The most striking observation of the present experiment was the pronounced sensitivity of ducks to ergot alkaloids. Ducks responded with adverse effects even to the lowest TEA concentration of 0.6 mg per kg diet. These results suggested that ducks obviously respond more sensitive to TEA than other poultry species.
3.2. Duck Experiment
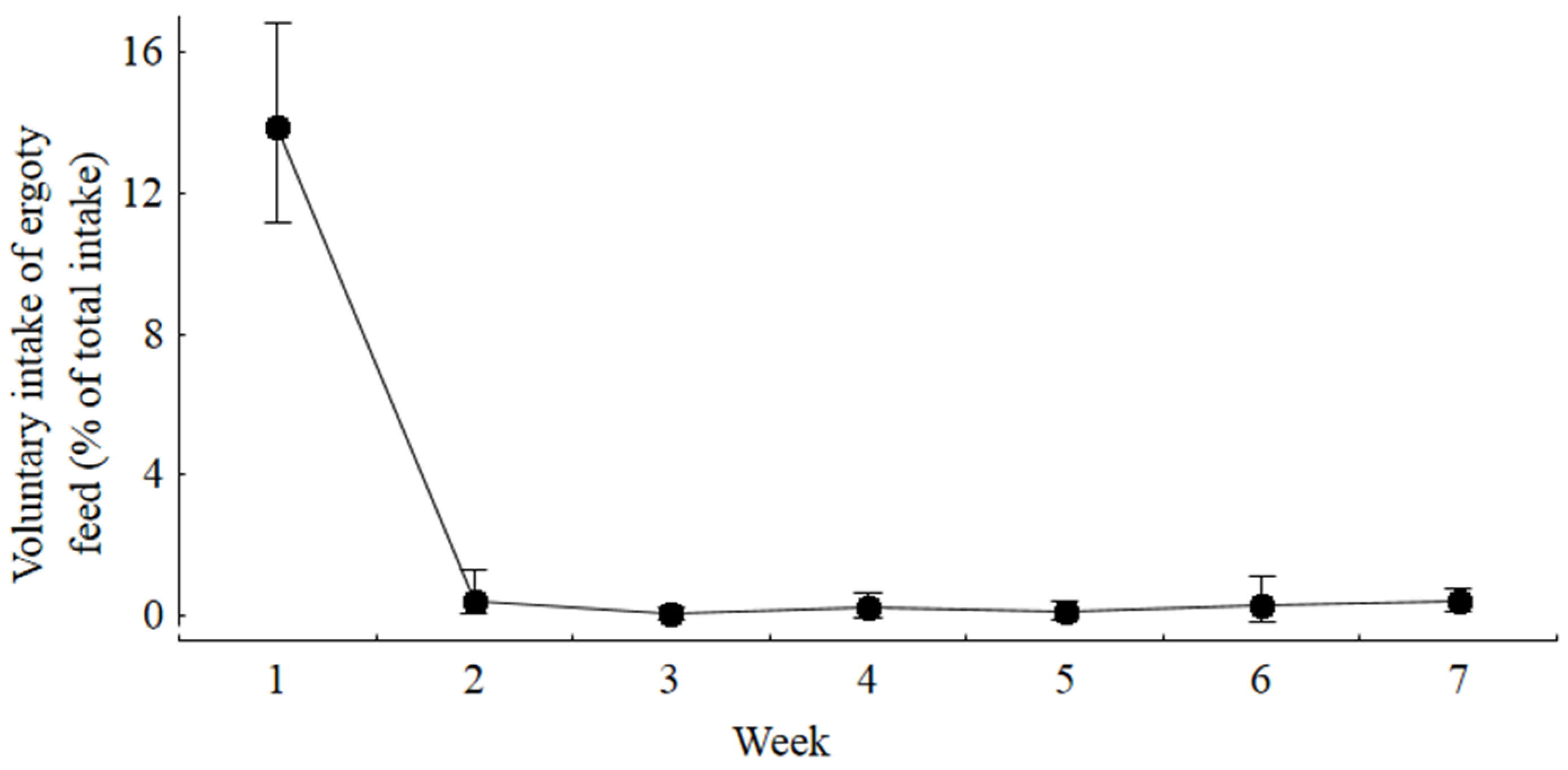
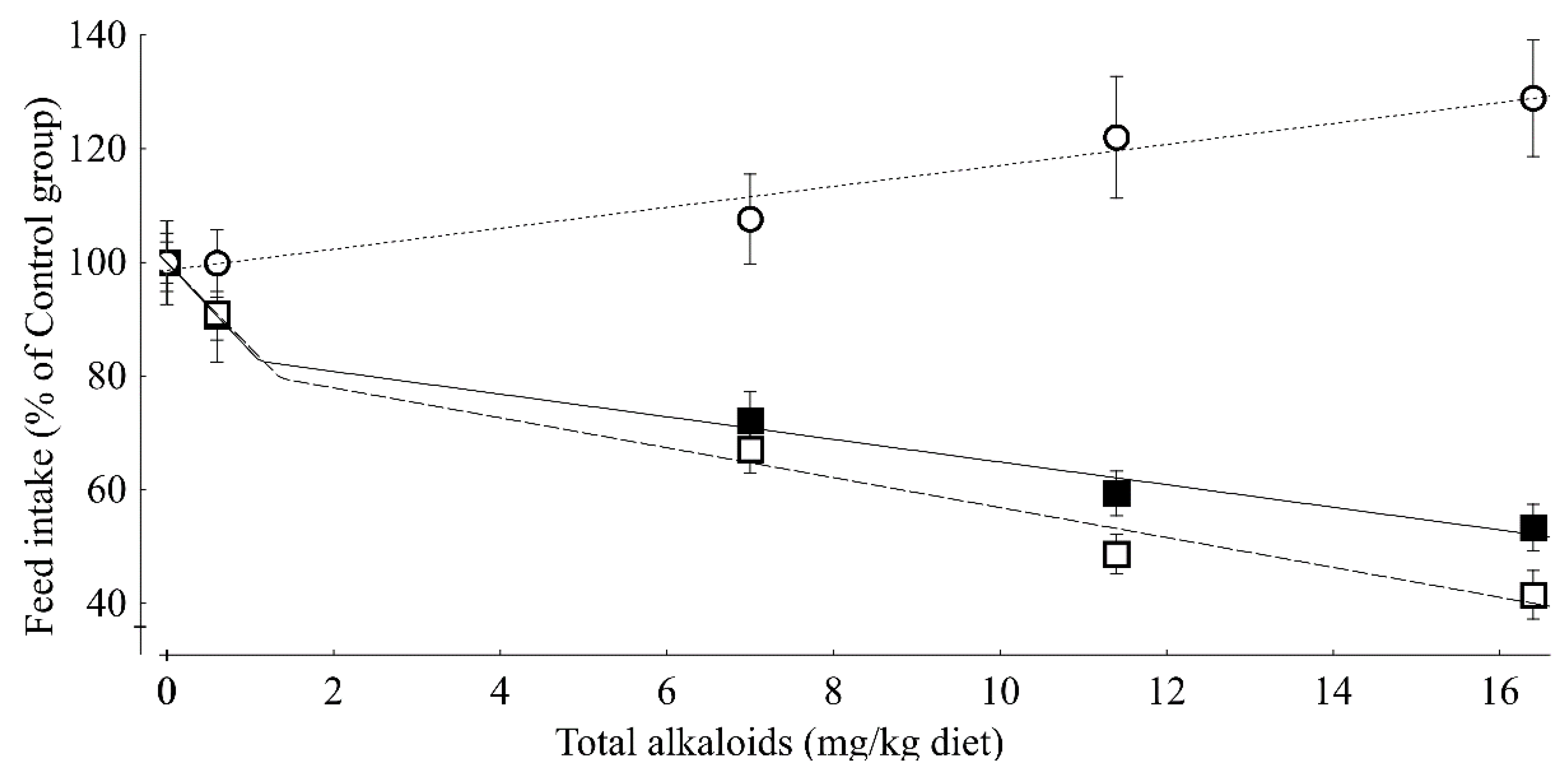
Our experiment demonstrated that ducks responded very sensitively to the presence of ergot alkaloids in feed with a significant decrease in voluntary feed intake even at the lowest TEA concentration of 0.6 mg/kg diet. Due to this pronounced feed intake-depressing effect, live weight gain was influenced in a similar manner. Although both feed intake and live weight gain responded in a similar direction, the resulting feed to gain ratio increased at the same time during week 1 of the experiment. Therefore, the nutrient and energy utilization was dose dependently compromised during the initial period of the experiment. In contrast, during week 2 of the experiment, the feed to gain ratio remained uninfluenced and was even decreased during the later periods of the experiment for the groups remaining in the experiment.
The ergot-associated decrease in feed intake might have been caused by several factors. Generally, common features of plant alkaloids are their bitter taste and the pharmacological activity. Plant alkaloids serve as a chemical defense against herbivory [
6] and it is assumed that the ecological role of ergot alkaloids is also to protect the fungi from consumption by vertebrate and invertebrate animals [
7]. Ducks and chickens, compared to pigeons, seem to be particularly sensitive to a bitter taste as indicated by a pronounced rejection of a solution of quinine hydrochloride, a model substance for bitterness, at low concentrations [
8]. Male geese (another waterfowl closely related to ducks), when compared to turkeys, Japanese quail and chickens, had a more pronounced response with a decrease in voluntary intake of quinine-containing diets in a dose-dependent manner [
9]. In discussing the feed intake depressing effects of ergot, the presence of further substances with potential anorectic or other toxic acting substances in ergot, such as ricinoleic acid, need to be considered. As ricinoleic acid increased with dietary ergot and consequently with TEA, it is impossible to assign individual or interactive effects conclusively. However, sub-acute toxicity studies with mice and rats fed graded levels of castor oil processed from the castor bean plant,
Ricinus communis L, which contains approximately 90% ricinoleic acid, revealed that dietary proportions of 10% (100g/kg) castor oil did not cause adverse effects [
10]. That ricinoleic acid might contribute only partly to the overall toxicity of ergot might be deduced from stepwise regressions of TEA and ricinoleic acid from ergot on the live weight gain of piglets. Although TEA alone could explain 82% of the variation in live weight gain, the additional inclusion of ricinoleic acid increased the variation explained to only 86% [
11]. Because of the low toxicity in rodents and the rather small contribution of ricinoleic acid to the overall toxicity of ergot, a rather low effect might be deduced for the present experiment where the highest dietary ricinoleic acid concentration amounted to 1.1 g/kg.
The chicken is generally believed to avoid toxin-containing diets that had previously caused illness, disturbances or discomfort [
12]. This general effect might be caused by metabolic signals due to ingesting the substance under question besides, or in addition to taste aversion. In the present experiment, we found a pronounced decrease in feed intake of
ad libitum fed ducks when no chance for avoiding the contaminated diets was offered, while choice fed ducks effectively learned to avoid the contaminated diet within one week in spite of weekly changes to the positions of the troughs containing the uncontaminated and the ergot-containing diet. Besides the bitter taste acting at the ingestive phase, metabolic alterations caused by the ergot alkaloid interactions with serotoninergic, dopaminergic and adrenergic receptors might also contribute to the post-ingestive modulation of voluntary feed intake. Ergot alkaloids are largely capable of permeating the blood-brain barrier [
13], which supports the view that decrease in feed intake in
ad libitum fed ducks and ergot-containing feed rejection in choice fed ducks might be mediated by central-nervous signals. Although we failed to detect ergot alkaloids in systemic blood, we confirmed the presence of ergonovine in bile, which indicates at least a portal absorption while a small fraction might have entered the systemic circulation and eventually the blood-brain barrier.
Recently, it was shown that colon and liver cell lines (HT-29, HepG2) are capable of intense hydroxylation at the peptide moiety while the toxico-dynamically relevant ergoline structure remained untouched [
14]. Thus, the peptide ergot alkaloids ergotamine/ergotaminine and ergocristine/ergocristinine were metabolized in this way while the lysergic acid amide derivatives ergonovine/ergonovinine were neither taken up by the cells nor metabolized [
14,
15]. The unaltered structure of ergonovine might explain, at least in part, why we could detect solely this ergot alkaloid in bile, as our HPLC-method barely detects the free forms of ergot alkaloids, while metabolized forms including lysergic acid escape detection. However, the fact that ergonovine/ergonovinine were not taken up either by intestinal or hepatic cell lines in these
in vitro experiments raises the question on the reasons for the presence of ergonovine in bile in our
in vivo study. It might be hypothesized that only ergonovine could be detected amongst the ergot alkaloids and solely in bile due to absorption of small amounts over a longer period of time in combination with its accumulation and its poor metabolism, leaving it detectable by our HPLC-method.
The liver not only plays a role in metabolizing and excretion of ergot alkaloids as discussed above, but has also been suggested as a primary target of portally delivered toxins.
Activities of enzymes in blood that are more or less indicative for hepatic lesions such as γ-glutamyl transferase (GGT), glutamate dehydrogenase (GLDH) and the less specific aspartate aminotransferase (ASAT) have been shown to respond either not at all (GLDH, GGT) or with an inconsistent and not always reproducible increase (ASAT) in piglets fed diets varying largely in the TEA between 3 and 21 mg/kg [
16,
17,
18,
19,
20], whereas in calves, fattening bulls and dairy cows no effects on these enzyme activities could be detected [
21,
22,
23]. Also to be considered in evaluating these effects are the differences in the pattern of individual alkaloids which are summed up by the TEA content, which was demonstrated to play a role for feed intake and live weight gain but was less important for liver lesions [
17,
19]. Broilers were shown to respond with an increase in GGT and ASAT activity when fed diets with TEA of 3 and 11 mg/kg diet [
20]. In our study, GGT activity in ducks increased significantly at a dietary TEA of 7 mg/kg diet, which is approximately 10-fold higher than the diet concentration found to be effective for decreasing feed intake, indicating that the liver responds less sensitively than feed intake in ducks, also. Interestingly, and in contrast to reports of ergot-associated decreases in blood albumin concentration in piglets and broiler chickens [
19,
20], ducks responded with an increase even at the lowest diet concentration of 0.6 mg TEA/kg. As albumin concentration in blood not only reflects the liver function with regard to protein synthesis function but might also be affected by hemo-concentration or -dilution, the contradictory results should not be overemphasized.
That the liver obviously responds to TEA at higher dietary concentrations than feed intake or live weight gain is further substantiated by the results of liver function tests. These tests are based on the hepatic cytochrome P450 (CYP1A2)-catalyzed conversion of stable isotope-labeled substrates to labeled CO
2, which can be measured in breath. These tests indicated LOAELs of 17 and 21 mg TEA/kg diet for male and female piglets, respectively [
18] while a literature compilation on the relationships between TEA content of the diet and performance revealed an incremental decrease of 0.9% and 1.24% per each 1 mg increase of TEA per kg diet when feed intake and live weight gain were used as response criteria [
16]. Based on the corresponding decreases of 15.7% and 15.0% for TEA concentrations ≤1.1 and 1.4 mg TEA/kg, and at 1.99% and 2.63% for TEA concentrations higher than these break-points, the ducks seem to respond more sensitively to dietary ergot alkaloids than piglets.
Whether the ergot-associated increase in liver weight relative to body weight is a direct consequence of the toxic action of ergot alkaloids or due to a retardation of muscle growth relative to the dynamics of weight development of inner organs as a reflection of the ontogenetic allometry cannot be answered conclusively as further parameters such as histopathological lesions were not recorded. Similarly, the ergot induced increase of the relative gizzard weight might be explained in this way although other digestive organs appeared to be unaffected.
The only conspicuous difference in white blood count was the ergot-related decrease in the proportions of monocytes, which might hint at an effect of ergot alkaloids in modulation of immune responses. Subcutaneous ergotamine tartrate doses equal or higher than 2 mg/kg body weight resulted in the secretion of pro-inflammatory cytokines by LPS stimulated murine splenocytes and macrophages, while differential white blood count remained unaffected [
24]. In steers, the LPS-induced acute phase response was attenuated by intravenous administration of ergotamine, which was supposed to result from the ergotamine-associated cortisol increase [
25]. Thus, the relevance and the nature of the ergot-associated decrease in the monocyte proportions observed in our experiment requires further experimental consideration. The general conclusion that poultry tolerates higher levels of ergot alkaloids than other non-ruminant livestock [
2] was drawn by deriving a NOAEL of 1.4 mg TEA/kg diet based on reviewing the literature published since the last EFSA-opinion on ergot alkaloids in 2005. However, mainly chickens were considered in the latest EFSA-opinion while no experiments on ducks were available at that time. As the lowest dose tested in our experiments caused adverse effects, we could not derive a NOAEL. Thus, the NOAEL for ducks is lower than 0.6 mg TEA/kg and consequently also markedly lower than that of 1.4 mg TEA/kg diet for chickens, as suggested by EFSA [
2].
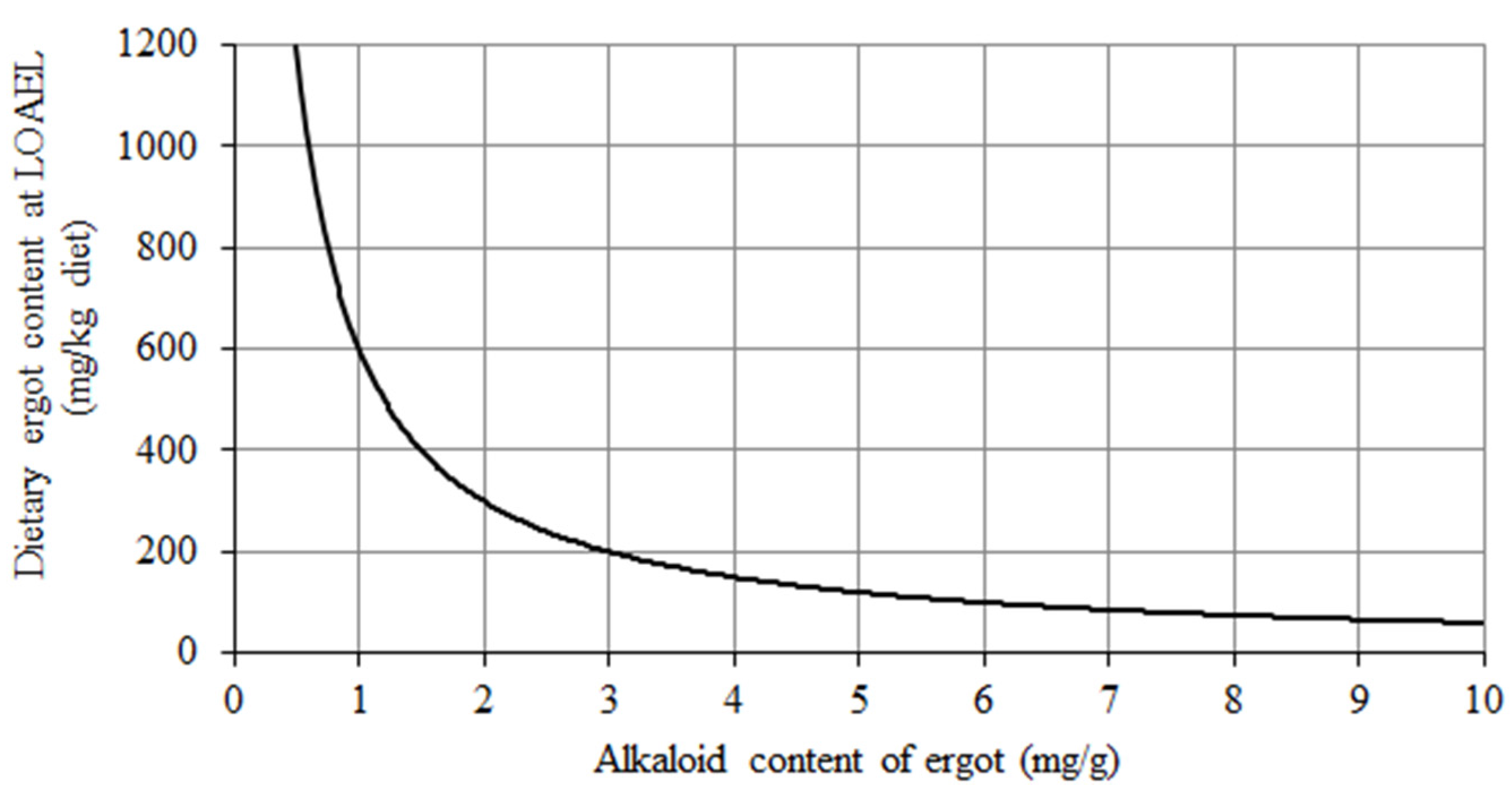
In view of the unreliability of the current feed safety regulations regarding the upper limit of 1000 mg ergot (
C. purpurea) per kg unground cereal grains (=0.1%), as specified by Directive 2002/32/EC, and the significant variation in TEA content of ergot as reviewed by EFSA [
2], a risk evaluation for the duck covering this variation and considering the LOAEL of 0.6 mg TEA/kg diet was performed to identify those dietary ergot levels where this LOAEL is reached for a particular TEA content of ergot (
Figure 3). As we failed to derive a NOAEL, the estimated dietary ergot contents indicate ergot levels where adverse effects already can be expected. For example, if the ergot alkaloid content of ergot reaches approximately 0.6 mg/g the permitted ergot content of 1000 mg/kg feed would also be exceeded. However, this is not a realistic scenario as the upper limit applies for pure (unground) cereal grains, which do not constitute the whole diet of a duck (see also
Table 2). Rather, the cereal content of a duck diet varies from a few percent up to more than 50%, which also needs to be considered when evaluating the risk of ergot contaminated cereal grains for this poultry species (
Figure 3). For example, the LOAEL of 0.6 mg TEA/kg diet would correspond to a dietary ergot content of 600 mg/kg diet when the diet would consist of 100% of cereal grains contaminated by ergot with a TEA content of 1 mg/g ergot (
Figure 3). If the same grain batch would be incorporated into the diet at a proportion of 50%, the TEA content of the ergot could approximate 2 mg/g. More generally expressed, the area to the right of the curve in
Figure 3 represents all combinations of TEA content of ergot and dietary ergot content, which would exceed the LOAEL. Again, as the curve was constructed based on LOAEL instead of the NOAEL, the combinations of dietary ergot content and TEA content of ergot, which would result in safe dietary TEA levels, are not known so far but are within the area left of the curve.
Figure 3.
Estimation of the ergot proportion in diets for ducks (mg/kg diet) where the LOAEL of 0.6 mg TEA/kg diet is reached in dependence on varying TEA contents of ergot (mg/g ergot).
Figure 3.
Estimation of the ergot proportion in diets for ducks (mg/kg diet) where the LOAEL of 0.6 mg TEA/kg diet is reached in dependence on varying TEA contents of ergot (mg/g ergot).



{kind=link}
{kind=link}
{kind=link}


