
Addition of Carbon to the Culture Medium Improves the Detection Efficiency of Aflatoxin Synthetic Fungi

Abstract
:1. Introduction
2. Results
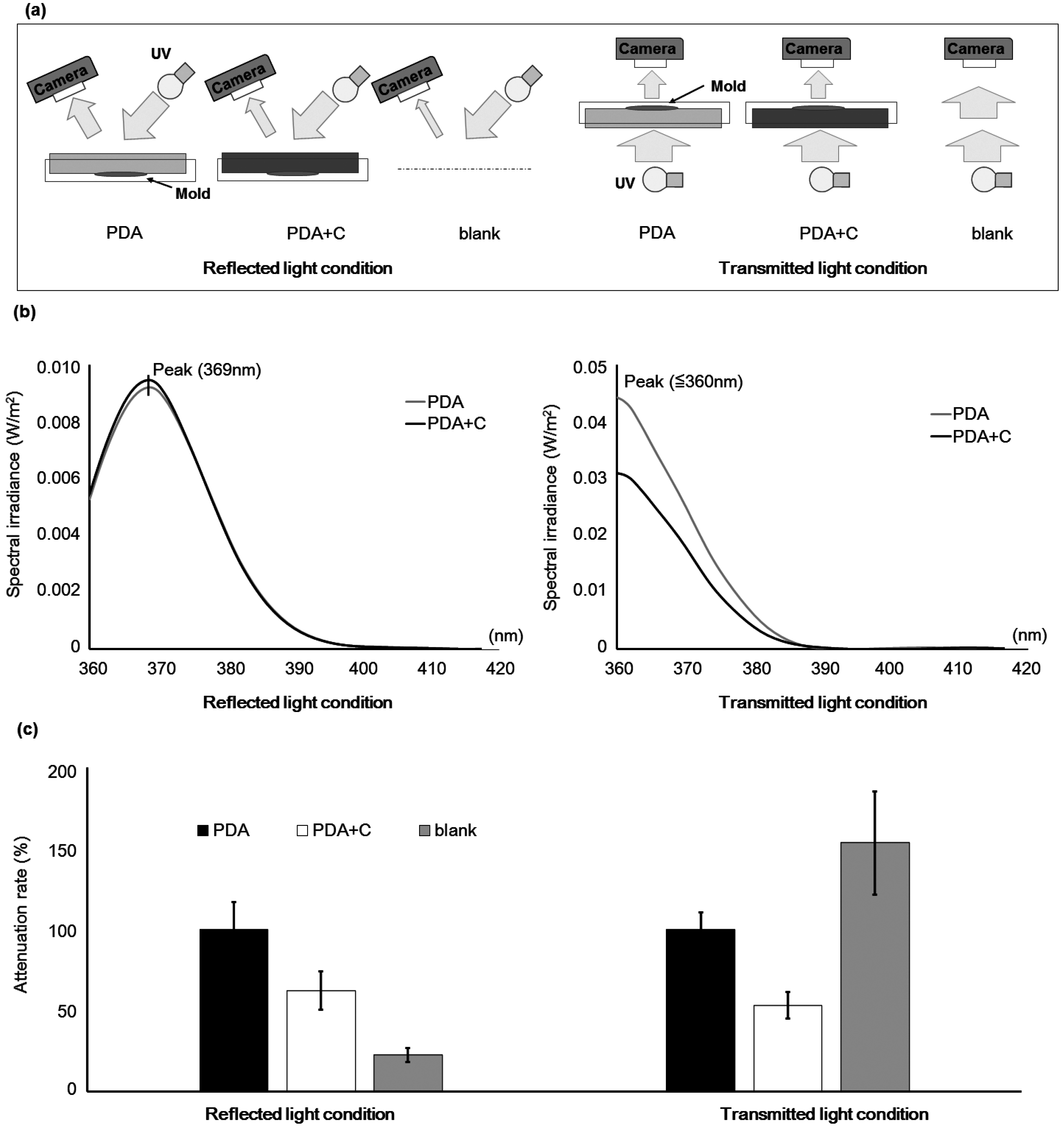
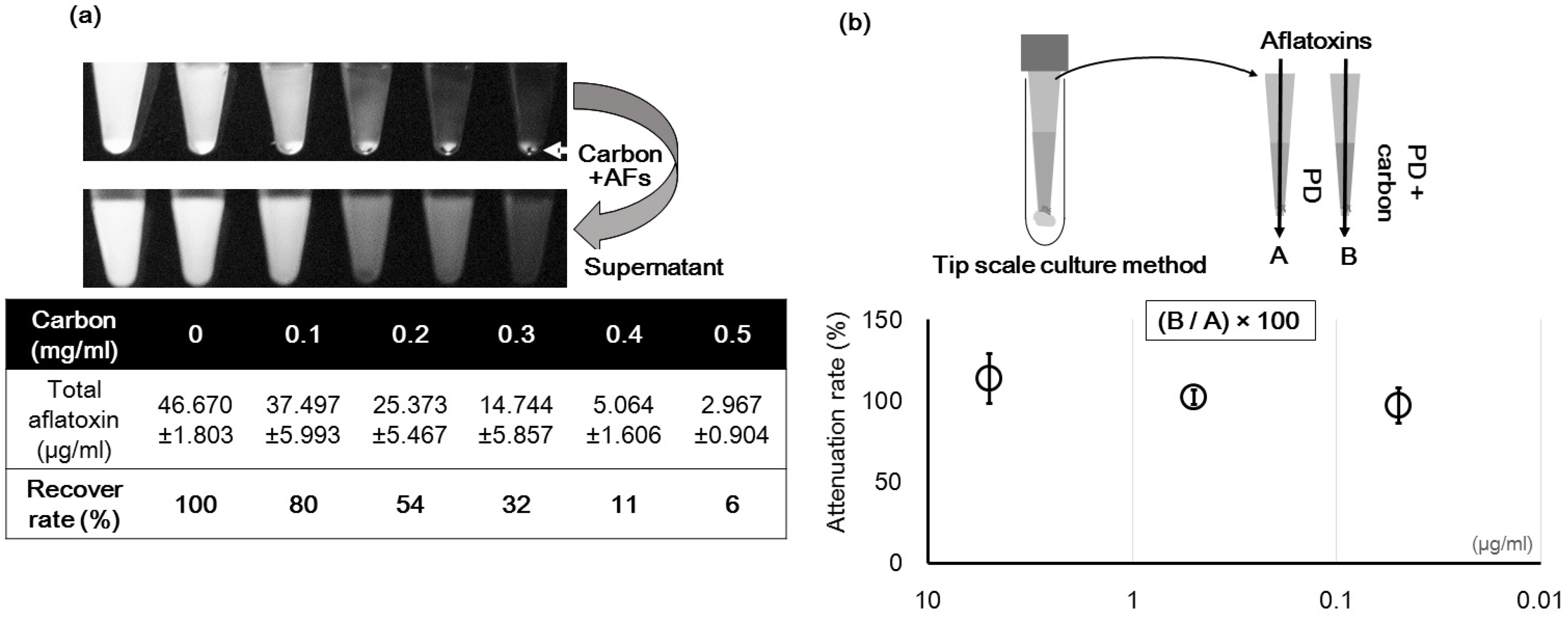
2.1. Changes in the Optical Characteristics of the Culture Medium with Addition of Carbon
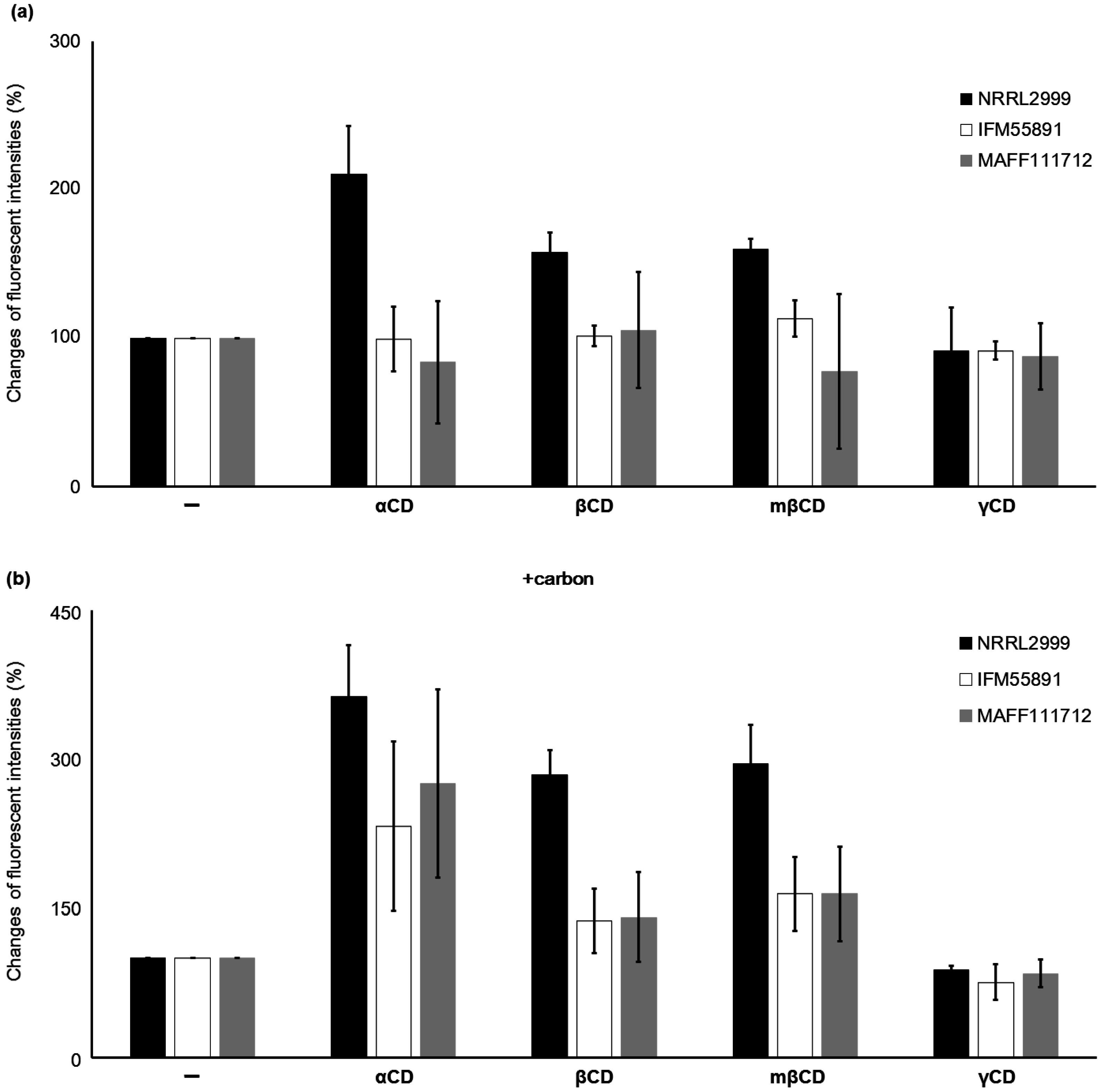
2.1.1. Changes in the Fluorescence with Different Culture Media
2.1.2. Effect of the Type of CD on the Fluorescence
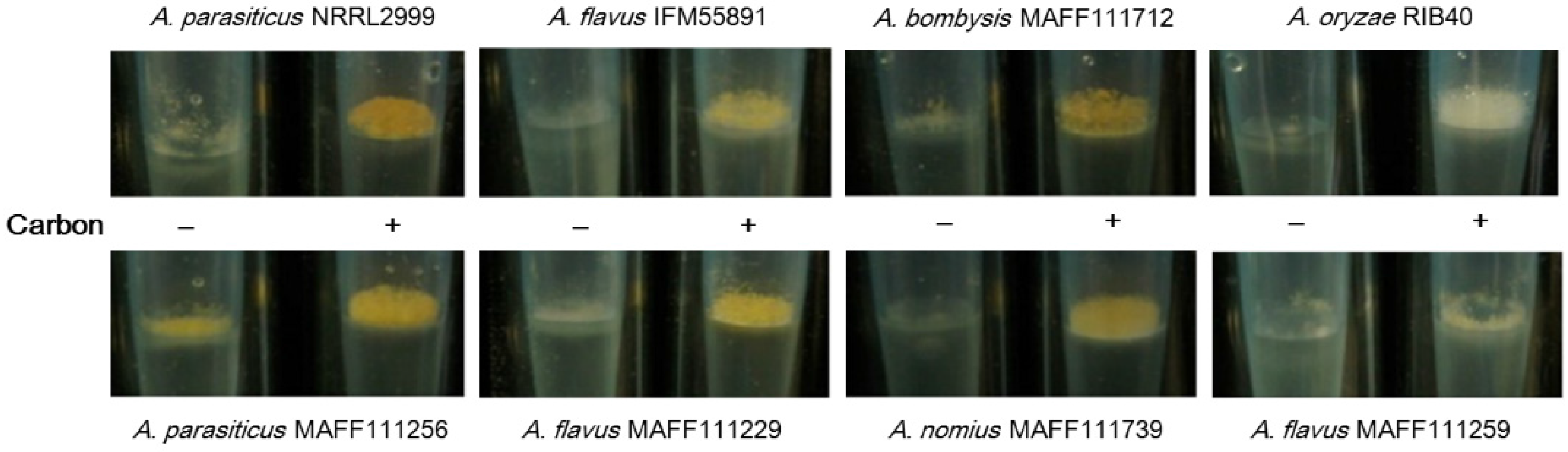
2.1.3. Correlation of the Color of Fluorescence with the Aspergillus Species and Strain
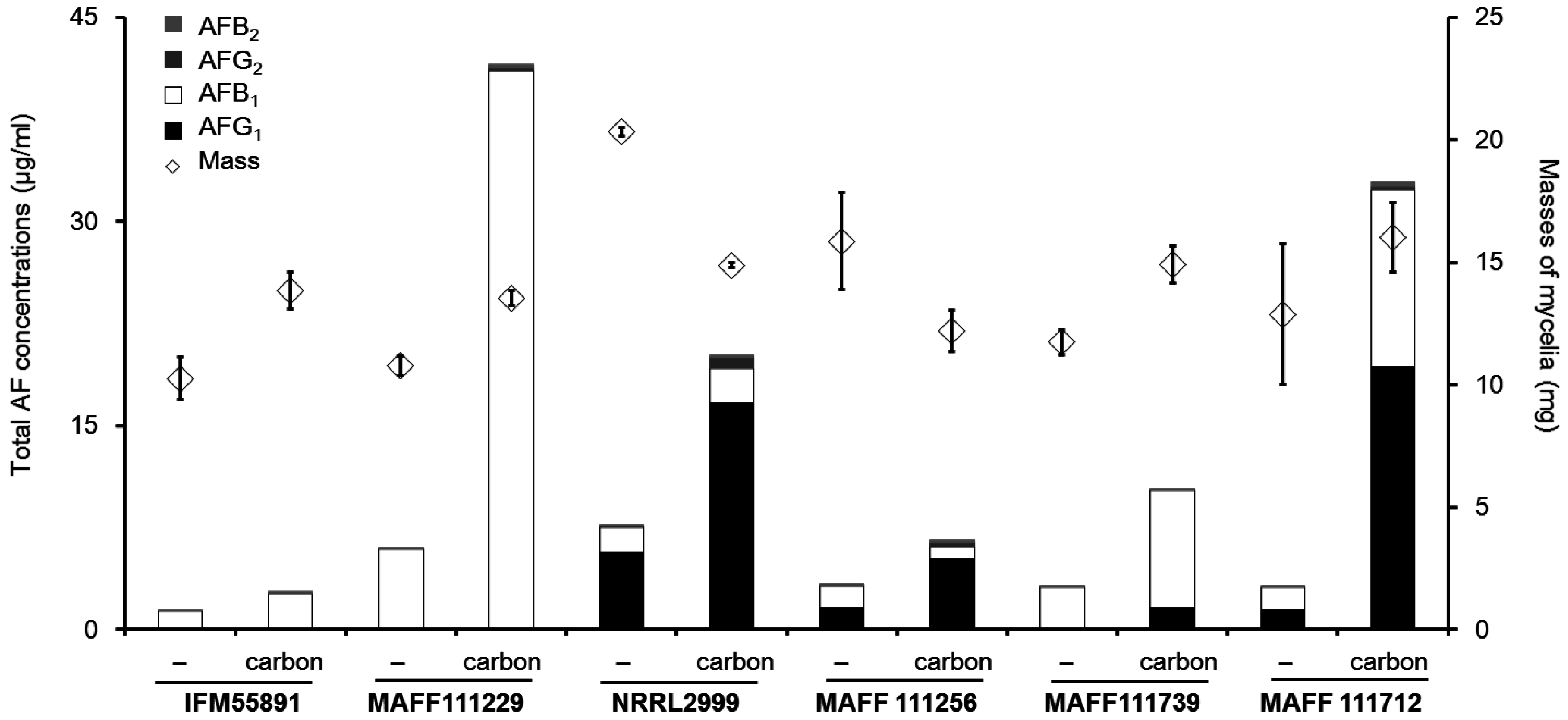
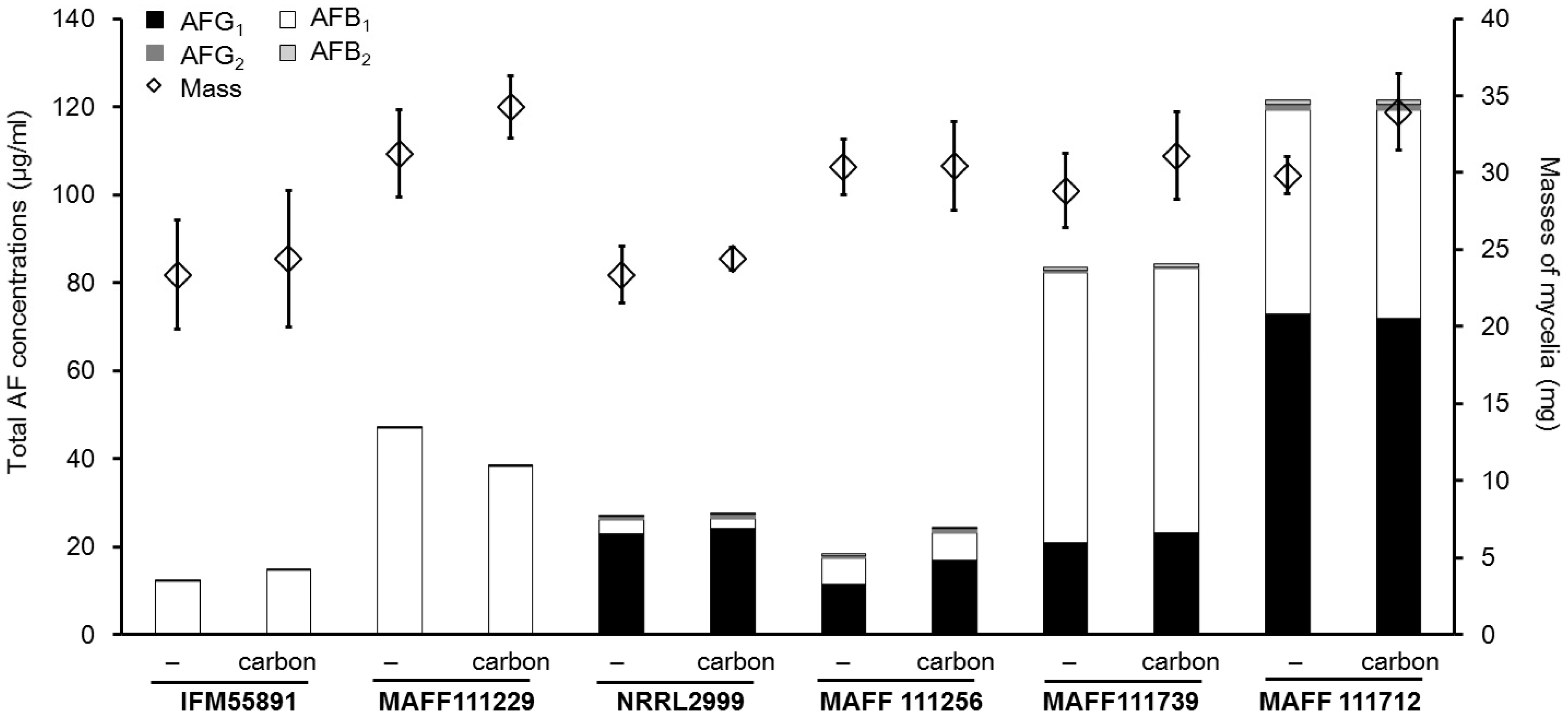
2.2. Effect of the Addition of Carbon to a Liquid Culture
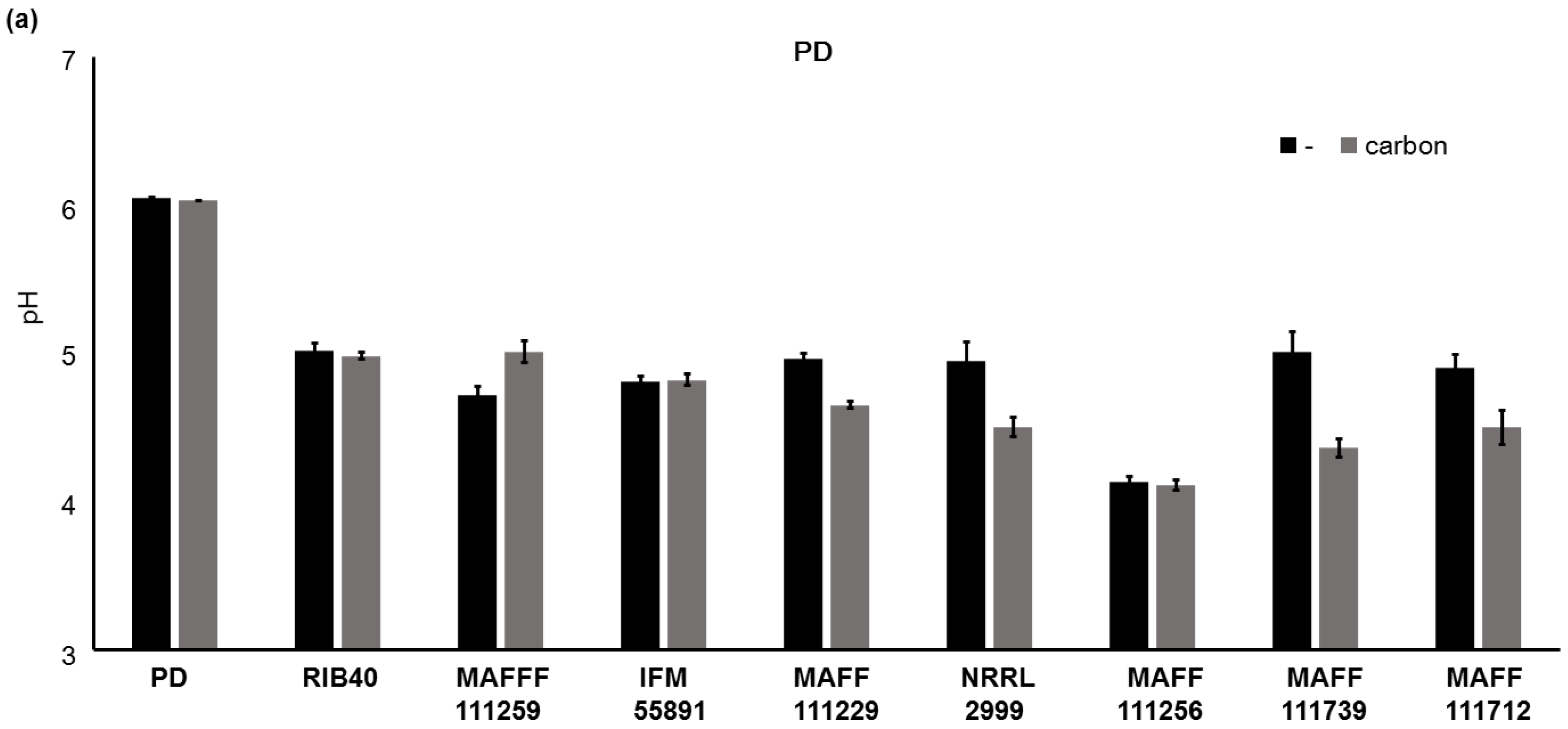
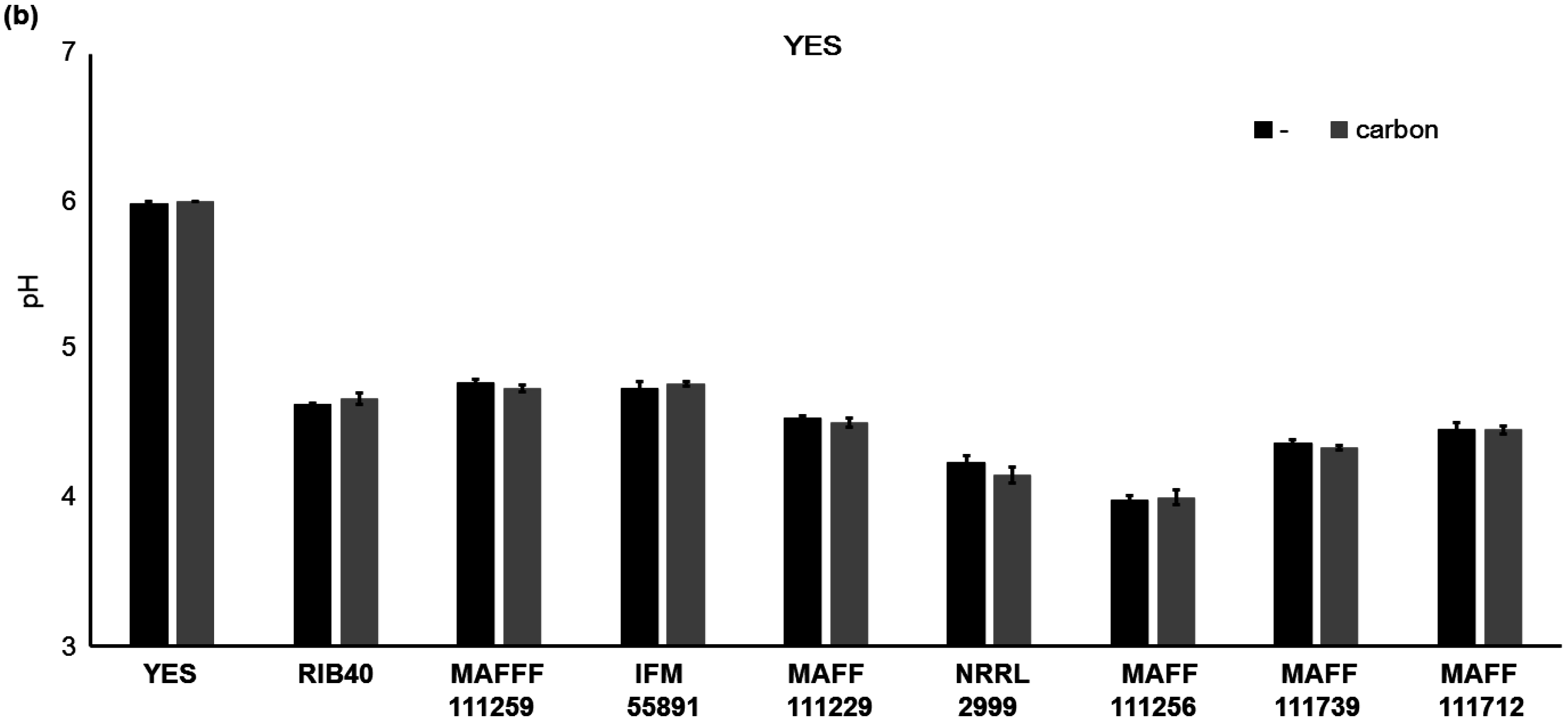
2.3. Factors not Related to the Changes in AF Concentrations
3. Discussion
3.1. Altering the Culture Medium to Enhance the Fluorescence Intensity
3.2. Effect of the Increased AF Volume Involved in Fungal Growth
4. Conclusions
5. Materials and Methods
5.1. Fungal Species
5.2. Culture Media Conditions, and Measurement of Fluorescence
5.3. Measurement of Light Attenuation under Reflected and Transmitted Light Conditions
5.4. Small-Scale Liquid Culture Conditions for AF Synthesis
5.5. High Performance Liquid Chromatography
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chang, P.K.; Horn, B.W.; Dorner, J.W. Sequence breakpoints in the aflatoxin biosynthesis gene cluster and flanking regions in nonaflatoxigenic Aspergillus flavus isolates. Fungal Genet. Biol. 2005, 42, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Ehrlich, K.C. What does genetic diversity of Aspergillus flavus tell us about Aspergillus oryzae? Int. J. Food Microbiol. 2010, 138, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, D.; Cary, J.W.; Ehrlich, K.; Yu, J.; Cleveland, T.E. Understanding the genetics of regulation of aflatoxin production and Aspergillus flavus development. Mycopathologia 2006, 162, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Ehrlich, K.C.; Beltz, S.B.; Harris-Coward, P.; Klich, M.A. Characterization of the Aspergillus ochraceoroseus aflatoxin/sterigmatocystin biosynthetic gene cluster. Mycologia 2009, 101, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C. Predicted roles of the uncharacterized clustered genes in aflatoxin biosynthesis. Toxins (Basel) 2009, 1, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins (Basel) 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Chihaya, N.; Hatabayashi, H.; Kito, M.; Hoshino, S.; Zeng, H.; Cai, J.; Nakajima, H. Production of M-/GM-group aflatoxins catalyzed by the OrdA enzyme in aflatoxin biosynthesis. Fungal Genet. Biol. 2012, 49, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.E. PCR detection of aflatoxin producing fungi and its limitations. Int. J. Food Microbiol. 2012, 156, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Abrar, M.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Randhawa, M.A.; Saeed, F.; Waqas, K. Aflatoxins: Biosynthesis, occurrence, toxicity, and remedies. Crit. Rev. Food Sci. Nutr. 2013, 53, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.; Venâncio, A.; Kozakiewicz, Z.; Lima, N. A polyphasic approach to the identification of aflatoxigenic and non-aflatoxigenic strains of Aspergillus Section Flavi isolated from Portuguese almonds. Int. J. Food Microbiol. 2009, 129, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.M.; Chung, S.H.; Chun, H.S. Multiplex PCR assay for the detection of aflatoxigenic and non-aflatoxigenic fungi in meju, a Korean fermented soybean food starter. Food Microbiol. 2011, 28, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.B.; Lee, M.; Kim, D.H.; Chung, S.H.; Shin, H.D.; Samson, R.A. The proportion of non-aflatoxigenic strains of the Aspergillus flavus/oryzae complex from meju by analyses of the aflatoxin biosynthetic genes. J. Microbiol. 2013, 51, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Bintvihok, A.; Treebonmuang, S.; Srisakwattana, K.; Nuanchun, W.; Patthanachai, K.; Usawang, S. A rapid and sensitive detection of aflatoxin-producing fungus using an optimized polymerase chain reaction (PCR). Toxicol. Res. 2016, 32, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.; Franco, C.; Cepeda, A.; Prognon, P.; Mahuzier, G. Liquid chromatographic study of the interaction between aflatoxins and β-cyclodextrin. Anal. Chim. Acta 1992, 269, 239–247. [Google Scholar] [CrossRef]
- Fente, C.A.; Ordaz, J.J.; Vázquez, B.I.; Franco, C.M.; Cepeda, A. New additive for culture media for rapid identification of aflatoxin-producing Aspergillus strains. Appl. Environ. Microbiol. 2001, 67, 4858–4862. [Google Scholar] [CrossRef] [PubMed]
- Jaimez Ordaz, J.; Fente, C.A.; Vázquez, B.I.; Franco, C.M.; Cepeda, A. Development of a method for direct visual determination of aflatoxin production by colonies of the Aspergillus flavus group. Int. J. Food Microbiol. 2003, 83, 219–225. [Google Scholar] [CrossRef]
- Hashemi, J.; Kram, G.A.; Alizadeh, N. Enhanced spectrofluorimetric determination of aflatoxin B1 in wheat by second-order standard addition method. Talanta 2008, 75, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Degola, F.; Dall’Asta, C.; Restivo, F.M. Development of a simple and high-throughput method for detecting aflatoxins production in culture media. Lett. Appl. Microbiol. 2012, 55, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Galicia, G.; Garduño-Juárez, R.; Gabriela Vargas, M. Effect of water molecules on the fluorescence enhancement of Aflatoxin B1 mediated by Aflatoxin B1: β-cyclodextrin complexes. A theoretical study. Photochem. Photobiol. Sci. 2007, 6, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, P.; Sharifnabi, B.; Bahar, M. Detection of Aflatoxin in Aspergillus species isolated from pistachio in Iran. J. Phytopathol. 2008, 156, 15–20. [Google Scholar] [CrossRef]
- Accinelli, C.; Abbas, H.K.; Zablotowicz, R.M.; Wilkinson, J.R. Aspergillus flavus aflatoxin occurrence and expression of aflatoxin biosynthesis genes in soil. Can. J. Microbiol. 2008, 54, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, M.; Cepeda, A.; Prognon, P.; Mahuzier, G.; Blais, J. Cyclodextrins as modifiers of the luminescence characteristics of aflatoxins. Anal. Chim. Acta 1991, 255, 343–350. [Google Scholar] [CrossRef]
- Rojas-Durán, T.; Sánchez-Barragán, I.; Costa-Fernández, J.M.; Sanz-Medel, A. Direct and rapid discrimination of aflatoxigenic strains based on fibre-optic room temperature phosphorescence detection. Analyst 2007, 132, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, D.; Zainal Abidin, M.; Tan, Y.; Kamaruzaman, S. Evaluation of the detection techniques of toxigenic Aspergillus isolates. Afr. J. Biotechnol. 2010, 9, 7654–7659. [Google Scholar]
- Assante, G.; Camarda, L.; Locci, R.; Merlini, L.; Nasini, G.; Papadopoulos, E. Isolation and structure of red pigments from Aspergillus flavus and related species, grown on a differential medium. J. Agric. Food Chem. 1981, 29, 787–789. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D.; Glenn, D.R. An improved medium for the detection of Aspergillus flavus and A. parasiticus. J. Appl. Bacteriol. 1983, 54, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Dianese, J. A coconut-agar medium for rapid detection of aflatoxin production by Aspergillus spp. Phytopathology 1976, 66, 1466–1469. [Google Scholar] [CrossRef]
- Davis, N.D.; Iyer, S.K.; Diener, U.L. Improved method of screening for aflatoxin with a coconut agar medium. Appl. Environ. Microbiol. 1987, 53, 1593–1595. [Google Scholar] [PubMed]
- Cucullu, A.F.; Lee, L.S.; Pons, W.A.; Stanley, J.B. Ammoniation of aflatoxin B1: Isolation and characterization of a product with molecular weight 206. J. Agric. Food Chem. 1976, 24, 408–410. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Hatabayashi, H.; Ikehata, A.; Zheng, Y.; Kushiro, M. Development of the dichlorvos-ammonia (DV-AM) method for the visual detection of aflatoxigenic fungi. Appl. Microbiol. Biotechnol. 2015, 99, 10681–10694. [Google Scholar] [CrossRef] [PubMed]
- Diaz, D.E.; Hagler, W.M.; Hopkins, B.A.; Whitlow, L.W. Aflatoxin binders I: In vitro binding assay for aflatoxin B1 by several potential sequestering agents. Mycopathologia 2002, 156, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Khadem, A.; Sharifi, D.; Barati, M.; Borji, M. Evaluation of the effectiveness of yeast, zeolite and active charcoal as aflatoxin absorbents in broiler diets. Glob. Vet. 2012, 8, 426–432. [Google Scholar]
- Jiang, Y.H.; Wang, P.; Yang, H.J.; Chen, Y. The efficacy of bamboo charcoal in comparison with smectite to reduce the detrimental effect of aflatoxin B1 on in vitro rumen fermentation of a hay-rich feed mixture. Toxins (Basel) 2014, 6, 2008–2023. [Google Scholar] [CrossRef] [PubMed]
- Yabe, K.; Yan, P.S.; Song, Y.; Ichinomiya, M.; Nakagawa, H.; Shima, Y.; Ando, Y.; Sakuno, E.; Nakajima, H. Isolation of microorganisms and substances inhibitory to aflatoxin production. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2008, 25, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Jindal, N.; Mahipal, S.K.; Mahajan, N.K. Toxicity of aflatoxin B1 in broiler chicks and its reduction by activated charcoal. Res. Vet. Sci. 1994, 56, 37–40. [Google Scholar] [CrossRef]
- National Institutes of Health. Image Processing and Analysis in Java. Available online: https://imagej.nih.gov/ij/ (accessed on 6 August 2015).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AF Type | AFB1 | AFB2 | AFG1 | AFG2 | Total |
|---|---|---|---|---|---|
| AF concentrations (μg/mg) ± S.D. 1 | |||||
| F1 | 0.135 ± 0.048 | 0.002 ± 0.001 | - | - | 0.137 ± 0.048 |
| F1 + C 2 | 0.194 ± 0.072 | 0.008 ± 0.003 | - | - | 0.202 ± 0.075 |
| F2 | 0.552 ± 0.133 | 0.001 ± 0.001 | - | - | 0.553 ± 0.134 |
| F2 + C | 3.029 ± 0.468 | 0.015 ± 0.003 | 0.008 ± 0.003 | 0.020 ± 0.005 | 3.072 ± 0.467 |
| P1 | 0.090 ± 0.012 | 0.005 ± 0.000 | 0.279 ± 0.057 | 0.006 ± 0.001 | 0.379 ± 0.069 |
| P1 + C | 0.176 ± 0.024 | 0.014 ± 0.002 | 1.117 ± 0.187 | 0.050 ± 0.007 | 1.357 ± 0.213 |
| P2 | 0.104 ± 0.009 | 0.008 ± 0.001 | 0.101 ± 0.009 | - | 0.212 ± 0.018 |
| P2 + C | 0.076 ± 0.016 | 0.009 ± 0.002 | 0.426 ± 0.050 | 0.025 ± 0.004 | 0.537 ± 0.071 |
| N | 0.263 ± 0.042 | 0.001 ± 0.001 | 0.007 ± 0.003 | - | 0.271 ± 0.045 |
| N + C | 0.578 ± 0.093 | 0.002 ± 0.000 | 0.108 ± 0.020 | - | 0.688 ± 0.108 |
| B | 0.130 ± 0.022 | 0.002 ± 0.001 | 0.110 ± 0.018 | - | 0.243 ± 0.040 |
| B + C | 0.816 ± 0.069 | 0.023 ± 0.002 | 1.203 ± 0.099 | 0.013 ± 0.002 | 2.054 ± 0.168 |
| Changes of AF concentrations caused by carbon supply (%) | |||||
| F1 + C | 144 | 409 | - | - | 148 |
| F2 + C | 548 | 2422 | - | - | 555 |
| P1 + C | 195 | 286 | 401 | 891 | 358 |
| P2 + C | 74 | 121 | 424 | 5342 | 253 |
| N + C | 220 | 276 | 1501 | - | 254 |
| B + C | 627 | 931 | 1090 | - | 845 |
| AF type | AFB1 | AFB2 | AFG1 | AFG2 | Total |
|---|---|---|---|---|---|
| AF concentrations (μg/mg) ± S.D. 1 | |||||
| F1 | 0.522 ± 0.090 | 0.006 ± 0.003 | - | - | 0.528 ± 0.092 |
| F1 + C 2 | 0.619 ± 0.146 | 0.011 ± 0.005 | - | - | 0.629 ± 0.150 |
| F2 | 1.503 ± 0.098 | 0.005 ± 0.000 | 0.005 ± 0.001 | 0.002 ± 0.000 | 1.516 ± 0.098 |
| F2 + C | 1.113 ± 0.094 | 0.004 ± 0.000 | 0.006 ± 0.002 | 0.003 ± 0.000 | 1.126 ± 0.096 |
| P1 | 0.137 ± 0.013 | 0.002 ± 0.001 | 0.981 ± 0.050 | 0.033 ± 0.004 | 1.153 ± 0.065 |
| P1 + C | 0.100 ± 0.010 | 0.001 ± 0.001 | 0.985 ± 0.057 | 0.039 ± 0.003 | 1.125 ± 0.070 |
| P2 | 0.200 ± 0.043 | 0.010 ± 0.002 | 0.377 ± 0.101 | 0.018 ± 0.005 | 0.606 ± 0.149 |
| P2 + C | 0.206 ± 0.049 | 0.011 ± 0.003 | 0.557 ± 0.104 | 0.032 ± 0.009 | 0.806 ± 0.158 |
| N | 2.141 ± 0.182 | 0.029 ± 0.001 | 0.730 ± 0.059 | 0.012 ± 0.002 | 2.912 ± 0.238 |
| N + C | 1.934 ± 0.117 | 0.025 ± 0.009 | 0.747 ± 0.104 | 0.011 ± 0.001 | 2.717 ± 0.165 |
| B | 1.565 ± 0.214 | 0.036 ± 0.006 | 2.444 ± 0.257 | 0.039 ± 0.005 | 4.085 ± 0.478 |
| B + C | 1.405 ± 0.272 | 0.033 ± 0.007 | 2.118 ± 0.236 | 0.035 ± 0.006 | 3.592 ± 0.516 |
| Changes of AF concentrations caused by carbon supply (%) | |||||
| F1 + C | 118.5 | 174.2 | - | - | 119.1 |
| F2 + C | 74.1 | 75.7 | 117.6 | 139.0 | 74.3 |
| P1 + C | 73.1 | 45.3 | 100.4 | 117.9 | 97.6 |
| P2 + C | 102.7 | 111.5 | 147.7 | 177.6 | 133.1 |
| N + C | 90.3 | 87.4 | 102.3 | 85.8 | 93.3 |
| B + C | 89.8 | 89.9 | 86.7 | 90.8 | 87.9 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Iwahashi, Y. Addition of Carbon to the Culture Medium Improves the Detection Efficiency of Aflatoxin Synthetic Fungi. Toxins 2016, 8, 338. https://doi.org/10.3390/toxins8110338
Suzuki T, Iwahashi Y. Addition of Carbon to the Culture Medium Improves the Detection Efficiency of Aflatoxin Synthetic Fungi. Toxins. 2016; 8(11):338. https://doi.org/10.3390/toxins8110338
Chicago/Turabian StyleSuzuki, Tadahiro, and Yumiko Iwahashi. 2016. "Addition of Carbon to the Culture Medium Improves the Detection Efficiency of Aflatoxin Synthetic Fungi" Toxins 8, no. 11: 338. https://doi.org/10.3390/toxins8110338
APA StyleSuzuki, T., & Iwahashi, Y. (2016). Addition of Carbon to the Culture Medium Improves the Detection Efficiency of Aflatoxin Synthetic Fungi. Toxins, 8(11), 338. https://doi.org/10.3390/toxins8110338