Toxins VapC and PasB from Prokaryotic TA Modules Remain Active in Mammalian Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Properties of Addictive Modules Chosen for Study: PasAB and VapBC
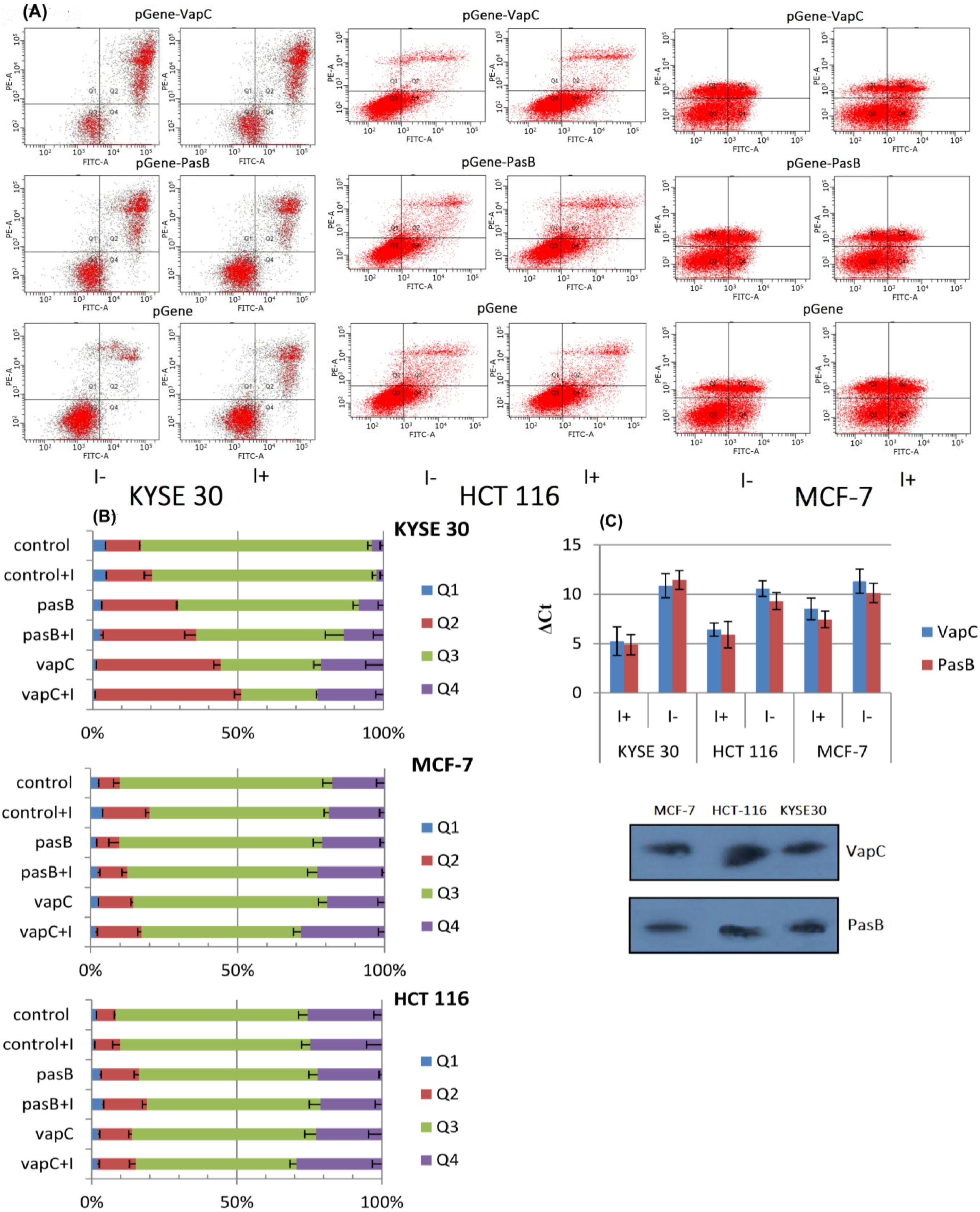
2.2. Effect of Induced Toxin Expression in Human Cancer Cell Lines


2.3. Toxin Expression with the EGFP Tracking System


2.4. Cell Cycle Analysis

3. Experimental Section
3.1. Gene and Cell Line Sources
3.2. Construct Preparation
3.3. Cell Cultures
3.4. Stable Transfection with Regulatory Vector pSwitch
3.5. Transfection with the pGene Vector and Induction
3.6. Transfection with the pBud4.1CE System
3.7. Apoptosis, Necrosis and Viable Cell Assay
3.8. Cell Cycle Assay
3.9. Gene Expression Assays
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Guglielmini, J.; van Melderen, L. Bacterial toxin-antitoxin systems translation inhibitors everywhere. Mob. Genet. Elements 2011, 1, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F. Toxins-antitoxins: Plasmid maintenance, programmed cell death, and cell cycle arrest. Science 2003, 301, 1496–1499. [Google Scholar] [CrossRef] [PubMed]
- Fineran, P.C.; Blower, T.R.; Foulds, I.J.; Humphreys, D.P.; Lilley, K.S.; Salmond, G.P. The phage abortive infection system, ToxIN, functions as a protein–RNA toxin–antitoxin pair. Proc. Natl. Acad. Sci. USA 2009, 106, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Awano, N.; Inouye, M. YeeV is an Escherichia coli toxin that inhibits cell division by targeting the cytoskeleton proteins, FtsZ and MreB. Mol. Microbiol. 2011, 79, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Tan, Q.; Awano, N.; Wu, K.P.; Inouye, M. YeeU enhances the bundling of cytoskeletal polymers of MreB and FtsZ, antagonizing the CbtA (YeeV) toxicity in Escherichia coli. Mol. Microbiol. 2012, 84, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lord, D.M.; Cheng, H.Y.; Osbourne, D.O.; Hong, S.H.; Sanchez-Torres, V.; Quiroga, C.; Zheng, K.; Herrmann, T.; Peti, W. A new type V toxin-antitoxin system where mRNA for toxin GhoT is cleaved by antitoxin GhoS. Nat. Chem. Biol. 2012, 8, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Sberro, H.; Leavitt, A.; Kiro, R.; Koh, E.; Peleg, Y.; Qimron, U.; Sorek, R. Discovery of functional toxin/antitoxin systems in bacteria by shotgun cloning. Mol. Cell 2013, 50, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Goeders, N.; Van Melderen, L. Toxin-antitoxin systems as multilevel interaction systems. Toxins 2014, 6, 304–324. [Google Scholar] [CrossRef] [PubMed]
- Cook, G.M.; Robson, J.R.; Frampton, R.A.; McKenzie, J.; Przybilski, R.; Fineran, P.C.; Arcus, V.L. Ribonucleases in bacterial toxin–antitoxin systems. Biochim. Biophys. Acta 2013, 1829, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K. Toxin-antitoxin modules may regulate synthesis of macromolecules during nutritional stress. J. Bacteriol. 2000, 182, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Audoly, G.; Vincentell, R.; Edouard, S.; Georgiades, K.; Mediannikov, O.; Gimenez, G.; Socolovschi, C.; Mège, J.L.; Cambillau, C.; Raoult, D. Effect of Rickettsial Toxin VapC on Its Eukaryotic Host. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Walker, A.N.; Daines, D.A. Toxin-antitoxin loci vapBC-1 and vapXD contribute to survival and virulence in nontypeable Haemophilus influenza. BMC Microbiol. 2012, 12. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.A.; Gerdes, K.; Tunnaclife, A. Bacterial toxin RelE induces apoptosis in human cell. FEBS Lett. 2002, 22, 191–194. [Google Scholar] [CrossRef]
- De la Cueva-Méndez, G.; Mills, A.D.; Clay-Farrace, L.; Díaz-Orejas, R.; Laskey, R.A. Regulatable killing of eukaryotic cells by the prokaryotic proteins Kid and Kis. EMBO J. 2003, 22, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Zhang, Y.; Chan, M.C.; Mal, T.K.; Hoeflich, K.P.; Inouye, M.; Ikura, M. Characterization of dual substrate binding sites in the homodimeric structure of Escherichia coli mRNA interferase MazF. J. Mol. Biol. 2006, 357, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Kamada, K.; Hanaoka, F.; Burley, S.K. Crystal structure of the MazE/MazF complex: Molecular bases of antidote-toxin recognition. Mol. Cell 2003, 11, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Erental, A.; Sharon, I.; Engelberg-Kulka, H. Two programmed cell death systems in Escherichia coli an apoptotic-like death is inhibited by the mazF mediated death pathway. PLoS Biol. 2012, 10. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; Kędzierska, B. Regulating toxin-antitoxin expression: Controlled detonation of intracellular molecular timebombs. Toxins 2014, 6, 337–358. [Google Scholar] [CrossRef] [PubMed]
- Ghafourian, S.; Good, L.; Sekawi, Z.; Hamat, R.A; Soheili, S.; Sadeghifard, N.; Neela, V. The mazEF toxin-antitoxin system as a novel antibacterial target in Acinetobacter baumannii. Mem. Inst. Oswaldo Cruz 2014. [Google Scholar] [CrossRef]
- Shapira, A.; Benhar, I. Toxin-based therapeutic approaches. Toxins 2010, 2, 2519–2583. [Google Scholar] [PubMed]
- Shapira, A.; Shapira, S.; Gal-Tanamy, M.; Zemel, R.; Tur-Kaspa, R.; Benhar, I. Removal of hepatitis C virus-infected cells by a zymogenized bacterial toxin. PLoS One 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Chono, H.; Kawano, Y.; Saito, N.; Tsuda, H.; Inoue, K.; Kato, I.; Mineno, J.; Baba, M. Sustained inhibition of HIV-1 replication by conditional expression of the E. coli—Derived endoribonuclease MazF in CD4+ T cells. Hum. Gene Ther. Methods 2013, 24, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Deane, S.M.; Rawlings, D.E. Plasmid evolution and interaction between the plasmid addiction stability systems of two related broad host-range incQ-like plasmids. J. Bacteriol. 2004, 186, 2123–2133. [Google Scholar] [CrossRef] [PubMed]
- Cherny, I.; Overgaard, M.; Borch, J.; Bram, Y.; Gerdes, K.; Gazit, E. Structural and thermodynamic characterization of the Escherichia coli RelBE toxin-antitoxin system: indication for a functional role of differential stability. Biochemistry 2007, 46, 12152–12163. [Google Scholar] [CrossRef] [PubMed]
- Arcus, V.L.; McKenzie, J.L.; Robson, J.; Cook, G.M. The PIN-domain ribonucleases and the prokaryotic VapBC toxin-antitoxin array. Protein Eng. Des. Sel. 2011, 24, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Ahidjo, B.A.; Kuhnert, D.; McKenzie, J.L.; Machowski, E.E.; Gordhan, B.G.; Arcus, V.; Abrahams, G.L.; Mizrahi, V. VapC toxins from Mycobacterium tuberculosis are ribonucleases that differentially inhibit growth and are neutralized by cognate VapB antitoxins. PLoS One 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Winther, K.S.; Gerdes, K. Enteric virulence associated protein VapC inhibits translation by cleavage of initiator tRNA. Proc. Natl. Acad. Sci. USA 2011, 108, 7403–7407. [Google Scholar] [CrossRef] [PubMed]
- McGuire, W.L.; Chamnes, G.C. Studies on the estrogen receptor in breast cancer. In Receptors for Reproductive Hormones; O’Malley, B.W., Chamnes, G.C., Eds.; Plenum Publishing Corporation: New York, NY, USA, 1973; pp. 113–136. [Google Scholar]
- Andreev, D.; Hauryliuk, V.; Terenin, I.; Dmitriev, S.; Ehrenberg, M.; Shatsky, I. The bacterial toxin RelE induces specific mRNA cleavage in the A site of the eukaryote ribosome. RNA 2008, 14, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Tsai, F.M.; Shyu, R.Y.; Tsai, Y.M.; Wang, C.H.; Jiang, S.Y. G protein-coupled receptor kinase 5 mediates Tazarotene-induced gene 1-induced growth suppression of human colon cancer cells. BMC Cancer 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor: New York, NY, USA, 2012. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wieteska, Ł.; Skulimowski, A.; Cybula, M.; Szemraj, J. Toxins VapC and PasB from Prokaryotic TA Modules Remain Active in Mammalian Cancer Cells. Toxins 2014, 6, 2948-2961. https://doi.org/10.3390/toxins6102948
Wieteska Ł, Skulimowski A, Cybula M, Szemraj J. Toxins VapC and PasB from Prokaryotic TA Modules Remain Active in Mammalian Cancer Cells. Toxins. 2014; 6(10):2948-2961. https://doi.org/10.3390/toxins6102948
Chicago/Turabian StyleWieteska, Łukasz, Aleksander Skulimowski, Magdalena Cybula, and Janusz Szemraj. 2014. "Toxins VapC and PasB from Prokaryotic TA Modules Remain Active in Mammalian Cancer Cells" Toxins 6, no. 10: 2948-2961. https://doi.org/10.3390/toxins6102948