1. Introduction
Fusarium spp. are among the most important pathogenic fungal communities affecting crops. These fungi can induce serious diseases on roots, stems, leaves, and fruits, resulting in huge economic losses. Maize ear rot caused by
Fusarium spp. affects maize production and kernel quality. The disease occurs in many countries; in recent years, it has gradually expanded because of global warming [
1,
2,
3]. In addition to causing ear rot and reducing yield,
Fusarium spp. produce mycotoxins that are directly synthesized in kernels and accumulated there, hence, seriously threatening the health of both humans and livestock [
4,
5,
6]. The most common
Fusarium toxins are deoxynivalenol (DON) and the fumonisins. They are very often detected in cereal crops such as maize, wheat, and rice and have drawn wide attention owing to their toxicity and carcinogenicity. In 1973,
Fusarium toxin was listed among the 16 research priorities of the World Health Organization and the Food and Agriculture Organization [
7,
8,
9,
10]. With improvements in living standards, higher requirements for food safety have been proposed.
Fusarium verticillioides is one of the most common pathogens causing ear rot in maize. This disease is widespread in temperate and semitropical areas, including all European maize-growing areas, and even prevailing in Mediterranean areas, such as the agricultural regions of Italy, which are characterized by dry, warm conditions [
3,
11]. In recent years,
F. verticillioides has also been characterized as an important pathogen causing maize ear rot in several regions of China [
12,
13,
14]. This fungus is able to produce dangerous fumonisins that can be toxic for humans and animals. The fumonisins have been classified into four main groups: fumonisins A, B, C, and P. The fumonisin B (FB) analogs—comprising toxicologically important FB
1, FB
2, and FB
3—are the most abundant naturally-occurring fumonisins. FB
1 is the most toxic and appears in the highest concentration in the host, accounting for 70% to 80% of total fumonisins [
5,
15,
16]. This mycotoxin can be carcinogenic in humans and livestock and even cause equine leukoencephalomalacia, rat hepatocarcinoma, and porcine pulmonary edema [
3,
7].
Production of fumonisins by
F. verticillioides is dependent on a biosynthetic gene cluster (
FUM) made up of 16 contiguous and coexpressed genes; deletion of the gene
FUM1 can reduce FB
1 production by 99% [
17,
18]. The various toxicological profiles of
F. verticillioides isolates may reflect significant differences in the risk for mycotoxin contamination, with potential implications for human and animal health and international trade [
19].
Fusarium graminearum species complex (FGSC) comprises a class of important pathogens of small-grain cereals and maize in many areas of the world. These fungi often cause economically devastating diseases that occur at different stages of plant development, such as seedling blight, maize root rot, wheat head blight (FHB or scab), and maize ear rot, causing a significant eventual reduction in the quality and yield of the crops [
20,
21,
22].
Moreover, these diseases are often associated with various trichothecenes and other mycotoxins produced in plants, thus representing an important problem of food safety. FGSC can produce zearalenone (ZEN), nivalenol (NIV), and DON, which can inhibit protein synthesis by combining with the 60S ribosomal subunit. This poses a significant threat to the health of both humans and livestock [
5,
23].
Fusarium spp. can produce many types of trichothecenes. Type B trichothecenes are among the mycotoxins produced by the FGSC. The particularly important groups within this type are DON and its acetylated derivatives, 3-acetyl-deoxynivalenol (3-ADON) and 15-acetyl-deoxynivalenol (15-ADON), as well as nivalenol (NIV) and its acetylated derivatives, 4-acetylnivalenol. 3-ADON has been shown to be more phytotoxic and fungi that produce 3-ADON have a higher pathogenic potential than those producing 15-ADON [
23,
24,
25].
Among specimens of
F. graminearum isolated from barley, wheat, potato, and beet in America, 15-ADON–producing isolates were more common than those producing 3-ADON, and NIV was detected only in the isolate number 21 [
26,
27]. In North America, 3-ADON–producing isolates have been found to produce a higher DON content, and these may be replacing 15-ADON isolates with weaker pathogenicity [
28,
29]. In northern Europe and northwestern Russia, more 3-ADON isolates were present as compared with 15-ADON isolates [
30,
31,
32]. The mycotoxins 15-ADON and NIV were produced by
F. graminearum in Luxembourg and 15-ADON was the dominant type, while no 3-ADON–producing isolates were detected [
33]. Of 42
F. culmorum isolates collected in Australia and nine European countries, 34 isolates produced DON and seven produced high concentrations of NIV but low levels of DON, while one produced neither DON nor NIV [
34]. A recent study showed that in
F. graminearum collected from Europe, the predominant genotype was 15-ADON (82.9%), followed by 3-ADON (13.6%), and nivalenol (NIV) (3.5%). In
F. culmorum, the prevalent genotype was 3-ADON (59.9%), while the NIV genotype accounted for 40.1% [
35].
Many methods can be used to detect
Fusarium mycotoxins, including enzyme-linked immunosorbent assay, thin-layer chromatography, capillary electrophoresis, gas chromatography, gas chromatography/mass spectrometry, high-performance liquid chromatography (HPLC), liquid chromatography/mass spectrography, and ultra-high-performance liquid chromatography/mass spectrometry (UHPLC-MS/MS). HPLC and UHPLC-MS/MS are widely used in quantitative assays of mycotoxin because of their high efficiency and accuracy [
36,
37,
38,
39].
A good knowledge of the potential for mycotoxin biosynthesis by fungal isolates from different geographic regions could help to predict the risk of contamination by mycotoxins in the surveyed areas. To date, very little information on chemotypes and the mycotoxin-producing capacity of the main pathogenic Fusarium spp. causing maize ear rot in China has been reported. In the research documented here, samples were collected from the main maize producing areas experiencing frequent ear rot; pathogenic Fusarium spp. were then isolated and identified. Mycotoxin chemotypes and their production were examined. The aim was to determine the pathogenic species of Fusarium causing maize ear rot in China, as well as the main mycotoxin chemotypes and their production capacity.
2. Results
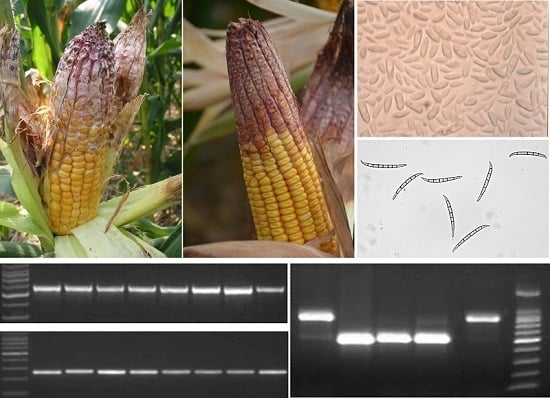
2.1. Identification of Fusarium spp.
According to morphological and molecular findings, 160 tested maize seed samples, accounting for 66.9% of total samples, were infected by
Fusarium spp. and most of them were
F. verticillioides and FGSC. 82
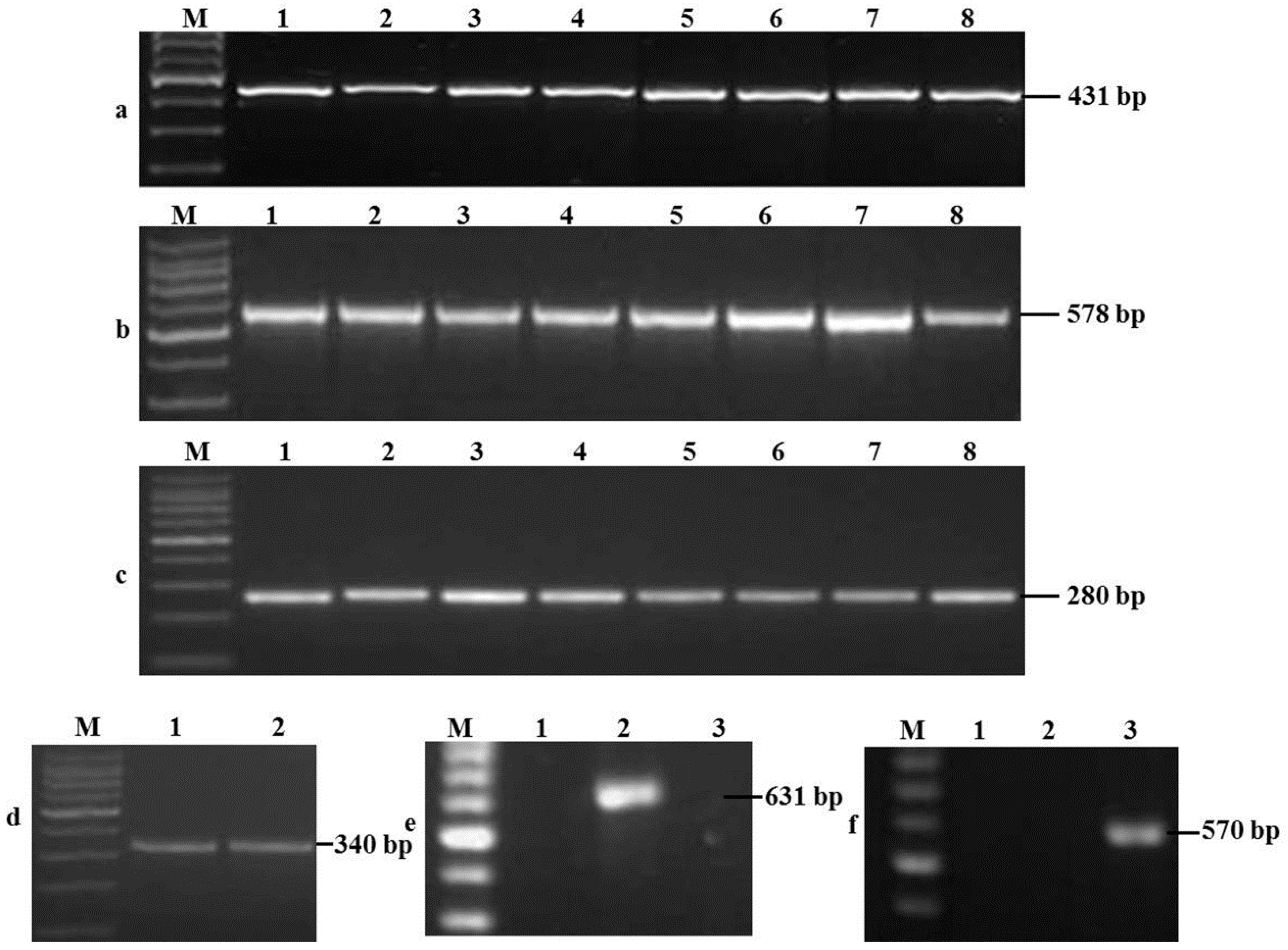
F. verticillioides isolates with the specific amplicon of 578 bp size were obtained from 82 samples (51.3%) at 67 places in 18 provinces, whereas FGSC was isolated from 70 samples (43.8%) at 66 places in 17 provinces. In addition,
F. culmorum,
F. oxysporum,
F. proliferatum,
F. subglutinans, and
F. solani clades were isolated from three (1.9%), two (1.3%), one (0.6%), one (0.6%), and one (0.6%) sample, respectively (
Figure 1).
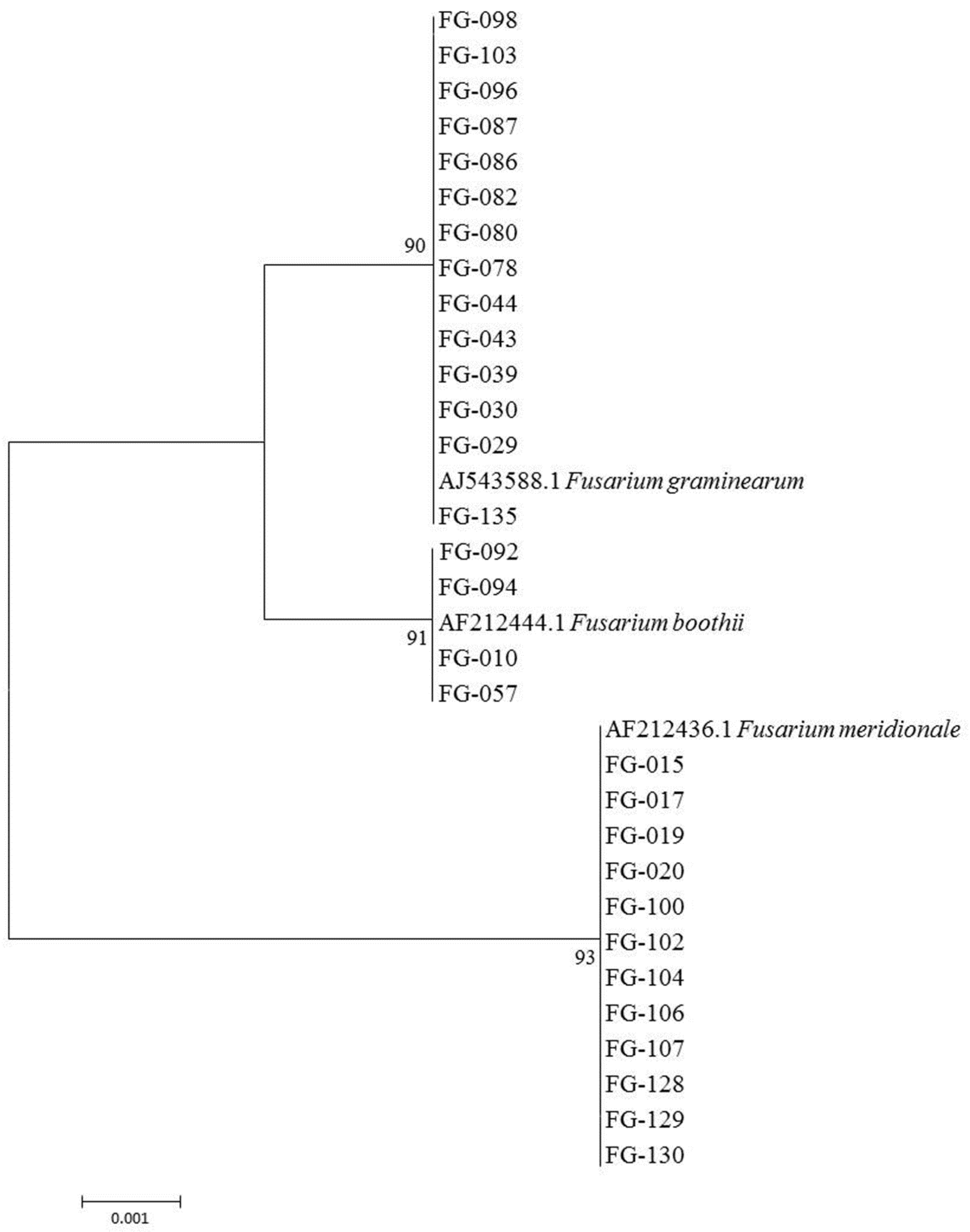
Sequencing of the
TEF-1α gene of 78 FGSC isolates and alignment with BLAST in the Fusarium Center’s database showed 99% to 100% homology between the tested FGSC and standard reference strains in this database. A total of 42 isolates—such as FG-029, FG-30, FG-043, FG-103 and others—exhibited 99% to 100% homology with
F. graminearum sensu stricto (AJ543588.1), while the other 20 isolates (e.g., FG-015, FG-020, FG-100, FG-130, and the like) showed 99% to 100% homology with
F. meridionale (AF212436.1). In addition, 16 isolates (e.g., FG-010, FG-057, and FG-094) expressed 100% homology with
F. boothii (AF212444.1). The phylogenetic tree of some FGSC based on
TEF-1α gene sequences confirmed
Fusarium species. The tree topologies of the
TEF-1α gene sequences showed the classification of FGSC into three distinct clades and the support values for these clades were high (no lower than 90% bootstrap value; BP) (
Figure 2). These results demonstrated that the FGSC causing maize ear rot in China is mainly
F. graminearum, which accounted for 53.8% of the total FGSC and was widely distributed in northern and southern maize-producing regions. The second main lineage was
F. meridionale, accounting for 25.6% of total FGSC. This lineage was isolated only from Yunnan, Guizhou, and southern Shaanxi provinces. A total of 16 isolates showed 100% homology with
F. boothii, accounting for 20.5% of the total; these were discovered in northern areas of China (e.g., Inner Mongolia, Jilin, Hebei, Shanxi, and Beijing).
2.2. Detection of Toxigenic Genes and Chemotypes
FUM1 was detected in 82 F. verticillioides isolates, thus indicating that these isolates can potentially synthesize FBs.
Three different mycotoxin chemotypes were detected from 78 isolates in FGSC. In all, 60 isolates with the specific 583-bp amplified fragments were 15-ADON, while 16 isolates producing the specific 859-bp amplified fragments were NIV. In addition, 583- and 859-bp amplified fragments produced by two isolates simultaneously were 15-ADON and NIV.
According to the molecular identification of NIV, 15-ADON, and 3-ADON of FGSC, 62 FGSC isolates collected from Beijing, Hebei, Liaoning, Jilin, Heilongjiang, Inner Mongolia, Henan, Shandong, Anhui, Ningxia, and Gansu were detected with only 15-ADON. Those in Guizhou were detected with only NIV. Both 15-ADON- and NIV-producing isolates were detected in samples from Shaanxi Province. Samples collected in Shaanxi Province had the most mycotoxin types. Isolates from northern Shaanxi had two main mycotoxin types, but 15-ADON was dominant in isolates from Guanzhong and northern Shaanxi Province. All 16 independent NIV isolates were identified as F. meridionale and mainly originated from Southwestern China. The two isolates containing specific 15-ADON- and NIV-producing segments were from southern Shaanxi.
2.3. Detection of Toxigenic Capacity
The results of FB detection with immunoafinity chromatography purification-HPLC and DON and ZEN assays based on UPLC-MS/MS agreed with the molecular identification of key mycotoxin-producing genes.
A fumonisin assay showed that the mycotoxin-producing capacity of
F. verticillioides isolates from different regions varied greatly and that FB
1 was the main chemotype. Among 82
F. verticillioides isolates, 79, 69, and 75 isolates produced FB
1, FB
2, and FB
3 respectively. A total of 70 isolates were able to produce FB
1, FB
2, and FB
3 simultaneously, and 12 isolates produced no FB
2. Among the latter, seven produced no FB
3. Among isolates producing FB, 57 isolates produced mainly FB
1 and 23 produced mainly FB
2 or FB
3. The production capacities of FB
1, FB
2, and FB
3 ranged from 2.52 to 18,416.44, 3.00 to 1864.90, and 0.28 to 1510.54 µg/g of dry hyphal weight, respectively (
Table 1).
Among FGSC, 15-ADON was detected in 68 isolates, showing a mycotoxin production range of 5.43 to 81,539.49 µg/g of dry hyphal weight. Among 10 isolates producing no 15-ADON, eight were
F. meridionale, which produced NIV according to molecular identification; they were mainly distributed in Hubei, Sichuan, and Bijie of Guizhou. A total of 22 isolates had a low mycotoxin-producing capacity (≤1000 µg/g of dry hyphal weight). Among these 22 isolates, 12 were
F. meridionale, including 11 NIV isolates from Guizhou, Yunnan, and Shangluo in southern Shaanxi Province. A total of 26 isolates had a moderate mycotoxin-producing capacity (from 1001 to 10,000 µg/g of dry hyphal weight). All of these were
F. graminearum and
F. boothii. Meanwhile 20 isolates had a high mycotoxin-producing capacity (>10,000 µg/g of hypha), all of which were
F. graminearum and
F. boothii (
Table 2).
3-ADON was detected in 65 isolates, which showed a mycotoxin-producing capacity ranging from 6.04 to 19,590.61 µg/g of dry hyphal weight. Among 13 isolates producing no 3-ADON, 10 were F. meridionale and eight were NIV-producing isolates. They were all distributed in Hubei, Sichuan, Guizhou, and Yunnan. A total of 23 isolates had a low mycotoxin-producing capacity (≤1000 µg/g of hypha), including 10 F. meridionale isolates, while 39 isolates had a moderate mycotoxin-producing capacity (1001–10,000 µg/g of dry hyphal weight), including 27 F. graminearum and 12 F. boothii isolates. Three isolates with a high mycotoxin-producing capacity (>10,000 µg/g of dry hyphal weight) were F. graminearum.
DON was detected in 51 isolates, which showed a mycotoxin-producing range of 13.35 to 19,795.33 µg/g of dry hyphal weight. A total of 27 isolates, including 21
F. meridionale, 4
F. graminearum, and two
F. boothii isolates, did not produce DON. A total of 16 isolates had a low mycotoxin-producing capacity (≤1000 µg/g of hypha), whereas 31 isolates had a moderate mycotoxin-producing capacity (1001–10,000 µg/g of dry hyphal weight), including 20
F. graminearum and 11
F. boothii isolates. Meanwhile, four
F. graminearum isolates had a high mycotoxin-producing capacity (>10,000 µg/g of dry hyphal weight) (
Table 2).
ZEN was detected in 24 isolates, which showed a mycotoxin-producing capacity varying from 1.77 to 430.24 µg/g of dry hyphal weight. Of these isolates, 13 were
F. graminearum, nine were
F. boothii, and two were
F. meridionale. However, 54 isolates did not produce ZEN (
Table 2).
Overall, F. graminearum had the highest mycotoxin-producing capacity, followed by F. boothii and F. meridionale successively in the laboratory. F. graminearum and F. boothii isolates with a high mycotoxin producing capacity were distributed mainly in Northern China.
Isolates with different chemotypes produced different amounts of mycotoxin. According to molecular identification and mycotoxin assay, 15-ADON–producing isolates produced more 15-ADON than DON and 3-ADON. None of the 18 isolates carrying NIV-specific segment produced any DON and 16 of them did nor ZEN. Most NIV-producing isolates produced more 3-ADON than 15-ADON. Also, the isolates carrying both 15-ADON and NIV chemotype-specific segments produced more 3-ADON than 15-ADON (
Table 2). As a result, the isolates with NIV chemotype-specific segments likely pose a greater risk to maize crops as well as to humans and livestock because 3-ADON is more toxic than 15-ADON.
3. Discussion
Many pathogenic
Fusarium spp. can be found causing maize ear rot in China, but the majority of isolates are determined to be
F. verticillioides and FGSC. According to the frequency of the isolation of these species from different regions, maize ear rot is most commonly caused by
F. verticillioides, followed by FGSC. This result is similar to that reported by other previous studies of the predominant pathogens causing ear rot in China, but compared with the regional research, there are also differences [
12,
13,
40].
Maize samples infected by
F. verticillioides are often contaminated with FBs. With regard to the levels of fumonisins produced by the isolates, three of 82 isolates (0.037%) were unable to produce fumonisins, indicating the presence of phenotypic variability among the
F. verticillioides isolates. Interestingly,
FUM1, a key gene responsible for FB biosynthesis, was detected in all 82 isolates. This result was similar to that of previous studies. Sanchèz-Rangel
et al. reported that some isolates of
F. verticillioides from maize in Mexico possessed the
FUM1 gene but did not produce fumonisins [
41]. Several
F. verticillioides isolates from Italy also carried the
FUM1 gene but did not produce fumonisins [
3]. These phenotypes could be explained by a mutation in the
FUM cluster or by altered expression of the
FUM genes.
In this study, the toxigenic capacity of isolates from different regions varied, indicating that under conditions of laboratory incubation the mycotoxin chemotype and toxigenic capacity of F. verticillioides may not be correlated with its region of origin. Actually, in addition to the particular isolate, mycotoxin synthesis in plants also depends heavily on environment (temperature, humidity, pH, and lighting) as well as crop rations in the field. Therefore, mycotoxin production in the laboratory primarily represents toxigenic potentiality.
The FGSC can cause various gramineous crops diseases and leads to a reduction in crop yield and quality. In recent years, a total of 16 species in FGSC have been identified [
42,
43,
44]. In the present study, three species lineages were found in the FGSC isolates collected mainly from Northeast China, Huang-Huai-Hai region, and Southwest China, regions in which the environments vary greatly.
In the present study, a total of 3 toxigenic chemotypes produced by FGSC were detected and DON was the main one. All F. graminearum and F. boothii isolates were 15-ADON–producing isolates, but F. meridionale produced one or more isolates of 15-ADON, NIV, and 3-ADON, which indicated that lineage or species was related to the mycotoxin chemotype to a certain extent. The genetic identification (molecular identification) of 10 isolates disagreed with the UHPLC assay: four were 15-ADON isolates (FG039, FG147, FG151, and FG152) and six were NIV isolates (FG017, FG140, FG142, FG143, FG145, and FG146). No mycotoxin had been detected by UHPLC in these isolates. In addition, based on the UHPLC quantitative assay of mycotoxin production, some amount of DON, 3-ADON, or ZEN was detected in 15-ADON–producing isolates and some 15-ADON or 3-ADON was detected in NIV-producing isolates. This outcome may be related to the expression and regulation of the toxin gene.
In the present study, 15-ADON–producing isolates generated more 15-ADON than 3-ADON, whereas NIV-type isolates generated more 3-ADON than 15-ADON, which may be related to the prior expression of a certain mycotoxin. Some DON-producing isolates coproduced a certain amount of NIV, as observed by Sugiura
et al. [
45].The mycotoxin production varied greatly among isolates from different regions. The occurrence of
Fusarium spp. isolates with strong toxicity or high mycotoxin production should be given particular attention. Furthermore, two isolates were found to carry both DON and NIV chemotype-specific segments, possibly resulting from recombination between the two chemotypes. Kim
et al. reported that a
F. graminearum isolate (A18) from corn generated a chimeric pattern containing features of both DON-specific and NIV-specific sequences [
46], which was similar to FG102 and FG105. With regard to the isolates carrying diverse chemotype-specific regions, special care must be taken, since these isolates have the potential to produce a variety of mycotoxins.
In summary, F. verticillioides and FGSC were the predominant fungi causing maize ear rot in China and the latter contained F. graminearum, F. meridionale, and F. boothii. Most of the F. verticillioides isolates mainly produced highly toxic fumonisin B1. The analysis of the current Fusarium chemotypes showed that trichothecene genotype was 15-ADON in all F. graminearum and F. boothii isolates. The prevalent chemotype was NIV (76.2%), followed by 15-ADON (14.3%) and NIV + 15-ADON (9.5%) in F. meridionale. The toxogenic capacity of F. graminearum and F. boothii was higher than that of F. meridionale.






{kind=link}
{kind=link}
{kind=link}