
An Overview of Helicobacter pylori VacA Toxin Biology

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Features of vacA and Related Genes in Helicobacter Species
3. VacA Transcription, Regulation, and Secretion
4. Membrane Channel Formation by VacA
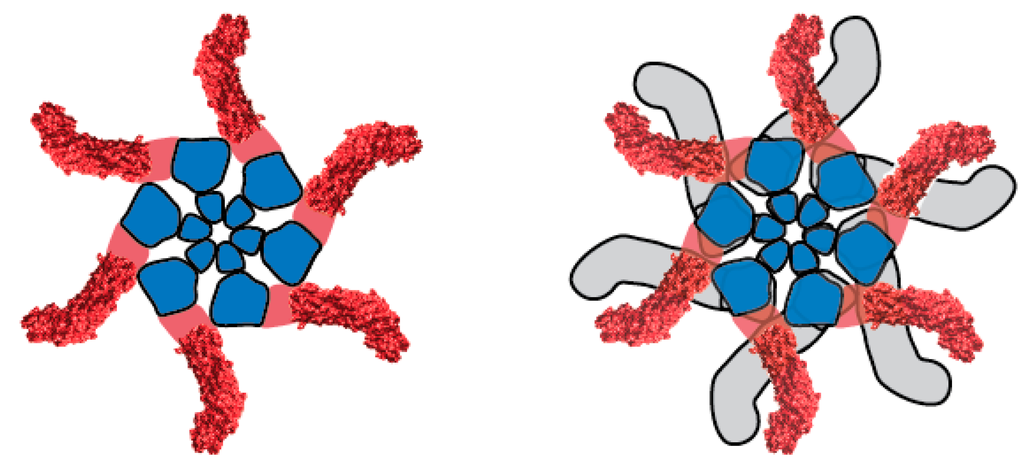
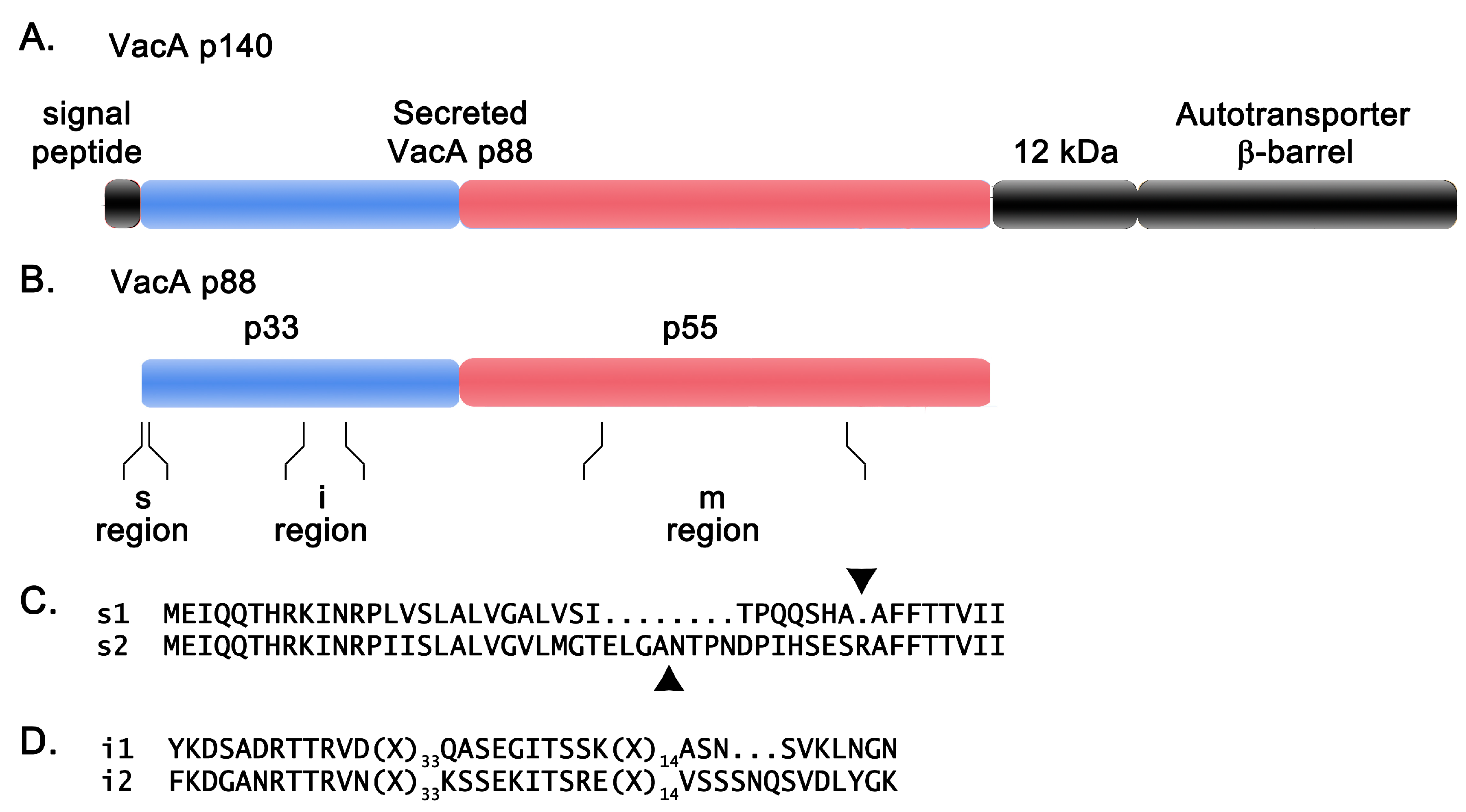
5. VacA Structure
6. VacA Diversity among H. pylori Strains
7. VacA Activities in Cell Culture Systems
7.1. Effects on Epithelial Cells
7.1.1. Endosomal Alterations
7.1.2. Autophagy
7.1.3. Mitochondrial Alterations
7.1.4. Epithelial Barrier Alterations
7.1.5. Altered Cell Signaling
7.1.6. Cell Death
7.2. Effects on Immune Cells and Parietal Cells
7.2.1. Effects on Immune Cells
7.2.2. Effects on Parietal Cells
8. Binding, Internalization, and Intracellular Trafficking of VacA
8.1. Cell Surface Binding and Receptors
8.2. Pore Formation at the Cell Surface
8.3. VacA Internalization and Intracellular Trafficking
9. Activity of VacA in Animal Models
10. Comparisons of VacA Activities in Vitro and in Vivo
11. Summary
Acknowledgments
Conflicts of Interest
References
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Leunk, R.D.; Johnson, P.T.; David, B.C.; Kraft, W.G.; Morgan, D.R. Cytotoxic activity in broth-culture filtrates of Campylobacter pylori. J. Med. Microbiol. 1988, 26, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blaser, M.J. Purification and characterization of the vacuolating toxin from Helicobacter pylori. J. Biol. Chem. 1992, 267, 10570–10575. [Google Scholar] [PubMed]
- Cover, T.L.; Tummuru, M.K.R.; Cao, P.; Thompson, S.A.; Blaser, M.J. Divergence of genetic sequences for the vacuolating cytotoxin among Helicobacter pylori strains. J. Biol. Chem. 1994, 269, 10566–10573. [Google Scholar] [PubMed]
- Schmitt, W.; Haas, R. Genetic analysis of the Helicobacter pylori vacuolating cytotoxin: Structural similarities with the IgA protease type of exported protein. Mol. Microbiol. 1994, 12, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Telford, J.L.; Ghiara, P.; Dell’Orco, M.; Comanducci, M.; Burroni, D.; Bugnoli, M.; Tecce, M.F.; Censini, S.; Covacci, A.; Xiang, Z.; et al. Gene structure of the Helicobacter pylori cytotoxin and evidence of its key role in gastric disease. J. Exp. Med. 1994, 179, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Phadnis, S.H.; Ilver, D.; Janzon, L.; Normark, S.; Westblom, T.U. Pathological significance and molecular characterization of the vacuolating toxin gene of Helicobacter pylori. Infect. Immun. 1994, 62, 1557–1565. [Google Scholar] [PubMed]
- Kersulyte, D.; Rossi, M.; Berg, D.E. Sequence divergence and conservation in genomes of Helicobacter cetorum strains from a dolphin and a whale. PLoS ONE 2013, 8, e83177. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.G.; Feng, Y.; Xu, S.; Taylor, N.S.; Kinsel, M.; Dewhirst, F.E.; Paster, B.J.; Greenwell, M.; Levine, G.; Rogers, A.; et al. Helicobacter cetorum sp. nov., a urease-positive helicobacter species isolated from dolphins and whales. J. Clin. Microbiol. 2002, 40, 4536–4543. [Google Scholar] [CrossRef] [PubMed]
- Goldman, C.G.; Loureiro, J.D.; Matteo, M.J.; Catalano, M.; Gonzalez, A.B.; Heredia, S.R.; Zubillaga, M.B.; Solnick, J.V.; Cremaschi, G.A. Helicobacter spp. From gastric biopsies of stranded south american fur seals (Arctocephalus australis). Res. Vet. Sci. 2009, 86, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Dailidiene, D.; Dailide, G.; Ogura, K.; Zhang, M.; Mukhopadhyay, A.K.; Eaton, K.A.; Cattoli, G.; Kusters, J.G.; Berg, D.E. Helicobacter acinonychis: Genetic and rodent infection studies of a Helicobacter pylori-like gastric pathogen of cheetahs and other big cats. J. Bacteriol. 2004, 186, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Eppinger, M.; Baar, C.; Linz, B.; Raddatz, G.; Lanz, C.; Keller, H.; Morelli, G.; Gressmann, H.; Achtman, M.; Schuster, S.C. Who ate whom? Adaptive helicobacter genomic changes that accompanied a host jump from early humans to large felines. PLoS Genet. 2006, 2, e120. [Google Scholar] [CrossRef]
- Gangwer, K.A.; Shaffer, C.L.; Suerbaum, S.; Lacy, D.B.; Cover, T.L.; Bordenstein, S.R. Molecular evolution of the Helicobacter pylori vacuolating toxin gene vacA. J. Bacteriol. 2010, 192, 6126–6135. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.; Buhrdorf, R.; Gerland, E.; Haas, R. Outer membrane targeting of passenger proteins by the vacuolating cytotoxin autotransporter of Helicobacter pylori. Infect. Immun. 2001, 69, 6769–6775. [Google Scholar] [CrossRef] [PubMed]
- Sause, W.E.; Castillo, A.R.; Ottemann, K.M. The Helicobacter pylori autotransporter ImaA (HP0289) modulates the immune response and contributes to host colonization. Infect. Immun. 2012, 80, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Radin, J.N.; Gaddy, J.A.; Gonzalez-Rivera, C.; Loh, J.T.; Algood, H.M.; Cover, T.L. Flagellar localization of a Helicobacter pylori autotransporter protein. MBio 2013, 4, e00613-12. [Google Scholar] [CrossRef] [PubMed]
- Kavermann, H.; Burns, B.P.; Angermuller, K.; Odenbreit, S.; Fischer, W.; Melchers, K.; Haas, R. Identification and characterization of Helicobacter pylori genes essential for gastric colonization. J. Exp. Med. 2003, 197, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, D.N.; Shepherd, B.; Kraemer, P.; Hall, M.K.; Sycuro, L.K.; Pinto-Santini, D.M.; Salama, N.R. Identification of Helicobacter pylori genes that contribute to stomach colonization. Infect. Immun. 2007, 75, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Castillo, A.R.; Woodruff, A.J.; Connolly, L.E.; Sause, W.E.; Ottemann, K.M. Recombination-based in vivo expression technology identifies Helicobacter pylori genes important for host colonization. Infect. Immun. 2008, 76, 5632–5644. [Google Scholar] [CrossRef] [PubMed]
- Voss, B.J.; Gaddy, J.A.; McDonald, W.H.; Cover, T.L. Analysis of surface-exposed outer membrane proteins in Helicobacter pylori. J. Bacteriol. 2014, 196, 2455–2471. [Google Scholar] [CrossRef] [PubMed]
- Albert, T.J.; Dailidiene, D.; Dailide, G.; Norton, J.E.; Kalia, A.; Richmond, T.A.; Molla, M.; Singh, J.; Green, R.D.; Berg, D.E. Mutation discovery in bacterial genomes: Metronidazole resistance in Helicobacter pylori. Nat. Methods 2005, 2, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, M.H.; Cover, T.L. Mutational analysis of the vacA promoter provides insight into gene transcription in Helicobacter pylori. J. Bacteriol. 1999, 181, 2261–2266. [Google Scholar] [PubMed]
- Amilon, K.R.; Letley, D.P.; Winter, J.A.; Robinson, K.; Atherton, J.C. Expression of the Helicobacter pylori virulence factor vacuolating cytotoxin A (vacA) is influenced by a potential stem-loop structure in the 5′ untranslated region of the transcript. Mol. Microbiol. 2015, 98, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, M.H.; Cover, T.L. Intercellular communication in Helicobacter pylori: LuxS is essential for the production of an extracellular signaling molecule. Infect. Immun. 2000, 68, 3193–3199. [Google Scholar] [CrossRef] [PubMed]
- Snider, C.A.; Voss, B.J.; McDonald, W.H.; Cover, T.L. Growth phase-dependent composition of the Helicobacter pylori exoproteome. J. Proteom. 2016, 130, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Merrell, D.S.; Thompson, L.J.; Kim, C.C.; Mitchell, H.; Tompkins, L.S.; Lee, A.; Falkow, S. Growth phase-dependent response of Helicobacter pylori to iron starvation. Infect. Immun. 2003, 71, 6510–6525. [Google Scholar] [CrossRef] [PubMed]
- Van Amsterdam, K.; van Vliet, A.H.; Kusters, J.G.; Feller, M.; Dankert, J.; van der Ende, A. Induced Helicobacter pylori vacuolating cytotoxin VacA expression after initial colonisation of human gastric epithelial cells. FEMS Immunol. Med. Microbiol. 2003, 39, 251–256. [Google Scholar] [CrossRef]
- Gancz, H.; Jones, K.R.; Merrell, D.S. Sodium chloride affects Helicobacter pylori growth and gene expression. J. Bacteriol. 2008, 190, 4100–4105. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.Q.; Caprioli, R.M.; Cover, T.L. Carboxy-terminal proteolytic processing of Helicobacter pylori vacuolating toxin. Infect. Immun. 2001, 69, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Bumann, D.; Aksu, S.; Wendland, M.; Janek, K.; Zimny-Arndt, U.; Sabarth, N.; Meyer, T.F.; Jungblut, P.R. Proteome analysis of secreted proteins of the gastric pathogen Helicobacter pylori. Infect. Immun. 2002, 70, 3396–3403. [Google Scholar] [CrossRef] [PubMed]
- Ilver, D.; Barone, S.; Mercati, D.; Lupetti, P.; Telford, J.L. Helicobacter pylori toxin VacA is transferred to host cells via a novel contact-dependent mechanism. Cell. Microbiol. 2004, 6, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Czajkowsky, D.M.; Iwamoto, H.; Cover, T.L.; Shao, Z. The vacuolating toxin from Helicobacter pylori forms hexameric pores in lipid bilayers at low pH. Proc. Natl. Acad. Sci. USA 1999, 96, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- Tombola, F.; Carlesso, C.; Szabo, I.; de Bernard, M.; Reyrat, J.M.; Telford, J.L.; Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M. Helicobacter pylori vacuolating toxin forms anion-selective channels in planar lipid bilayers: Possible implications for the mechanism of cellular vacuolation. Biophys. J. 1999, 76, 1401–1409. [Google Scholar] [CrossRef]
- Iwamoto, H.; Czajkowsky, D.M.; Cover, T.L.; Szabo, G.; Shao, Z. VacA from Helicobacter pylori: A hexameric chloride channel. FEBS Lett. 1999, 450, 101–104. [Google Scholar] [CrossRef]
- Tombola, F.; Oregna, F.; Brutsche, S.; Szabo, I.; Del Giudice, G.; Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M. Inhibition of the vacuolating and anion channel activities of the VacA toxin of Helicobacter pylori. FEBS Lett. 1999, 460, 221–225. [Google Scholar] [CrossRef]
- Szabo, I.; Brutsche, S.; Tombola, F.; Moschioni, M.; Satin, B.; Telford, J.L.; Rappuoli, R.; Montecucco, C.; Papini, E.; Zoratti, M. Formation of anion-selective channels in the cell plasma membrane by the toxin VacA of Helicobacter pylori is required for its biological activity. Embo J. 1999, 18, 5517–5527. [Google Scholar] [CrossRef] [PubMed]
- Czajkowsky, D.M.; Iwamoto, H.; Szabo, G.; Cover, T.L.; Shao, Z. Mimicry of a host anion channel by a Helicobacter pylori pore-forming toxin. Biophys. J. 2005, 89, 3093–3101. [Google Scholar] [CrossRef] [PubMed]
- Vinion-Dubiel, A.D.; McClain, M.S.; Czajkowsky, D.M.; Iwamoto, H.; Ye, D.; Cao, P.; Schraw, W.; Szabo, G.; Blanke, S.R.; Shao, Z.; et al. A dominant negative mutant of Helicobacter pylori vacuolating toxin (VacA) inhibits VacA-induced cell vacuolation. J. Biol. Chem. 1999, 274, 37736–37742. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Iwamoto, H.; Cao, P.; Vinion-Dubiel, A.D.; Li, Y.; Szabo, G.; Shao, Z.; Cover, T.L. Essential role of a GXXXG motif for membrane channel formation by Helicobacter pylori vacuolating toxin. J. Biol. Chem. 2003, 278, 12101–12108. [Google Scholar] [CrossRef] [PubMed]
- Torres, V.J.; McClain, M.S.; Cover, T.L. Interactions between p-33 and p-55 domains of the Helicobacter pylori vacuolating cytotoxin (VacA). J. Biol. Chem. 2004, 279, 2324–2331. [Google Scholar] [CrossRef] [PubMed]
- Torres, V.J.; Ivie, S.E.; McClain, M.S.; Cover, T.L. Functional properties of the p33 and p55 domains of the Helicobacter pylori vacuolating cytotoxin. J. Biol. Chem. 2005, 280, 21107–21114. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rivera, C.; Gangwer, K.A.; McClain, M.S.; Eli, I.M.; Chambers, M.G.; Ohi, M.D.; Lacy, D.B.; Cover, T.L. Reconstitution of Helicobacter pylori VacA toxin from purified components. Biochemistry 2010, 49, 5743–5752. [Google Scholar] [CrossRef] [PubMed]
- Gangwer, K.A.; Mushrush, D.J.; Stauff, D.L.; Spiller, B.; McClain, M.S.; Cover, T.L.; Lacy, D.B. Crystal structure of the Helicobacter pylori vacuolating toxin p55 domain. Proc. Natl. Acad. Sci. USA 2007, 104, 16293–16298. [Google Scholar] [CrossRef] [PubMed]
- Junker, M.; Schuster, C.C.; McDonnell, A.V.; Sorg, K.A.; Finn, M.C.; Berger, B.; Clark, P.L. Pertactin beta-helix folding mechanism suggests common themes for the secretion and folding of autotransporter proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 4918–4923. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Arico, B.; Papini, E.; Rizzuto, R.; Grandi, G.; Rappuoli, R.; Montecucco, C. Helicobacter pylori toxin VacA induces vacuole formation by acting in the cell cytosol. Mol. Microbiol. 1997, 26, 665–674. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Burroni, D.; Papini, E.; Rappuoli, R.; Telford, J.; Montecucco, C. Identification of the Helicobacter pylori VacA toxin domain active in the cell cytosol. Infect. Immun. 1998, 66, 6014–6016. [Google Scholar] [PubMed]
- Ye, D.; Willhite, D.C.; Blanke, S.R. Identification of the minimal intracellular vacuolating domain of the Helicobacter pylori vacuolating toxin. J. Biol. Chem. 1999, 274, 9277–9282. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chamberlain, A.K.; Bowie, J.U. Membrane channel structure of Helicobacter pylori vacuolating toxin: Role of multiple GXXXG motifs in cylindrical channels. Proc. Natl. Acad. Sci. USA 2004, 101, 5988–5991. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Cao, P.; Cover, T.L. Amino-terminal hydrophobic region of Helicobacter pylori vacuolating cytotoxin (VacA) mediates transmembrane protein dimerization. Infect. Immun. 2001, 69, 1181–1184. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Blanke, S.R. Mutational analysis of the Helicobacter pylori vacuolating toxin amino terminus: Identification of amino acids essential for cellular vacuolation. Infect. Immun. 2000, 68, 4354–4357. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Czajkowsky, D.M.; Torres, V.J.; Szabo, G.; Shao, Z.; Cover, T.L. Random mutagenesis of Helicobacter pylori VacA to identify amino acids essential for vacuolating cytotoxic activity. Infect. Immun. 2006, 74, 6188–6195. [Google Scholar] [CrossRef] [PubMed]
- Lupetti, P.; Heuser, J.E.; Manetti, R.; Massari, P.; Lanzavecchia, S.; Bellon, P.L.; Dallai, R.; Rappuoli, R.; Telford, J.L. Oligomeric and subunit structure of the Helicobacter pylori vacuolating cytotoxin. J. Cell Biol. 1996, 133, 801–807. [Google Scholar] [CrossRef] [PubMed]
- El-Bez, C.; Adrian, M.; Dubochet, J.; Cover, T.L. High resolution structural analysis of Helicobacter pylori VacA toxin oligomers by cryo-negative staining electron microscopy. J. Struct. Biol. 2005, 151, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Chambers, M.G.; Pyburn, T.M.; Gonzalez-Rivera, C.; Collier, S.E.; Eli, I.; Yip, C.K.; Takizawa, Y.; Lacy, D.B.; Cover, T.L.; Ohi, M.D. Structural analysis of the oligomeric states of Helicobacter pylori VacA toxin. J. Mol. Biol. 2013, 425, 524–535. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.; Blanke, S.R. Functional complementation reveals the importance of intermolecular monomer interactions for Helicobacter pylori VacA vacuolating activity. Mol. Microbiol. 2002, 43, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Willhite, D.C.; Ye, D.; Blanke, S.R. Fluorescence resonance energy transfer microscopy of the Helicobacter pylori vacuolating cytotoxin within mammalian cells. Infect. Immun. 2002, 70, 3824–3832. [Google Scholar] [CrossRef] [PubMed]
- Genisset, C.; Galeotti, C.L.; Lupetti, P.; Mercati, D.; Skibinski, D.A.; Barone, S.; Battistutta, R.; de Bernard, M.; Telford, J.L. A Helicobacter pylori vacuolating toxin mutant that fails to oligomerize has a dominant negative phenotype. Infect. Immun. 2006, 74, 1786–1794. [Google Scholar] [CrossRef] [PubMed]
- Ivie, S.E.; McClain, M.S.; Torres, V.J.; Algood, H.M.; Lacy, D.B.; Yang, R.; Blanke, S.R.; Cover, T.L. Helicobacter pylori VacA subdomain required for intracellular toxin activity and assembly of functional oligomeric complexes. Infect. Immun. 2008, 76, 2843–2851. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Hanson, P.I.; Heuser, J.E. Acid-induced dissociation of VacA, the Helicobacter pylori vacuolating cytotoxin, reveals its pattern of assembly. J. Cell Biol. 1997, 138, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Niidome, T.; Kimura, M.; Hatakeyama, T.; Aoyagi, H.; Kurazono, H.; Imagawa, K.; Wada, A.; Moss, J.; Hirayama, T. Activation of Helicobacter pylori VacA toxin by alkaline or acid conditions increases its binding to a 250-kDa receptor protein-tyrosine phosphatase beta. J. Biol. Chem. 1999, 274, 36693–36699. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Papini, E.; de Filippis, V.; Gottardi, E.; Telford, J.; Manetti, R.; Fontana, A.; Rappuoli, R.; Montecucco, C. Low pH activates the vacuolating toxin of Helicobacter pylori, which becomes acid and pepsin resistant. J. Biol. Chem. 1995, 270, 23937–23940. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Schraw, W.; Ricci, V.; Boquet, P.; Cover, T.L. Acid-activation of Helicobacter pylori vacuolating cytotoxin (VacA) results in toxin internalization by eukaryotic cells. Mol. Microbiol. 2000, 37, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, M.H.; Atherton, J.C.; Blaser, M.J.; Cover, T.L. Heterogeneity in levels of vacuolating cytotoxin gene (vacA) transcription among Helicobacter pylori strains. Infect. Immun. 1998, 66, 3088–3094. [Google Scholar] [PubMed]
- Ito, Y.; Azuma, T.; Ito, S.; Suto, H.; Miyaji, H.; Yamazaki, Y.; Kohli, Y.; Kuriyama, M. Full-length sequence analysis of the vacA gene from cytotoxic and noncytotoxic Helicobacter pylori. J. Infect. Dis. 1998, 178, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.S.; Valk, P.L.; McClain, M.S.; Shaffer, C.L.; Metcalf, J.A.; Bordenstein, S.R.; Cover, T.L. Comparative genomic analysis of east asian and non-asian Helicobacter pylori strains identifies rapidly evolving genes. PLoS ONE 2013, 8, e55120. [Google Scholar] [CrossRef] [PubMed]
- Atherton, J.C.; Cao, P.; Peek, R.M., Jr.; Tummuru, M.K.; Blaser, M.J.; Cover, T.L. Mosaicism in vacuolating cytotoxin alleles of Helicobacter pylori. Association of specific vacA types with cytotoxin production and peptic ulceration. J. Biol. Chem. 1995, 270, 17771–17777. [Google Scholar] [PubMed]
- Rhead, J.L.; Letley, D.P.; Mohammadi, M.; Hussein, N.; Mohagheghi, M.A.; Eshagh Hosseini, M.; Atherton, J.C. A new Helicobacter pylori vacuolating cytotoxin determinant, the intermediate region, is associated with gastric cancer. Gastroenterology 2007, 133, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Letley, D.P.; Atherton, J.C. Natural diversity in the n terminus of the mature vacuolating cytotoxin of Helicobacter pylori determines cytotoxin activity. J. Bacteriol. 2000, 182, 3278–3280. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Cao, P.; Iwamoto, H.; Vinion-Dubiel, A.D.; Szabo, G.; Shao, Z.; Cover, T.L. A 12-amino-acid segment, present in type s2 but not type s1 Helicobacter pylori VacA proteins, abolishes cytotoxin activity and alters membrane channel formation. J. Bacteriol. 2001, 183, 6499–6508. [Google Scholar] [CrossRef] [PubMed]
- Letley, D.P.; Rhead, J.L.; Twells, R.J.; Dove, B.; Atherton, J.C. Determinants of non-toxicity in the gastric pathogen Helicobacter pylori. J. Biol. Chem. 2003, 278, 26734–26741. [Google Scholar] [CrossRef] [PubMed]
- Pagliaccia, C.; de Bernard, M.; Lupetti, P.; Ji, X.; Burroni, D.; Cover, T.L.; Papini, E.; Rappuoli, R.; Telford, J.L.; Reyrat, J.M. The m2 form of the Helicobacter pylori cytotoxin has cell type-specific vacuolating activity. Proc. Natl. Acad. Sci. USA 1998, 95, 10212–10217. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Fernandez, T.; Burroni, D.; Pagliaccia, C.; Atherton, J.C.; Reyrat, J.M.; Rappuoli, R.; Telford, J.L. Cell specificity of Helicobacter pylori cytotoxin is determined by a short region in the polymorphic midregion. Infect. Immun. 2000, 68, 3754–3757. [Google Scholar] [CrossRef] [PubMed]
- Tombola, F.; Pagliaccia, C.; Campello, S.; Telford, J.L.; Montecucco, C.; Papini, E.; Zoratti, M. How the loop and middle regions influence the properties of Helicobacter pylori VacA channels. Biophys. J. 2001, 81, 3204–3215. [Google Scholar] [CrossRef]
- Gonzalez-Rivera, C.; Algood, H.M.; Radin, J.N.; McClain, M.S.; Cover, T.L. The intermediate region of Helicobacter pylori VacA is a determinant of toxin potency in a jurkat t cell assay. Infect. Immun. 2012, 80, 2578–2588. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.; Machado, J.C.; Pharoah, P.; Seruca, R.; Sousa, S.; Carvalho, R.; Capelinha, A.F.; Quint, W.; Caldas, C.; van Doorn, L.J.; et al. Helicobacter pylori and interleukin 1 genotyping: An opportunity to identify high-risk individuals for gastric carcinoma. J. Natl. Cancer Inst. 2002, 94, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L. Helicobacter pylori diversity and gastric cancer risk. MBio 2016, 7, e01869-15. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Vaughn, S.G.; Cao, P.; Blaser, M.J. Potentiation of Helicobacter pylori vacuolating toxin activity by nicotine and other weak bases. J. Infect. Dis. 1992, 166, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Papini, E.; de Bernard, M.; Milia, E.; Bugnoli, M.; Zerial, M.; Rappuoli, R.; Montecucco, C. Cellular vacuoles induced by Helicobacter pylori originate from late endosomal compartments. Proc. Natl. Acad. Sci. USA 1994, 91, 9720–9724. [Google Scholar] [CrossRef] [PubMed]
- Molinari, M.; Galli, C.; Norais, N.; Telford, J.L.; Rappuoli, R.; Luzio, J.P.; Montecucco, C. Vacuoles induced by Helicobacter pylori toxin contain both late endosomal and lysosomal markers. J. Biol. Chem. 1997, 272, 25339–25344. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wandinger-Ness, A.; Goldenring, J.R.; Cover, T.L. Clustering and redistribution of late endocytic compartments in response to Helicobacter pylori vacuolating toxin. Mol. Biol. Cell 2004, 15, 1946–1959. [Google Scholar] [CrossRef] [PubMed]
- Kern, B.; Jain, U.; Utsch, C.; Otto, A.; Busch, B.; Jimenez-Soto, L.; Becher, D.; Haas, R. Characterization of Helicobacter pylori VacA-containing vacuoles (VCVs), VacA intracellular trafficking and interference with calcium signalling in T lymphocytes. Cell. Microbiol. 2015, 17, 1811–1832. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Moschioni, M.; Habermann, A.; Griffiths, G.; Montecucco, C. Cell vacuolization induced by Helicobacter pylori VacA cytotoxin does not depend on late endosomal snares. Cell. Microbiol. 2002, 4, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Montecucco, C.; Rappuoli, R. Living dangerously: How Helicobacter pylori survives in the human stomach. Nat. Rev. Mol. Cell Biol. 2001, 2, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blanke, S.R. Helicobacter pylori VacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Ricci, V.; Sommi, P.; Fiocca, R.; Romano, M.; Solcia, E.; Ventura, U. Helicobacter pylori vacuolating toxin accumulates within the endosomal-vacuolar compartment of cultured gastric cells and potentiates the vacuolating activity of ammonia. J. Pathol. 1997, 183, 453–459. [Google Scholar] [CrossRef]
- Morbiato, L.; Tombola, F.; Campello, S.; Del Giudice, G.; Rappuoli, R.; Zoratti, M.; Papini, E. Vacuolation induced by VacA toxin of Helicobacter pylori requires the intracellular accumulation of membrane permeant bases, Cl(-) and water. FEBS Lett. 2001, 508, 479–483. [Google Scholar] [CrossRef]
- Genisset, C.; Puhar, A.; Calore, F.; de Bernard, M.; Dell’Antone, P.; Montecucco, C. The concerted action of the Helicobacter pylori cytotoxin VacA and of the v-atpase proton pump induces swelling of isolated endosomes. Cell. Microbiol. 2007, 9, 1481–1490. [Google Scholar] [CrossRef] [PubMed]
- Satin, B.; Norais, N.; Telford, J.; Rappuoli, R.; Murgia, M.; Montecucco, C.; Papini, E. Effect of Helicobacter pylori vacuolating toxin on maturation and extracellular release of procathepsin d and on epidermal growth factor degradation. J. Biol. Chem. 1997, 272, 25022–25028. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Noto, J.M.; Romero-Gallo, J.; Peek, R.M., Jr.; Amieva, M.R. Helicobacter pylori perturbs iron trafficking in the epithelium to grow on the cell surface. PLoS Pathog. 2011, 7, e1002050. [Google Scholar] [CrossRef] [PubMed]
- Molinari, M.; Salio, M.; Galli, C.; Norais, N.; Rappuoli, R.; Lanzavecchia, A.; Montecucco, C. Selective inhibition of Ii-dependent antigen presentation by Helicobacter pylori toxin VacA. J. Exp. Med. 1998, 187, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Terebiznik, M.R.; Raju, D.; Vazquez, C.L.; Torbricki, K.; Kulkarni, R.; Blanke, S.R.; Yoshimori, T.; Colombo, M.I.; Jones, N.L. Effect of Helicobacter pylori’s vacuolating cytotoxin on the autophagy pathway in gastric epithelial cells. Autophagy 2009, 5, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Satoh, M.; Nakano, M.; Hisatsune, J.; Isomoto, H.; Sap, J.; Suzuki, H.; Nomura, F.; Noda, M.; Moss, J.; et al. Low-density lipoprotein receptor-related protein-1 (LRP1) mediates autophagy and apoptosis caused by Helicobacter pylori VacA. J. Biol. Chem. 2012, 287, 31104–31115. [Google Scholar] [CrossRef] [PubMed]
- Raju, D.; Hussey, S.; Ang, M.; Terebiznik, M.R.; Sibony, M.; Galindo-Mata, E.; Gupta, V.; Blanke, S.R.; Delgado, A.; Romero-Gallo, J.; et al. Vacuolating cytotoxin and variants in Atg16l1 that disrupt autophagy promote Helicobacter pylori infection in humans. Gastroenterology 2012, 142, 1160–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfield, L.K.; Jones, N.L. Modulation of autophagy by Helicobacter pylori and its role in gastric carcinogenesis. Trends Microbiol. 2013, 21, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Goto, S.; Wada, A.; Yahiro, K.; Niidome, T.; Hatakeyama, T.; Aoyagi, H.; Hirayama, T.; Kondo, T. Vacuolating cytotoxin purified from Helicobacter pylori causes mitochondrial damage in human gastric cells. Microb. Pathog. 1999, 26, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Willhite, D.C.; Blanke, S.R. Helicobacter pylori vacuolating cytotoxin enters cells, localizes to the mitochondria, and induces mitochondrial membrane permeability changes correlated to toxin channel activity. Cell. Microbiol. 2004, 6, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, E.; Wada, A.; Kumatori, A.; Nakagawa, I.; Funao, J.; Nakayama, M.; Hisatsune, J.; Kimura, M.; Moss, J.; Hirayama, T. Helicobacter pylori vacuolating cytotoxin induces activation of the proapoptotic proteins Bax and Bak, leading to cytochrome c release and cell death, independent of vacuolation. J. Biol. Chem. 2006, 281, 11250–11259. [Google Scholar] [CrossRef] [PubMed]
- Galmiche, A.; Rassow, J.; Doye, A.; Cagnol, S.; Chambard, J.C.; Contamin, S.; de Thillot, V.; Just, I.; Ricci, V.; Solcia, E.; et al. The N-terminal 34 kDa fragment of Helicobacter pylori vacuolating cytotoxin targets mitochondria and induces cytochrome c release. Embo J. 2000, 19, 6361–6370. [Google Scholar] [CrossRef] [PubMed]
- Willhite, D.C.; Cover, T.L.; Blanke, S.R. Cellular vacuolation and mitochondrial cytochrome c release are independent outcomes of Helicobacter pylori vacuolating cytotoxin activity that are each dependent on membrane channel formation. J. Biol. Chem. 2003, 278, 48204–48209. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Luo, Z.Q.; Blanke, S.R. Helicobacter pylori vacuolating cytotoxin a (VacA) engages the mitochondrial fission machinery to induce host cell death. Proc. Natl. Acad. Sci. USA 2011, 108, 16032–16037. [Google Scholar] [CrossRef] [PubMed]
- Oldani, A.; Cormont, M.; Hofman, V.; Chiozzi, V.; Oregioni, O.; Canonici, A.; Sciullo, A.; Sommi, P.; Fabbri, A.; Ricci, V.; et al. Helicobacter pylori counteracts the apoptotic action of its VacA toxin by injecting the CagA protein into gastric epithelial cells. PLoS Pathog. 2009, 5, e1000603. [Google Scholar] [CrossRef] [PubMed]
- Domanska, G.; Motz, C.; Meinecke, M.; Harsman, A.; Papatheodorou, P.; Reljic, B.; Dian-Lothrop, E.A.; Galmiche, A.; Kepp, O.; Becker, L.; et al. Helicobacter pylori VacA toxin/subunit p34: Targeting of an anion channel to the inner mitochondrial membrane. PLoS Pathog. 2010, 6, e1000878. [Google Scholar] [CrossRef] [PubMed]
- Foo, J.H.; Culvenor, J.G.; Ferrero, R.L.; Kwok, T.; Lithgow, T.; Gabriel, K. Both the p33 and p55 subunits of the Helicobacter pylori VacA toxin are targeted to mammalian mitochondria. J. Mol. Biol. 2010, 401, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Tombola, F.; Morbiato, L.; Del Giudice, G.; Rappuoli, R.; Zoratti, M.; Papini, E. The Helicobacter pylori VacA toxin is a urea permease that promotes urea diffusion across epithelia. J. Clin. Investig. 2001, 108, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Debellis, L.; Papini, E.; Caroppo, R.; Montecucco, C.; Curci, S. Helicobacter pylori cytotoxin VacA increases alkaline secretion in gastric epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1440–G1448. [Google Scholar] [PubMed]
- Papini, E.; Satin, B.; Norais, N.; de Bernard, M.; Telford, J.L.; Rappuoli, R.; Montecucco, C. Selective increase of the permeability of polarized epithelial cell monolayers by Helicobacter pylori vacuolating toxin. J. Clin. Investig. 1998, 102, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Amieva, M.R.; Vogelmann, R.; Covacci, A.; Tompkins, L.S.; Nelson, W.J.; Falkow, S. Disruption of the epithelial apical-junctional complex by Helicobacter pylori CagA. Science 2003, 300, 1430–1434. [Google Scholar] [CrossRef] [PubMed]
- Pelicic, V.; Reyrat, J.M.; Sartori, L.; Pagliaccia, C.; Rappuoli, R.; Telford, J.L.; Montecucco, C.; Papini, E. Helicobacter pylori VacA cytotoxin associated with the bacteria increases epithelial permeability independently of its vacuolating activity. Microbiology 1999, 145, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Kimura, M.; Wada, A.; Yahiro, K.; Ogushi, K.I.; Niidome, T.; Fujikawa, A.; Shirasaka, D.; Aoyama, N.; Kurazono, H.; et al. Helicobacter pylori VacA activates the p38/ATF-2-mediated signal pathway in AZ-521 cells. J. Biol. Chem. 2004, 279, 7024–7028. [Google Scholar] [CrossRef] [PubMed]
- Hisatsune, J.; Yamasaki, E.; Nakayama, M.; Shirasaka, D.; Kurazono, H.; Katagata, Y.; Inoue, H.; Han, J.; Sap, J.; Yahiro, K.; et al. Helicobacter pylori VacA enhances PGE2 production through induction of COX-2 expression via a p38 MAP kinase/ATF-2 cascade in AZ-521 cells. Infect. Immun. 2007, 75, 4472–4481. [Google Scholar] [CrossRef] [PubMed]
- Boncristiano, M.; Paccani, S.R.; Barone, S.; Ulivieri, C.; Patrussi, L.; Ilver, D.; Amedei, A.; D’Elios, M.M.; Telford, J.L.; Baldari, C.T. The Helicobacter pylori vacuolating toxin inhibits T cell activation by two independent mechanisms. J. Exp. Med. 2003, 198, 1887–1897. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, A.; Shirasaka, D.; Yamamoto, S.; Ota, H.; Yahiro, K.; Fukada, M.; Shintani, T.; Wada, A.; Aoyama, N.; Hirayama, T.; et al. Mice deficient in protein tyrosine phosphatase receptor type z are resistant to gastric ulcer induction by VacA of Helicobacter pylori. Nat. Genet. 2003, 33, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Caputo, R.; Tuccillo, C.; Manzo, B.A.; Zarrilli, R.; Tortora, G.; Blanco Cdel, V.; Ricci, V.; Ciardiello, F.; Romano, M. Helicobacter pylori VacA toxin up-regulates vascular endothelial growth factor expression in MKN 28 gastric cells through an epidermal growth factor receptor-, cyclooxygenase-2-dependent mechanism. Clin. Cancer Res. 2003, 9, 2015–2021. [Google Scholar] [PubMed]
- Nakayama, M.; Hisatsune, J.; Yamasaki, E.; Isomoto, H.; Kurazono, H.; Hatakeyama, M.; Azuma, T.; Yamaoka, Y.; Yahiro, K.; Moss, J.; et al. Helicobacter pylori VacA-induced inhibition of GSK3 through the PI3K/Akt signaling pathway. J. Biol. Chem. 2009, 284, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Kuck, D.; Kolmerer, B.; Iking-Konert, C.; Krammer, P.H.; Stremmel, W.; Rudi, J. Vacuolating cytotoxin of Helicobacter pylori induces apoptosis in the human gastric epithelial cell line ags. Infect. Immun. 2001, 69, 5080–5087. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Krishna, U.S.; Israel, D.A.; Peek, R.M., Jr. Induction of gastric epithelial cell apoptosis by Helicobacter pylori vacuolating cytotoxin. Cancer Res. 2003, 63, 951–957. [Google Scholar] [PubMed]
- Radin, J.N.; Gonzalez-Rivera, C.; Ivie, S.E.; McClain, M.S.; Cover, T.L. Helicobacter pylori VacA induces programmed necrosis in gastric epithelial cells. Infect. Immun. 2011, 79, 2535–2543. [Google Scholar] [CrossRef] [PubMed]
- Radin, J.N.; Gonzalez-Rivera, C.; Frick-Cheng, A.E.; Sheng, J.; Gaddy, J.A.; Rubin, D.H.; Algood, H.M.; McClain, M.S.; Cover, T.L. Role of connexin 43 in Helicobacter pylori VacA-induced cell death. Infect. Immun. 2014, 82, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Calore, F.; Genisset, C.; Casellato, A.; Rossato, M.; Codolo, G.; Esposti, M.D.; Scorrano, L.; de Bernard, M. Endosome-mitochondria juxtaposition during apoptosis induced by H. pylori VacA. Cell Death Differ. 2010, 17, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Isomoto, H.; Nakayama, M.; Hisatsune, J.; Nishi, Y.; Nakashima, Y.; Matsushima, K.; Kurazono, H.; Nakao, K.; Hirayama, T.; et al. Helicobacter pylori VacA reduces the cellular expression of STAT3 and pro-survival Bcl-2 family proteins, Bcl-2 and Bcl-XL, leading to apoptosis in gastric epithelial cells. Dig. Dis. Sci. 2011, 56, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Akazawa, Y.; Isomoto, H.; Matsushima, K.; Kanda, T.; Minami, H.; Yamaghchi, N.; Taura, N.; Shiozawa, K.; Ohnita, K.; Takeshima, F.; et al. Endoplasmic reticulum stress contributes to Helicobacter pylori VacA-induced apoptosis. PLoS ONE 2013, 8, e82322. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.J.; Blanke, S.R. Remodeling the host environment: Modulation of the gastric epithelium by the Helicobacter pylori vacuolating toxin (VacA). Front. Cell. Infect. Microbiol. 2012, 2, 37. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Kim, J.S.; Lee, J.Y.; Kim, Y.J.; Youn, H.J.; Kim, I.Y.; Chee, Y.J.; Oh, Y.K.; Kim, N.; Jung, H.C.; et al. Vacuolating cytotoxin in Helicobacter pylori water-soluble proteins upregulates chemokine expression in human eosinophils via Ca2+ influx, mitochondrial reactive oxygen intermediates, and NF-kappaB activation. Infect. Immun. 2007, 75, 3373–3381. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Kim, J.S.; Lee, J.Y.; Sim, Y.S.; Kim, Y.J.; Oh, Y.K.; Yoon, H.J.; Kang, J.S.; Youn, J.; Kim, N.; et al. Dual effects of Helicobacter pylori vacuolating cytotoxin on human eosinophil apoptosis in early and late periods of stimulation. Eur. J. Immunol. 2010, 40, 1651–1662. [Google Scholar] [CrossRef] [PubMed]
- Supajatura, V.; Ushio, H.; Wada, A.; Yahiro, K.; Okumura, K.; Ogawa, H.; Hirayama, T.; Ra, C. Cutting edge: VacA, a vacuolating cytotoxin of Helicobacter pylori, directly activates mast cells for migration and production of proinflammatory cytokines. J. Immunol. 2002, 168, 2603–2607. [Google Scholar] [CrossRef] [PubMed]
- De Bernard, M.; Cappon, A.; Pancotto, L.; Ruggiero, P.; Rivera, J.; Del Giudice, G.; Montecucco, C. The Helicobacter pylori VacA cytotoxin activates RBL-2H3 cells by inducing cytosolic calcium oscillations. Cell. Microbiol. 2005, 7, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Kim, J.S.; Yoo, D.Y.; Ko, S.H.; Kim, N.; Kim, H.; Kim, Y.J. Stimulation of dendritic cells with Helicobacter pylori vacuolating cytotoxin negatively regulates their maturation via the restoration of E2F1. Clin. Exp. Immunol. 2011, 166, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Oertli, M.; Noben, M.; Engler, D.B.; Semper, R.P.; Reuter, S.; Maxeiner, J.; Gerhard, M.; Taube, C.; Muller, A. Helicobacter pylori gamma-glutamyl transpeptidase and vacuolating cytotoxin promote gastric persistence and immune tolerance. Proc. Natl. Acad. Sci. USA 2013, 110, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Gebert, B.; Fischer, W.; Weiss, E.; Hoffman, R.; Haas, R. Helicobacter pylori vacuolating cytotoxin inhibits T lymphocyte activation. Science 2003, 301, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Sundrud, M.S.; Torres, V.J.; Unutmaz, D.; Cover, T.L. Inhibition of primary human T cell proliferation by Helicobacter pylori vacuolating toxin (VacA) is independent of VacA effects on IL-2 secretion. Proc. Natl. Acad. Sci. USA 2004, 101, 7727–7732. [Google Scholar] [CrossRef] [PubMed]
- Torres, V.J.; VanCompernolle, S.E.; Sundrud, M.S.; Unutmaz, D.; Cover, T.L. Helicobacter pylori vacuolating cytotoxin inhibits activation-induced proliferation of human T and B lymphocyte subsets. J. Immunol. 2007, 179, 5433–5440. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.A.; Schlesinger, L.S.; Kang, B. Virulent strains of Helicobacter pylori demonstrate delayed phagocytosis and stimulate homotypic phagosome fusion in macrophages. J. Exp. Med. 2000, 191, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.Y.; Jones, N.L. Helicobacter pylori strains expressing the vacuolating cytotoxin interrupt phagosome maturation in macrophages by recruiting and retaining TACO (coronin 1) protein. Cell. Microbiol. 2003, 5, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Forster, S.; Irving, A.; Tate, M.; Ferrero, R.L.; Hertzog, P.; Frokiaer, H.; Kaparakis-Liaskos, M. Helicobacter pylori VacA suppresses lactobacillus acidophilus-induced interferon beta signaling in macrophages via alterations in the endocytic pathway. MBio 2013, 4, e00609-12. [Google Scholar] [CrossRef] [PubMed]
- Hisatsune, J.; Nakayama, M.; Isomoto, H.; Kurazono, H.; Mukaida, N.; Mukhopadhyay, A.K.; Azuma, T.; Yamaoka, Y.; Sap, J.; Yamasaki, E.; et al. Molecular characterization of Helicobacter pylori VacA induction of IL-8 in U937 cells reveals a prominent role for p38 MAPK in activating transcription factor-2, cAMP response element binding protein, and NF-kappaB activation. J. Immunol. 2008, 180, 5017–5027. [Google Scholar] [CrossRef] [PubMed]
- Menaker, R.J.; Ceponis, P.J.; Jones, N.L. Helicobacter pylori induces apoptosis of macrophages in association with alterations in the mitochondrial pathway. Infect. Immun. 2004, 72, 2889–2898. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Kamiya, S.; Suzuki, T.; Kohda, K.; Muramatsu, S.; Kurumada, T.; Ohta, U.; Miyazawa, M.; Kimura, N.; Mutoh, N.; et al. The effect of Helicobacter pylori on gastric acid secretion by isolated parietal cells from a guinea pig. Association with production of vacuolating toxin by H. pylori. Scand. J. Gastroenterol. 1996, 31, 428–433. [Google Scholar] [PubMed]
- Wang, F.; Xia, P.; Wu, F.; Wang, D.; Wang, W.; Ward, T.; Liu, Y.; Aikhionbare, F.; Guo, Z.; Powell, M.; et al. Helicobacter pylori VacA disrupts apical membrane-cytoskeletal interactions in gastric parietal cells. J. Biol. Chem. 2008, 283, 26714–26725. [Google Scholar] [CrossRef] [PubMed]
- Massari, P.; Manetti, R.; Burroni, D.; Nuti, S.; Norais, N.; Rappuoli, R.; Telford, J.L. Binding of the Helicobacter pylori vacuolating cytotoxin to target cells. Infect. Immun. 1998, 66, 3981–3984. [Google Scholar] [PubMed]
- Wang, H.J.; Wang, W.C. Expression and binding analysis of GST-VacA fusions reveals that the C-terminal approximately 100-residue segment of exotoxin is crucial for binding in hela cells [in process citation]. Biochem. Biophys. Res. Commun. 2000, 278, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-C.; Wang, H.-J.; Kuo, C.-H. Two distinctive cell binding patterns by vacuolating toxin fused with glutathione S-transferase: One high-affinity m1-specific binding and the other lower-affinity binding for variant m forms. Biochemistry 2001, 40, 11887–11896. [Google Scholar] [CrossRef] [PubMed]
- Ricci, V.; Galmiche, A.; Doye, A.; Necchi, V.; Solcia, E.; Boquet, P. High cell sensitivity to Helicobacter pylori VacA toxin depends on a GPI-anchored protein and is not blocked by inhibition of the clathrin-mediated pathway of endocytosis. Mol. Biol. Cell 2000, 11, 3897–3909. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.K.; Willhite, D.C.; Patel, R.M.; Ye, D.; Williams, C.L.; Torres, E.M.; Marty, K.B.; MacDonald, R.A.; Blanke, S.R. Plasma membrane cholesterol modulates cellular vacuolation induced by the Helicobacter pylori vacuolating cytotoxin. Infect. Immun. 2002, 70, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Wada, A.; Nakayama, M.; Kimura, T.; Ogushi, K.; Niidome, T.; Aoyagi, H.; Yoshino, K.; Yonezawa, K.; Moss, J.; et al. Protein-tyrosine phosphatase alpha, RPTP alpha, is a Helicobacter pylori VacA receptor. J. Biol. Chem. 2003, 278, 19183–19189. [Google Scholar] [CrossRef] [PubMed]
- Yahiro, K.; Wada, A.; Yamasaki, E.; Nakayama, M.; Nishi, Y.; Hisatsune, J.; Morinaga, N.; Sap, J.; Noda, M.; Moss, J.; et al. Essential domain of receptor tyrosine phosphatase beta (RPTPbeta) for interaction with Helicobacter pylori vacuolating cytotoxin. J. Biol. Chem. 2004, 279, 51013–51021. [Google Scholar] [CrossRef] [PubMed]
- Seto, K.; Hayashi-Kuwabara, Y.; Yoneta, T.; Suda, H.; Tamaki, H. Vacuolation induced by cytotoxin from Helicobacter pylori is mediated by the EGF receptor in hela cells. FEBS Lett. 1998, 431, 347–350. [Google Scholar] [CrossRef]
- Utt, M.; Danielsson, B.; Wadstrom, T. Helicobacter pylori vacuolating cytotoxin binding to a putative cell surface receptor, heparan sulfate, studied by surface plasmon resonance. FEMS Immunol. Med. Microbiol. 2001, 30, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.R.; Patel, H.K.; Kostolansky, S.S.; Ballivian, R.A.; Eichberg, J.; Blanke, S.R. Sphingomyelin functions as a novel receptor for Helicobacter pylori VacA. PLoS Pathog. 2008, 4, e1000073. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.R.; Wilson, B.A.; Blanke, S.R. Sphingomyelin is important for the cellular entry and intracellular localization of Helicobacter pylori VacA. Cell. Microbiol. 2010, 12, 1517–1533. [Google Scholar] [CrossRef] [PubMed]
- Roche, N.; Ilver, D.; Angstrom, J.; Barone, S.; Telford, J.L.; Teneberg, S. Human gastric glycosphingolipids recognized by Helicobacter pylori vacuolating cytotoxin VacA. Microbes Infect. 2007, 9, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Molinari, M.; Galli, C.; de Bernard, M.; Norais, N.; Ruysschaert, J.M.; Rappuoli, R.; Montecucco, C. The acid activation of Helicobacter pylori toxin VacA: Structural and membrane binding studies. Biochem. Biophys. Res. Commun. 1998, 248, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Schraw, W.; Li, Y.; McClain, M.S.; van der Goot, F.G.; Cover, T.L. Association of Helicobacter pylori vacuolating toxin (VacA) with lipid rafts. J. Biol. Chem. 2002, 277, 34642–34650. [Google Scholar] [CrossRef] [PubMed]
- Geisse, N.A.; Cover, T.L.; Henderson, R.M.; Edwardson, J.M. Targeting of Helicobacter pylori vacuolating toxin to lipid raft membrane domains analysed by atomic force microscopy. Biochem. J. 2004, 381, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Sewald, X.; Gebert-Vogal, B.; Prassl, S.; Barwig, I.; Weiss, E.; Fabbri, M.; Osicka, R.; Schiemann, M.; Busch, D.H.; Semmrich, M.; et al. CD18 is the T-lymphocyte receptor of the Helicobacter pylori vacuolating cytotoxin. Cell Host Microbe 2008, 3, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, N.C.; Ricci, V.; Gounon, P.; Doye, A.; Tauc, M.; Poujeol, P.; Boquet, P. Glycosylphosphatidylinositol-anchored proteins and actin cytoskeleton modulate chloride transport by channels formed by the Helicobacter pylori vacuolating cytotoxin VacA in Hela cells. J. Biol. Chem. 2004, 279, 9481–9489. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, N.C.; Monzo, P.; Kaddai, V.; Doye, A.; Ricci, V.; Boquet, P. Helicobacter pylori VacA cytotoxin: A probe for a clathrin-independent and Cdc42-dependent pinocytic pathway routed to late endosomes. Mol. Biol. Cell 2005, 16, 4852–4866. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, N.C.; Monzo, P.; Gonzalez, T.; Doye, A.; Oldani, A.; Gounon, P.; Ricci, V.; Cormont, M.; Boquet, P. Early endosomes associated with dynamic f-actin structures are required for late trafficking of H. pylori VacA toxin. J. Cell Biol. 2007, 177, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Sewald, X.; Jimenez-Soto, L.; Haas, R. Pkc-dependent endocytosis of the Helicobacter pylori vacuolating cytotoxin in primary t lymphocytes. Cell. Microbiol. 2011, 13, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Eaton, K.A.; Cover, T.L.; Tummuru, M.K.; Blaser, M.J.; Krakowka, S. Role of vacuolating cytotoxin in gastritis due to Helicobacter pylori in gnotobiotic piglets. Infect. Immun. 1997, 65, 3462–3464. [Google Scholar] [PubMed]
- Wirth, H.P.; Beins, M.H.; Yang, M.; Tham, K.T.; Blaser, M.J. Experimental infection of mongolian gerbils with wild-type and mutant Helicobacter pylori strains. Infect. Immun. 1998, 66, 4856–4866. [Google Scholar] [PubMed]
- Ogura, K.; Maeda, S.; Nakao, M.; Watanabe, T.; Tada, M.; Kyutoku, T.; Yoshida, H.; Shiratori, Y.; Omata, M. Virulence factors of Helicobacter pylori responsible for gastric diseases in mongolian gerbil. J. Exp. Med. 2000, 192, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Salama, N.R.; Otto, G.; Tompkins, L.; Falkow, S. Vacuolating cytotoxin of Helicobacter pylori plays a role during colonization in a mouse model of infection. Infect. Immun. 2001, 69, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.A.; Letley, D.P.; Cook, K.W.; Rhead, J.L.; Zaitoun, A.A.; Ingram, R.J.; Amilon, K.R.; Croxall, N.J.; Kaye, P.V.; Robinson, K.; et al. A role for the vacuolating cytotoxin, VacA, in colonization and Helicobacter pylori-induced metaplasia in the stomach. J. Infect. Dis. 2014, 210, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Ghiara, P.; Marchetti, M.; Blaser, M.J.; Tummuru, M.K.; Cover, T.L.; Segal, E.D.; Tompkins, L.S.; Rappuoli, R. Role of the Helicobacter pylori virulence factors vacuolating cytotoxin, CagA, and urease in a mouse model of disease. Infect. Immun. 1995, 63, 4154–4160. [Google Scholar] [PubMed]
- Shirasaka, D.; Aoyama, N.; Sakashita, M.; Kuroda, K.; Maekawa, S.; Wambura, C.M.; Miyamoto, M.; Tamura, T.; Yahiro, K.; Wada, A.; et al. Relationship between gastric ulcer and Helicobacter pylori VacA detected in gastric juice using bead-ELISA method. Helicobacter 2002, 7, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Cao, P.; Murthy, U.K.; Sipple, M.S.; Blaser, M.J. Serum neutralizing antibody response to the vacuolating cytotoxin of Helicobacter pylori. J. Clin. Investig. 1992, 90, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Perez-Perez, G.I.; Peek, R.M., Jr.; Atherton, J.C.; Blaser, M.J.; Cover, T.L. Detection of anti-VacA antibody responses in serum and gastric juice samples using type s1/m1 and s2/m2 Helicobacter pylori VacA antigens. Clin. Diagn. Lab. Immunol. 1999, 6, 489–493. [Google Scholar] [PubMed]
- Marchetti, M.; Aricò, B.; Burroni, D.; Figura, N.; Rappuoli, R.; Ghiara, P. Development of a mouse model of Helicobacter pylori infection that mimics human disease. Science 1995, 267, 1655–1658. [Google Scholar] [CrossRef] [PubMed]
- Ghiara, P.; Rossi, M.; Marchetti, M.; Di Tommaso, A.; Vindigni, C.; Ciampolini, F.; Covacci, A.; Telford, J.L.; De Magistris, M.T.; Pizza, M.; et al. Therapeutic intragastric vaccination against Helicobacter pylori in mice eradicates an otherwise chronic infection and confers protection against reinfection. Infect. Immun. 1997, 65, 4996–5002. [Google Scholar] [PubMed]
- Marchetti, M.; Rossi, M.; Giannelli, V.; Giuliani, M.M.; Pizza, M.; Censini, S.; Covacci, A.; Massari, P.; Pagliaccia, C.; Manetti, R.; et al. Protection against Helicobacter pylori infection in mice by intragastric vaccination with H. pylori antigens is achieved using a non-toxic mutant of E. coli heat-labile enterotoxin (LT) as adjuvant. Vaccine 1998, 16, 33–37. [Google Scholar] [CrossRef]
- Rossi, G.; Ruggiero, P.; Peppoloni, S.; Pancotto, L.; Fortuna, D.; Lauretti, L.; Volpini, G.; Mancianti, S.; Corazza, M.; Taccini, E.; et al. Therapeutic vaccination against Helicobacter pylori in the beagle dog experimental model: Safety, immunogenicity, and efficacy. Infect. Immun. 2004, 72, 3252–3259. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Schultze, V.; Rosenkranz, B.; Kaufmann, S.H.; Ulrichs, T.; Novicki, D.; Norelli, F.; Contorni, M.; Peppoloni, S.; Berti, D.; et al. Safety and immunogenicity of an intramuscular Helicobacter pylori vaccine in noninfected volunteers: A phase I study. Gastroenterology 2008, 135, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.Y.; Shi, Y.; Luo, P.; Yu, S.; Chen, L.; Zhao, Z.; Mao, X.H.; Guo, G.; Wu, C.; Zou, Q.M. Therapeutic efficacy of oral immunization with attenuated salmonella typhimurium expressing Helicobacter pylori CagA, VacA and UreB fusion proteins in mice model. Vaccine 2011, 29, 6679–6685. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Blaser, M.J. Helicobacter pylori in health and disease. Gastroenterology 2009, 136, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Atherton, J.C.; Blaser, M.J. Coadaptation of Helicobacter pylori and humans: Ancient history, modern implications. J. Clin. Investig. 2009, 119, 2475–2487. [Google Scholar] [CrossRef] [PubMed]
- Oertli, M.; Sundquist, M.; Hitzler, I.; Engler, D.B.; Arnold, I.C.; Reuter, S.; Maxeiner, J.; Hansson, M.; Taube, C.; Quiding-Jarbrink, M.; et al. DC-derived IL-18 drives Treg differentiation, murine Helicobacter pylori-specific immune tolerance, and asthma protection. J. Clin. Investig. 2012, 122, 1082–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, D.B.; Reuter, S.; van Wijck, Y.; Urban, S.; Kyburz, A.; Maxeiner, J.; Martin, H.; Yogev, N.; Waisman, A.; Gerhard, M.; et al. Effective treatment of allergic airway inflammation with Helicobacter pylori immunomodulators requires BATF3-dependent dendritic cells and IL-10. Proc. Natl. Acad. Sci. USA 2014, 111, 11810–11815. [Google Scholar] [CrossRef] [PubMed]
- Smoot, D.T.; Resau, J.H.; Earlington, M.H.; Simpson, M.; Cover, T.L. Effects of Helicobacter pylori vacuolating cytotoxin on primary cultures of human gastric epithelial cells. Gut 1996, 39, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.R.; Cover, T.L.; Crowe, D.R.; Orenstein, J.M.; Graham, M.F.; Blaser, M.J.; Smith, P.D. Helicobacter pylori cytotoxin induces vacuolation of primary human mucosal epithelial cells. Infect. Immun. 1996, 64, 4867–4871. [Google Scholar] [PubMed]
- Fiocca, R.; Luinetti, O.; Villani, L.; Chiaravalli, A.M.; Capella, C.; Solcia, E. Epithelial cytotoxicity, immune responses, and inflammatory components of Helicobacter pylori gastritis. Scand. J. Gastroenterol. Suppl. 1994, 205, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Terebiznik, M.R.; Vazquez, C.L.; Torbicki, K.; Banks, D.; Wang, T.; Hong, W.; Blanke, S.R.; Colombo, M.I.; Jones, N.L. Helicobacter pylori VacA toxin promotes bacterial intracellular survival in gastric epithelial cells. Infect. Immun. 2006, 74, 6599–6614. [Google Scholar] [CrossRef] [PubMed]
- Amieva, M.R.; Salama, N.R.; Tompkins, L.S.; Falkow, S. Helicobacter pylori enter and survive within multivesicular vacuoles of epithelial cells. Cell. Microbiol. 2002, 4, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Argent, R.H.; Thomas, R.J.; Letley, D.P.; Rittig, M.G.; Hardie, K.R.; Atherton, J.C. Functional association between the Helicobacter pylori virulence factors VacA and CagA. J. Med. Microbiol. 2008, 57, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Higashi, H.; Ishikawa, S.; Fujii, Y.; Kondo, S.; Kato, H.; Azuma, T.; Wada, A.; Hirayama, T.; Aburatani, H.; et al. Functional antagonism between Helicobacter pylori CagA and vacuolating toxin VacA in control of the NFAT signaling pathway in gastric epithelial cells. Proc. Natl. Acad. Sci. USA 2005, 102, 9661–9666. [Google Scholar] [CrossRef] [PubMed]
- Tegtmeyer, N.; Zabler, D.; Schmidt, D.; Hartig, R.; Brandt, S.; Backert, S. Importance of EGF receptor, Her2/Neu and Erk1/2 kinase signalling for host cell elongation and scattering induced by the Helicobacter pylori CagA protein: Antagonistic effects of the vacuolating cytotoxin VacA. Cell. Microbiol. 2009, 11, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Akada, J.K.; Aoki, H.; Torigoe, Y.; Kitagawa, T.; Kurazono, H.; Hoshida, H.; Nishikawa, J.; Terai, S.; Matsuzaki, M.; Hirayama, T.; et al. Helicobacter pylori CagA inhibits endocytosis of cytotoxin VacA in host cells. Dis. Models Mech. 2010, 3, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Suzuki, H.; Saya, H.; Hatakeyama, M.; Hirayama, T.; Hirata, K.; Nagano, O.; Matsuzaki, J.; Hibi, T. Reactive oxygen species-induced autophagic degradation of Helicobacter pylori CagA is specifically suppressed in cancer stem-like cells. Cell Host Microbe 2012, 12, 764–777. [Google Scholar] [CrossRef] [PubMed]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foegeding, N.J.; Caston, R.R.; McClain, M.S.; Ohi, M.D.; Cover, T.L. An Overview of Helicobacter pylori VacA Toxin Biology. Toxins 2016, 8, 173. https://doi.org/10.3390/toxins8060173
Foegeding NJ, Caston RR, McClain MS, Ohi MD, Cover TL. An Overview of Helicobacter pylori VacA Toxin Biology. Toxins. 2016; 8(6):173. https://doi.org/10.3390/toxins8060173
Chicago/Turabian StyleFoegeding, Nora J., Rhonda R. Caston, Mark S. McClain, Melanie D. Ohi, and Timothy L. Cover. 2016. "An Overview of Helicobacter pylori VacA Toxin Biology" Toxins 8, no. 6: 173. https://doi.org/10.3390/toxins8060173