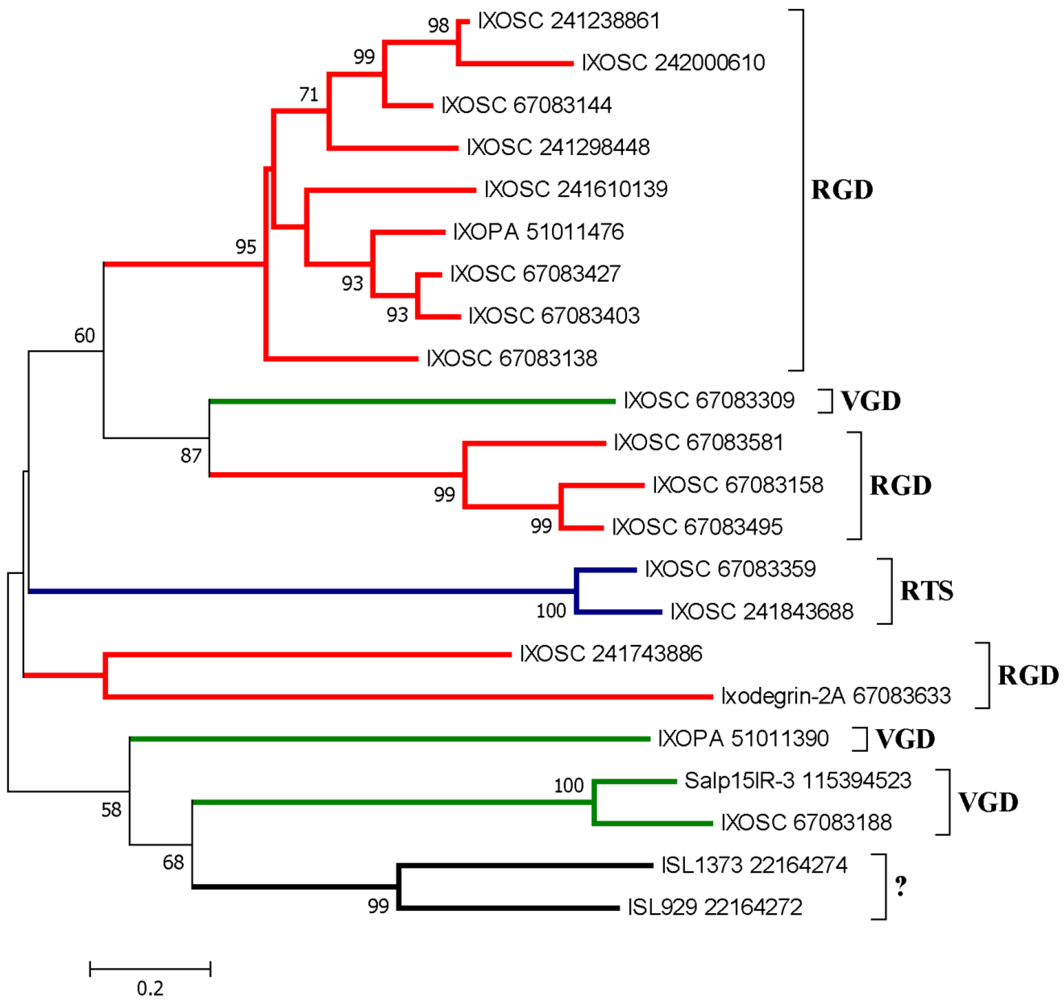
2.3. Disintegrins from Prostriate Ticks (Ixodes sp.).
2.3.1. RGD, KGD, and VGD Disintegrin Family



2.3.2. RTS Disintegrin Family

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
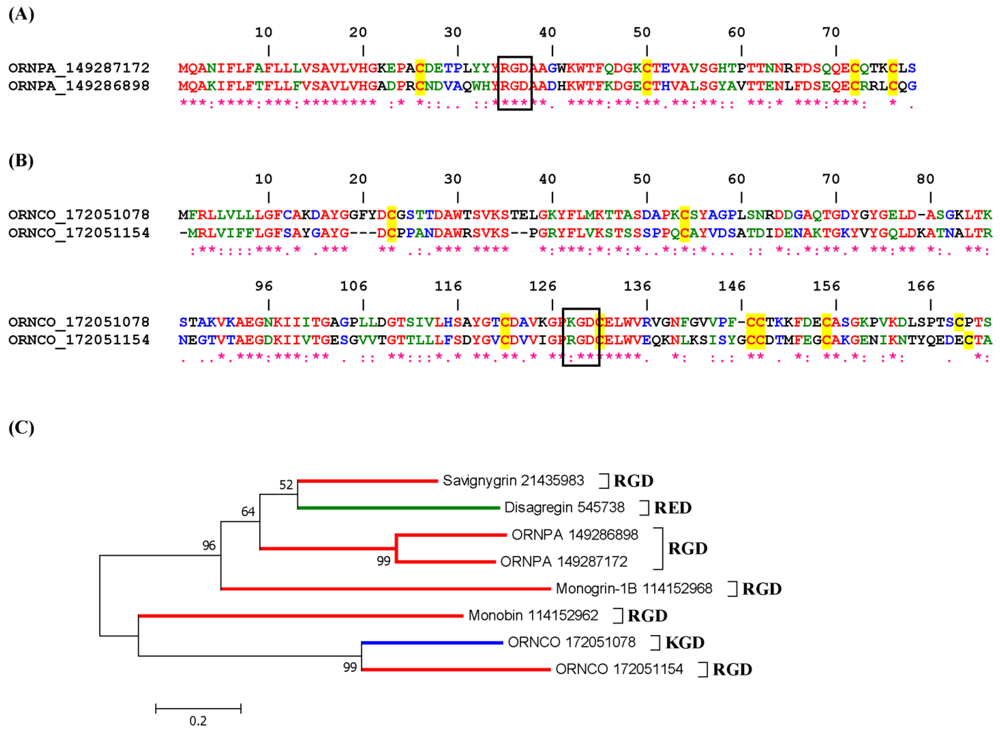
2.3.3. Disintegrins from Ornithodorus sp.

2.3.4. Disintegrins from Fleas

3. Conclusions
Conflict of Interest
Acknowledgments
References
- Avraamides, C.J.; Garmy-Susini, B.; Varner, J.A. Integrins in angiogenesis and lymphangiogenesis. Nat. Rev. Cancer 2008, 8, 604–617. [Google Scholar]
- Hood, J.D.; Cheresh, D.A. Role of integrins in cell invasion and migration. Nat. Rev. Cancer 2002, 2, 91–100. [Google Scholar]
- Silva, R.; D’Amico, G.; Hodivala-Dilke, K.M.; Reynolds, L.E. Integrins: The keys to unlocking angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1703–1713. [Google Scholar]
- Huang, T.F. What have snakes taught us about integrins? Cell. Mol. Life Sci. 1998, 54, 527–540. [Google Scholar] [CrossRef]
- McLane, M.A.; Joerger, T.; Mahmoud, A. Disintegrins in health and disease. Front Biosci. 2008, 13, 6617–6637. [Google Scholar]
- Fox, J.W.; Serrano, S.M. Insights into and speculations about snake venom metalloproteinase (SVMP) synthesis, folding and disulfide bond formation and their contribution to venom complexity. FEBS J. 2008, 275, 3016–3030. [Google Scholar]
- Kini, R.M.; Evans, H.J. Structural domains in venom proteins: Evidence that metalloproteinases and nonenzymatic platelet aggregation inhibitors (disintegrins) from snake venoms are derived by proteolysis from a common precursor. Toxicon 1992, 30, 265–293. [Google Scholar]
- Calvete, J.J. Brief history and molecular determinants of snake venom disintegrin evolution. In Toxins and Hemostasis from Bench to Bedside; Kini, R.M., Clemetson, K., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: New York, NY, USA, 2010; pp. 285–300. [Google Scholar]
- Walsh, E.M.; Marcinkiewicz, C. Non-RGD-containing snake venom disintegrins, functional and structural relations. Toxicon 2011, 58, 355–362. [Google Scholar]
- Francischetti, I.M. Platelet aggregation inhibitors from hematophagous animals. Toxicon 2010, 56, 1130–1144. [Google Scholar]
- Swenson, S.; Minea, R.; Zidovetzki, S.; Helchowski, C.; Costa, F.; Markland, F.S. Anti-Angiogenesis and disintegrins. In Toxins and Hemostasis from Bench to Bedside; Kini, R.M., Clemetson, K., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: New York, NY, USA, 2010; pp. 301–329. [Google Scholar]
- Eble, J.A. Matrix biology meets toxinology. Matrix Biol. 2010, 29, 239–247. [Google Scholar]
- McLane, M.A.; Marcinkiewicz, C.; Vijay-Kumar, S.; Wierzbicka-Patynowski, I.; Niewiarowski, S. Viper venom disintegrins and related molecules. Proc. Soc. Exp. Biol. Med. 1998, 219, 109–119. [Google Scholar]
- Swenson, S.; Ramu, S.; Markland, F.S. Anti-Angiogenesis and RGD-containing snake venom disintegrins. Curr. Pharm. Des. 2007, 13, 2860–2871. [Google Scholar]
- Coelho, A.L.; De Freitas, M.S.; Mariano-Oliveira, A.; Rapozo, D.C.; Pinto, L.F.; Niewiarowski, S.; Zingali, R.B.; Marcinkiewicz, C.; Barja-Fidalgo, C. RGD- and MLD-disintegrins, jarastatin and EC3, activate integrin-mediated signaling modulating the human neutrophils chemotaxis, apoptosis and IL-8 gene expression. Exp. Cell Res. 2004, 292, 371–384. [Google Scholar] [CrossRef]
- Sanz, L.; Chen, R.Q.; Perez, A.; Hilario, R.; Juarez, P.; Marcinkiewicz, C.; Monleon, D.; Celda, B.; Xiong, Y.L.; Perez-Paya, E.; Calvete, J.J. cDNA cloning and functional expression of jerdostatin, anovel RTS-disintegrin from Trimeresurus jerdonii and a specific antagonist of the alpha1beta1 integrin. J. Biol. Chem. 2005, 280, 40714–40722. [Google Scholar]
- Barja-Fidalgo, C.; Coelho, A.L.; Saldanha-Gama, R.; Helal-Neto, E.; Mariano-Oliveira, A.; Freitas, M.S. Disintegrins: Integrin selective ligands which activate integrin-coupled signaling and modulate leukocyte functions. Braz. J. Med. Biol. Res. 2005, 38, 1513–1520. [Google Scholar]
- Calvete, J.J.; Marcinkiewicz, C.; Monleon, D.; Esteve, V.; Celda, B.; Juarez, P.; Sanz, L. Snake venom disintegrins: Evolution of structure and function. Toxicon 2005, 45, 1063–1074. [Google Scholar]
- Huang, T.F. The discovery of disintegrins. In Toxins and Hemostasis from Bench to Bedside; Kini, R.M., Clemetson, K., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: New York, NY, USA, 2010; pp. 269–284. [Google Scholar]
- Watson, S.P.; Auger, J.M.; McCarty, O.J.; Pearce, A.C. GPVI and integrin alphaIIb beta3 signaling in platelets. J. Thromb. Haemost. 2005, 3, 1752–1762. [Google Scholar]
- Kasirer-Friede, A.; Kahn, M.L.; Shattil, S.J. Platelet integrins and immunoreceptors. Immunol. Rev. 2007, 218, 247–264. [Google Scholar]
- Huang, T.F.; Holt, J.C.; Lukasiewicz, H.; Niewiarowski, S.T. A low molecular weight peptide inhibiting fibrinogen interaction with platelet receptors expressed on glycoprotein IIb-IIIa complex. J. Biol. Chem. 1987, 262, 16157–16163. [Google Scholar]
- Clemetson, K.J.; Kini, R.M. Introduction. In Toxins and Hemostasis from Bench to Bedside; Kini, R.M., Clemetson, K., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: New York, NY, USA, 2010; pp. 1–8. [Google Scholar]
- Zhou, Q.; Nakada, M.T.; Arnold, C.; Shieh, K.Y.; Markland, F.S.J. Contortrostatin, a dimeric disintegrin from Agkistrodon contortrix contortrix, inhibits angiogenesis. Angiogenesis 1999, 3, 259–269. [Google Scholar] [CrossRef]
- Marcinkiewicz, C.; Weinreb, P.H.; Calvete, J.J.; Kisiel, D.G.; Mousa, S.A.; Tuszynski, G.P.; Lobb, R.R. Obtustatin: A potent selective inhibitor of alpha1beta1 integrin in vitro and angiogenesis in vivo. Cancer Res. 2003, 63, 2020–2023. [Google Scholar]
- Kusuma, N.; Denoyer, D.; Eble, J.A.; Redvers, R P.; Parker, B.S.; Pelzer, R.; Anderson, R.L.; Pouliot, N. Integrin-dependent response to laminin-511 regulates breast tumor cell invasion and metastasis. Int. J. Cancer 2012, 130, 555–566. [Google Scholar] [CrossRef]
- Francischetti, I.M.; Mather, T.N.; Ribeiro, J.M. Tick saliva is a potent inhibitor of endothelial cell proliferation and angiogenesis. Thromb. Haemost. 2005, 94, 167–174. [Google Scholar]
- Ma, D.; Wang, Y.; Yang, H.; Wu, J.; An, S.; Gao, L.; Xu, X.; Lai, R. Anti-Thrombosis repertoire of blood-feeding horsefly salivary glands. Mol. Cell Proteomics 2009, 8, 2071–2079. [Google Scholar]
- Ma, D.; Xu, X.; An, S.; Liu, H.; Yang, X.; Andersen, J.F.; Wang, Y.; Tokumasu, F.; Ribeiro, J.M.; Francischetti, I.M.; Lai, R. A novel family of RGD-containing disintegrins (Tablysin-15) from the salivary gland of the horsefly Tabanus yao targets alphaIIbbeta3 or alphaVbeta3 and inhibits platelet aggregation and angiogenesis. Thromb. Haemost. 2011, 105, 1032–1045. [Google Scholar]
- Ma, D.; Gao, L.; An, S.; Song, Y.; Wu, J.; Xu, X.; Lai, R. A horsefly saliva antigen 5-like protein containing RTS motif is an angiogenesis inhibitor. Toxicon 2010, 55, 45–51. [Google Scholar]
- Ribeiro, J.M.; Francischetti, I.M. Role of arthropod saliva in blood feeding: Sialome and post-sialome perspectives. Annu. Rev. Entomol. 2003, 48, 73–88. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; Renjifo, C.; de la Vega, R.C. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar]
- Steen, N.A.; Barker, S.C.; Alewood, P.F. Proteins in the saliva of the Ixodida (ticks): Pharmacological features and biological significance. Toxicon 2006, 47, 1–20. [Google Scholar]
- Mans, B.J.; Francischetti, I.M.B. Sialomic perspectives on the evolution of blood-feeding behavior in arthropods: Future therapeutics by natural design. In Toxins and Hemostasis from Bench to Bedside; Kini, R.M., Clemetson, K., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: New York, NY, USA, 2010; pp. 21–44. [Google Scholar]
- Francischetti, I.M.; Sa-Nunes, A.; Mans, B.J.; Santos, I.M.; Ribeiro, J.M. The role of saliva in tick feeding. Front Biosci. 2009, 14, 2051–2088. [Google Scholar]
- Wang, X.; Coons, L.B.; Taylor, D.B.; Stevens, S.E.J.; Gartner, T.K. Variabilin, a novel RGD-containing antagonist of glycoprotein IIb-IIIa and platelet aggregation inhibitor from the hard tick Dermacentor variabilis. J. Biol. Chem. 1996, 271, 17785–17790. [Google Scholar]
- Guo, X.; Booth, C.J.; Paley, M.A.; Wang, X.; DePonte, K.; Fikrig, E.; Narasimhan, S.; Montgomery, R.R. Inhibition of neutrophil function by two tick salivary proteins. Infect Immun. 2009, 77, 2320–2329. [Google Scholar]
- Mans, B.J.; Andersen, J.F.; Schwan, T.G.; Ribeiro, J.M. Characterization of anti-hemostatic factors in the argasid, Argas monolakensis: Implications for the evolution of blood-feeding in the soft tick family. Insect Biochem. Mol. Biol. 2008, 38, 22–41. [Google Scholar]
- Karczewski, J.; Waxman, L.; Endris, R.G.; Connolly, T.M. An inhibitor from the argasid tick Ornithodoros moubata of cell adhesion to collagen. Biochem. Biophys. Res. Commun. 1995, 208, 532–541. [Google Scholar]
- Karczewski, J.; Endris, R.; Connolly, T.M. Disagregin is a fibrinogen receptor antagonist lacking the Arg-Gly-Asp sequence from the tick, Ornithodoros moubata. J. Biol. Chem. 1994, 269, 6702–6708. [Google Scholar]
- Francischetti, I.M.; My-Pham, V.; Mans, B.J.; Andersen, J.F.; Mather, T.N.; Lane, R.S.; Ribeiro, J.M. The transcriptome of the salivary glands of the female western black-legged tick Ixodes pacificus (Acari: Ixodidae). Insect Biochem. Mol. Biol. 2005, 35, 1142–1161. [Google Scholar]
- Mans, B.J.; Louw, A.I.; Neitz, A.W. Savignygrin, a platelet aggregation inhibitor from the soft tick Ornithodoros savignyi, presents the RGD integrin recognition motif on the Kunitz-BPTI fold. J. Biol. Chem. 2002, 277, 21371–21378. [Google Scholar]
- Seymour, J.L.; Henzel, W.J.; Nevins, B.; Stults, J.T.; Lazarus, R.A. A potent glycoprotein IIb-IIIa antagonist and platelet aggregation inhibitor from the leech Macrobdella decora. J. Biol. Chem. 1990, 265, 10143–10147. [Google Scholar]
- Mazur, P.; Henzel, W.J.; Seymour, J.L.; Lazarus, R.A. Ornatins: Potent glycoprotein IIb-IIIa antagonists and platelet aggregation inhibitors from the leech Placobdella ornata. Eur. J. Biochem. 1991, 202, 1073–1082. [Google Scholar]
- Del Valle, A.; Jones, B.F.; Harrison, L.M.; Chadderdon, R.C.; Cappello, M. Isolation and molecular cloning of a secreted hookworm platelet inhibitor from adult Ancylostoma caninum. Mol. Biochem. Parasitol. 2003, 129, 167–177. [Google Scholar]
- Moyle, M.; Foster, D.L.; McGrath, D.E.; Brown, S.M.; Laroche, Y.; De Meutter, J.; Stanssens, P.; Bogowitz, C.A.; Fried, V.A.; Ely, J.A. A hookworm glycoprotein that inhibits neutrophil function is a ligand of the integrin CD11b/CD18. J. Biol. Chem. 1994, 269, 10008–10015. [Google Scholar]
- Karczewski, J.; Connolly, T.M. The interaction of disagregin with the platelet fibrinogen receptor, glycoprotein IIb-IIIa. Biochem. Biophys. Res. Commun. 1997, 241, 744–748. [Google Scholar]
- Francischetti, I.M.; Meng, Z.; Mans, B.J.; Gudderra, N.; Hall, M.; Veenstra, T.D.; Pham, V.M.; Kotsyfakis, M.; Ribeiro, J.M. An insight into the salivary transcriptome and proteome of the soft tick and vector of epizootic bovine abortion, Ornithodoros coriaceus. J. Proteomics 2008, 71, 493–512. [Google Scholar]
- Ribeiro, J.M.; Anderson, J.M.; Manoukis, N.C.; Meng, Z.; Francischetti, I.M. A further insight into the sialome of the tropical bont tick, Amblyomma variegatum. BMC Genomics 2011, 12, 136. [Google Scholar]
- Xu, X.; Francischetti, I.M.; Lai, R.; Ribeiro, J.M.; Andersen, J.F. Structure of a protein having inhibitory disintegrin and leukotriene scavenging functions contained in a single domain. J. Biol. Chem. 2012, in press. [Google Scholar]
- Min, G.S.; Sarkar, I.N.; Siddall, M.E. Salivary transcriptome of the North American medicinal leech, Macrobdella decora. J. Parasitol. 2010, 96, 1211–1221. [Google Scholar]
- Krezel, A.M.; Wagner, G.; Seymour-Ulmer, J.; Lazarus, R.A. Structure of the RGD protein decorsin: Conserved motif and distinct function in leech proteins that affect blood clotting. Science 1994, 264, 1944–1947. [Google Scholar]
- Mazur, P.; Dennis, M.S.; Seymour, J.L.; Lazarus, R.A. Expression, purification, and characterization of recombinant ornatin E, a potent glycoprotein IIb-IIIa antagonist. Protein Expr. Purif. 1993, 4, 282–289. [Google Scholar] [CrossRef]
- Muchowski, P.J.; Zhang, L.; Chang, E.R.; Soule, H.R.; Plow, E.F.; Moyle, M. Functional interaction between the integrin antagonist neutrophil inhibitory factor and the I domain of CD11b/CD18. J. Biol. Chem. 1994, 269, 26419–26423. [Google Scholar]
- Rieu, P.; Ueda, T.; Haruta, I.; Sharma, C.P.; Arnaout, M.A. The A-domain of beta 2 integrin CR3 (CD11b/CD18) is a receptor for the hookworm-derived neutrophil adhesion inhibitor NIF. J. Cell Biol. 1994, 127, 2081–2091. [Google Scholar]
- Zhou, M.Y.; Lo, S.K.; Bergenfeldt, M.; Tiruppathi, C.; Jaffe, A.; Xu, N.; Malik, A.B. In vivo expression of neutrophil inhibitory factor via gene transfer prevents lipopolysaccharide-induced lung neutrophil infiltration and injury by a beta2 integrin-dependent mechanism. J. Clin. Invest. 1998, 101, 2427–2437. [Google Scholar] [CrossRef]
- Nielsen, H.; Brunak, S.; von Heijne, G. Machine learning approaches for the prediction of signal peptides and other protein sorting signals. Protein Eng. 1999, 12, 3–9. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar]
- Ribeiro, J.M.; Alarcon-Chaidez, F.; Francischetti, I.M.; Mans, B.J.; Mather, T.N.; Valenzuela, J.G.; Wikel, S.K. An annotated catalog of salivary gland transcripts from Ixodes scapularis ticks. Insect Biochem. Mol. Biol. 2006, 36, 111–129. [Google Scholar]
- Francischetti, I.M.; Calvo, E.; Andersen, J.F.; Pham, V.M.; Favreau, A.J.; Barbian, K.D.; Romero, A.; Valenzuela, J.G.; Ribeiro, J.M. Insight into the Sialome of the Bed Bug, Cimex lectularius. J. Proteome Res. 2010, 9, 3820–3831. [Google Scholar]
- Andersen, J.F.; Hinnebusch, B.J.; Lucas, D.A.; Conrads, T.P.; Veenstra, T.D.; Pham, V.M.; Ribeiro, J.M. An insight into the sialome of the oriental rat flea, Xenopsylla cheopis (Rots). BMC Genomics 2007, 8, 102. [Google Scholar]
- Francischetti, I.M.; Valenzuela, J.G.; Pham, V.M.; Garfield, M.K.; Ribeiro, J.M. Toward a catalog for the transcripts and proteins (sialome) from the salivary gland of the malaria vector Anopheles gambiae. J. Exp. Biol. 2002, 205, 2429–2451. [Google Scholar]
- Assumpcao, T.C.; Charneau, S.; Santiago, P.B.; Francischetti, I.M.; Meng, Z.; Araujo, C.N.; Pham, V.M.; Queiroz, R.M.; de Castro, C.N.; Ricart, C.A.; Santana, J.M.; Ribeiro, J.M. Insight into the salivary transcriptome and proteome of Dipetalogaster maxima. J. Proteome Res. 2011, 10, 669–679. [Google Scholar]
- Assumpcao, T.C.; Francischetti, I.M.; Andersen, J.F.; Schwarz, A.; Santana, J.M.; Ribeiro, J.M. An insight into the sialome of the blood-sucking bug Triatoma infestans, a vector of Chagas’ disease. Insect Biochem. Mol. Biol. 2008, 38, 213–232. [Google Scholar]
- Andersen, J.F. Structure and mechanism in salivary proteins from blood-feeding arthropods. Toxicon 2010, 56, 1120–1129. [Google Scholar]
- Noeske-Jungblut, C.; Haendler, B.; Donner, P.; Alagon, A.; Possani, L.; Schleuning, W.D. Triabin, a highly potent exosite inhibitor of thrombin. J. Biol. Chem. 1995, 270, 28629–28634. [Google Scholar]
- Isawa, H.; Orito, Y.; Jingushi, N.; Iwanaga, S.; Morita, A.; Chinzei, Y.; Yuda, M. Identification and characterization of plasma kallikrein-kinin system inhibitors from salivary glands of the blood-sucking insect Triatoma infestans. FEBS J. 2007, 274, 4271–4286. [Google Scholar]
- Willadsen, P.; Riding, G.A.; McKenna, R.V.; Kemp, D.H.; Tellam, R.L.; Nielsen, J.N.; Lahnstein, J.; Cobon, G.S.; Gough, J.M. Immunologic control of a parasitic arthropod. Identification of a protective antigen from Boophilus Microplus. J. Immunol. 1989, 143, 1346–1351. [Google Scholar]
- Willadsen, P.; McKenna, R.V. Vaccination with “concealed” antigens: Myth or reality? Parasite Immunol. 1991, 13, 605–616. [Google Scholar] [CrossRef]
- Juncadella, I.J.; Anguita, J. The immunosuppresive tick salivary protein, Salp15. Adv. Exp. Med. Biol. 2009, 666, 121–131. [Google Scholar]
- Mans, B.J.; Andersen, J.F.; Francischetti, I.M.; Valenzuela, J.G.; Schwan, T.G.; Pham, V.M.; Garfield, M.K.; Hammer, C.H.; Ribeiro, J.M. Comparative sialomics between hard and soft ticks: Implications for the evolution of blood-feeding behavior. Insect Biochem. Mol. Biol. 2008, 38, 42–58. [Google Scholar]
- Valenzuela, J.G.; Francischetti, I.M.; Pham, V.M.; Garfield, M.K.; Mather, T.N.; Ribeiro, J.M. Exploring the sialome of the tick Ixodes scapularis. J. Exp. Biol. 2002, 205, 2843–2864. [Google Scholar]
- Francischetti, I.M.; Mans, B.J.; Meng, Z.; Gudderra, N.; Veenstra, T.D.; Pham, V.M.; Ribeiro, J.M. An insight into the sialome of the soft tick, Ornithodorus parkeri. Insect Biochem. Mol. Biol. 2008, 38, 1–21. [Google Scholar]
- Wermelinger, L.S.; Geraldo, R.B.; Frattani, F.S.; Rodrigues, C.R.; Juliano, M.A.; Castro, H.C.; Zingali, R.B. Integrin inhibitors from snake venom: Exploring the relationship between the structure and activity of RGD-peptides. Arch. Biochem. Biophys. 2009, 482, 25–32. [Google Scholar]
- Bilgrami, S.; Tomar, S.; Yadav, S.; Kaur, P.; Kumar, J.; Jabeen, T.; Sharma, S.; Singh, T.P. Crystal structure of schistatin, a disintegrin homodimer from saw-scaled viper (Echis carinatus) at 2.5 A resolution. J. Mol. Biol. 2004, 341, 829–837. [Google Scholar] [CrossRef]
- Fujii, Y.; Okuda, D.; Fujimoto, Z.; Horii, K.; Morita, T.; Mizuno, H. Crystal structure of trimestatin, a disintegrin containing a cell adhesion recognition motif RGD. J. Mol. Biol. 2003, 332, 1115–1122. [Google Scholar]
Supplementary Files and Correction
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Assumpcao, T.C.F.; Ribeiro, J.M.C.; Francischetti, I.M.B. Disintegrins from Hematophagous Sources. Toxins 2012, 4, 296-322. https://doi.org/10.3390/toxins4050296
Assumpcao TCF, Ribeiro JMC, Francischetti IMB. Disintegrins from Hematophagous Sources. Toxins. 2012; 4(5):296-322. https://doi.org/10.3390/toxins4050296
Chicago/Turabian StyleAssumpcao, Teresa C. F., José M. C. Ribeiro, and Ivo M. B. Francischetti. 2012. "Disintegrins from Hematophagous Sources" Toxins 4, no. 5: 296-322. https://doi.org/10.3390/toxins4050296