
Purification and Characterization of a Novel Kazal-Type Trypsin Inhibitor from the Leech of Hirudinaria manillensis

 , ,
, , 

Abstract
:
1. Introduction
2. Results
2.1. Purification of Bdellin-HM
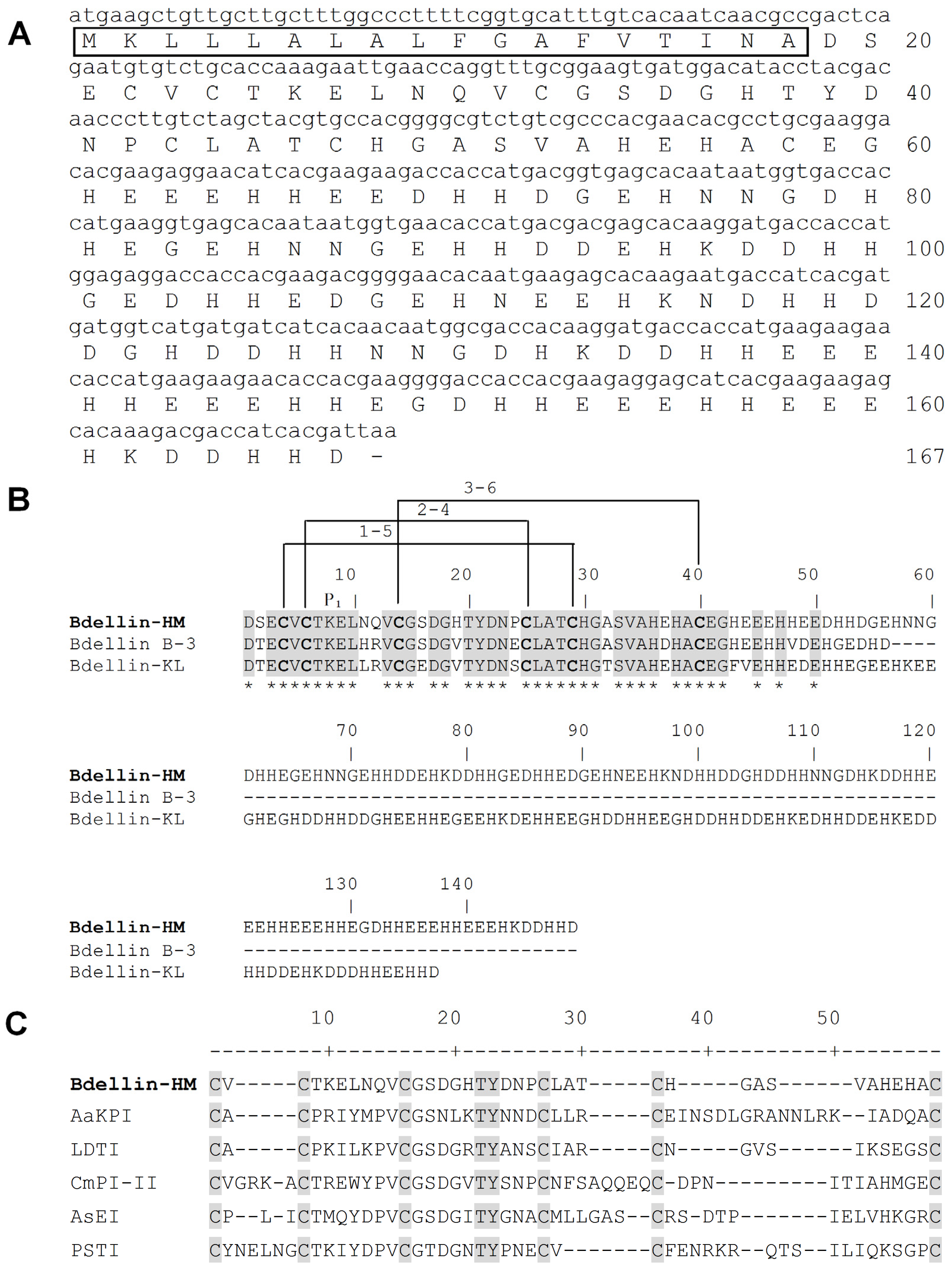
2.2. Primary Structure of Bdellin-HM
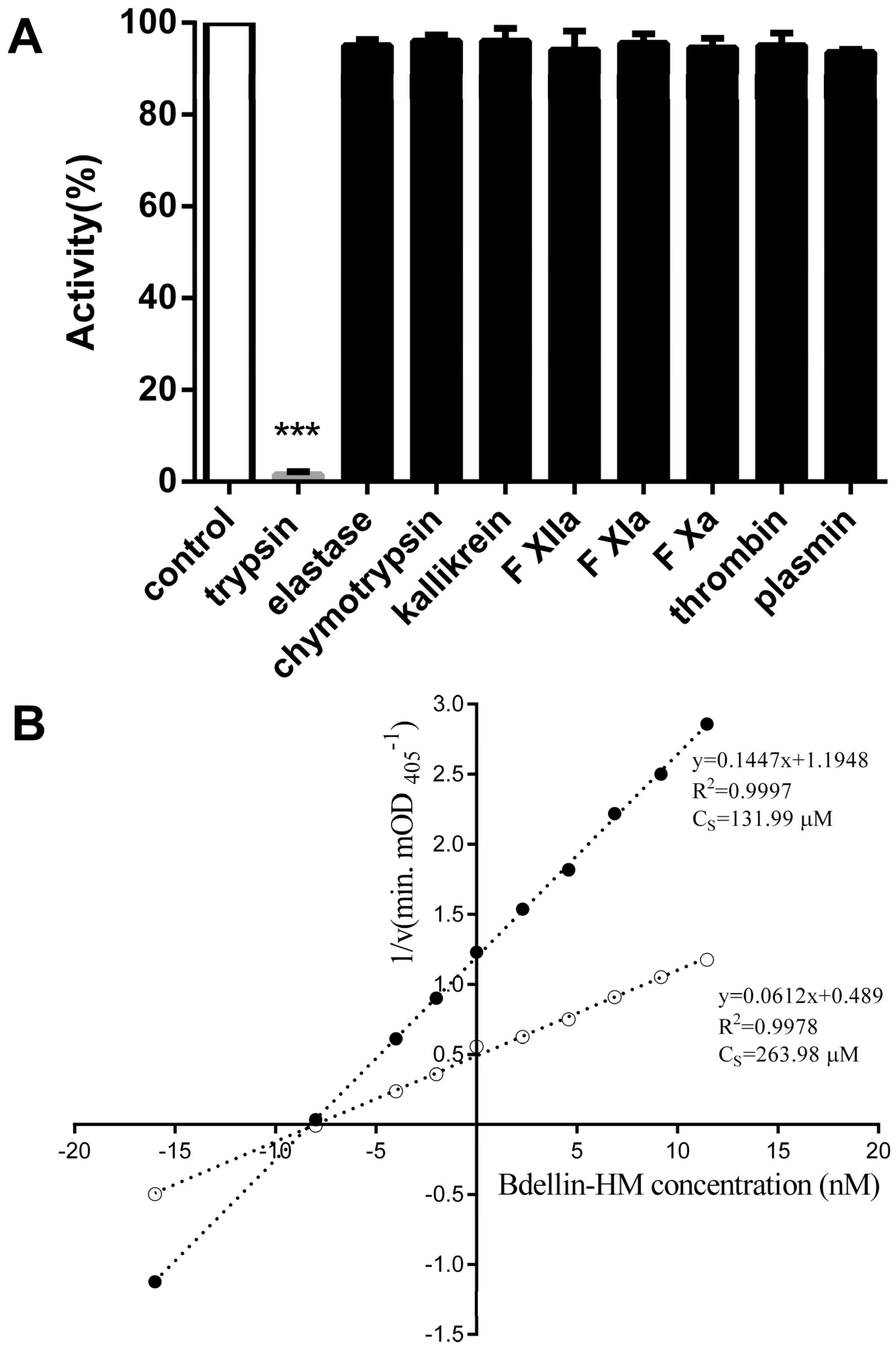
2.3. Protease Inhibitory Activity and Enzyme Kinetics
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Collection of Crude Extracts
5.2. Purification of Bdellin-HM
5.3. Mass Spectrometric Analysis and Sequencing of Peptide
5.4. RNA Extraction and cDNA Library Construction
5.5. Screening of cDNA Encoding Bdellin-HM
5.6. Protease Inhibitory Assays and Enzyme Kinetic Study
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Heutinck, K.M.; ten Berge, I.J.; Hack, C.E.; Hamann, J.; Rowshani, A.T. Serine proteases of the human immune system in health and disease. Mol. Immunol. 2010, 47, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Scott, C.J.; Taggart, C.C. Biologic protease inhibitors as novel therapeutic agents. Biochimie 2010, 92, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Sabotic, J.; Kos, J. Microbial and fungal protease inhibitors—Current and potential applications. Appl. Microbiol. Biotechnol. 2012, 93, 1351–1375. [Google Scholar] [CrossRef] [PubMed]
- Ashton-Rickardt, P.G. An emerging role for serine protease inhibitors in T lymphocyte immunity and beyond. Immunol. Lett. 2013, 152, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Shadrin, N.; Shapira, M.G.; Khalfin, B.; Uppalapati, L.; Parola, A.H.; Nathan, I. Serine protease inhibitors interact with IFN-gamma through up-regulation of FasR; a novel therapeutic strategy against cancer. Exp. Cell Res. 2015, 330, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Stapels, D.A.; Geisbrecht, B.V.; Rooijakkers, S.H. Neutrophil serine proteases in antibacterial defense. Curr. Opin. Microbiol. 2015, 23, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.I.; Mota, J.; Monteiro, S.A.; Ferreira, R.M. Legume seeds and colorectal cancer revisited: Protease inhibitors reduce MMP-9 activity and colon cancer cell migration. Food Chem. 2016, 197, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Lecaille, F.; Lalmanach, G.; Andrault, P.M. Antimicrobial proteins and peptides in human lung diseases: A friend and foe partnership with host proteases. Biochimie 2016, 122, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.; Abbenante, G.; Fairlie, D.P. Protease inhibitors: Current status and future prospects. J. Med. Chem. 2000, 43, 305–341. [Google Scholar] [CrossRef] [PubMed]
- Kanost, M.R. Serine proteinase inhibitors in arthropod immunity. Dev. Comp. Immunol. 1999, 23, 291–301. [Google Scholar] [CrossRef]
- Kazal, L.A.; Spicer, D.S.; Brahinsky, R.A. Isolation of a crystalline trypsin inhibitor-anticoagulant protein from pancreas. J. Am. Chem. Soc. 1948, 70, 3034–3040. [Google Scholar] [CrossRef] [PubMed]
- Rimphanitchayakit, V.; Tassanakajon, A. Structure and function of invertebrate Kazal-type serine proteinase inhibitors. Dev. Comp. Immunol. 2010, 34, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Pariani, S.; Contreras, M.; Rossi, F.R.; Sander, V.; Corigliano, M.G.; Simon, F.; Busi, M.V.; Gomez-Casati, D.F.; Pieckenstain, F.L.; Duschak, V.G.; et al. Characterization of a novel Kazal-type serine proteinase inhibitor of Arabidopsis thaliana. Biochimie 2016, 123, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M., Jr.; Kato, I. Protein inhibitors of proteinases. Annu. Rev. Biochem. 1980, 49, 593–626. [Google Scholar] [CrossRef] [PubMed]
- Kreutzmann, P.; Schulz, A.; Standker, L.; Forssmann, W.G.; Magert, H.J. Recombinant production, purification and biochemical characterization of domain 6 of lekti: A temporary Kazal-type-related serine proteinase inhibitor. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 803, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Magert, H.J.; Standker, L.; Kreutzmann, P.; Zucht, H.D.; Reinecke, M.; Sommerhoff, C.P.; Fritz, H.; Forssmann, W.G. Lekti, a novel 15-domain type of human serine proteinase inhibitor. J. Biol. Chem. 1999, 274, 21499–21502. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Huitema, E.; Da Cunha, L.; Torto-Alalibo, T.; Kamoun, S. A Kazal-like extracellular serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis-related protease P69B. J. Biol. Chem. 2004, 279, 26370–26377. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrot, C.; Ultsch, M.; Lipari, M.T.; Moran, P.; Lin, S.J.; Ganesan, R.; Quan, C.; Tom, J.; Sandoval, W.; van Lookeren Campagne, M.; et al. Structural and functional analysis of HtrA1 and its subdomains. Structure 2012, 20, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Derache, C.; Epinette, C.; Roussel, A.; Gabant, G.; Cadene, M.; Korkmaz, B.; Gauthier, F.; Kellenberger, C. Crystal structure of greglin, a novel non-classical Kazal inhibitor, in complex with subtilisin. FEBS J. 2012, 279, 4466–4478. [Google Scholar] [CrossRef] [PubMed]
- Li, X.C.; Wang, X.W.; Wang, Z.H.; Zhao, X.F.; Wang, J.X. A three-domain Kazal-type serine proteinase inhibitor exhibiting domain inhibitory and bacteriostatic activities from freshwater crayfish Procambarus clarkii. Dev. Comp. Immunol. 2009, 33, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Baskova, I.P.; Zavalova, L.L. Proteinase inhibitors from the medicinal leech Hirudo medicinalis. Biochem. Biokhimiia 2001, 66, 703–714. [Google Scholar] [CrossRef]
- Jung, H.I.; Kim, S.I.; Ha, K.S.; Joe, C.O.; Kang, K.W. Isolation and characterization of guamerin, a new human leukocyte elastase inhibitor from Hirudo nipponia. J. Biol. Chem. 1995, 270, 13879–13884. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Choi, J.G.; Lee, G.M.; Kang, K.W. Domain and genomic sequence analysis of bdellin-KL, a leech-derived trypsin-plasmin inhibitor. J. Biochem. 2001, 130, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Krejci, K.; Fritz, H. Structural homology of a trypsin-plasmin inhibitor from leeches (bdelin B-3) with secretory trypsin inhibitors from mammals. FEBS Lett. 1976, 64, 152–155. [Google Scholar] [CrossRef]
- Fritz, H.; Oppitz, K.H.; Gebhardt, M.; Oppitz, I.; Werle, E.; Marx, R. On the presence of a trypsin-plasmin inhibitor in hirudin. Hoppe-Seyler's Z. Physiol. Chem. 1969, 350, 91–92. [Google Scholar] [PubMed]
- Fritz, H.; Krejci, K. Trypsin-plasmin inhibitors (Bdellins) from leeches. Methods Enzymol. 1976, 45, 797–806. [Google Scholar] [PubMed]
- Electricwala, A.; Hartwell, R.; Scawen, M.D.; Atkinson, T. The complete amino acid sequence of a hirudin variant from the leech Hirudinaria manillensis. J. Protein Chem. 1993, 12, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Tschesche, H.; Kolkenbrock, H.; Bode, W. The covalent structure of the elastase inhibitor from Anemonia sulcata—A “non-classical” Kazal-type protein. Biol. Chem. Hoppe-Seyler 1987, 368, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Fink, E.; Rehm, H.; Gippner, C.; Bode, W.; Eulitz, M.; Machleidt, W.; Fritz, H. The primary structure of bdellin B-3 from the leech Hirudo medicinalis. Bdellin B-3 is a compact proteinase inhibitor of a “non-classical” kazal type. It is present in the leech in a high molecular mass form. Biol. Chem. Hoppe-Seyler 1986, 367, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.S.; Mishra, M.; Suresh, C.G.; Gupta, V.S.; Giri, A.P. Complementation of intramolecular interactions for structural-functional stability of plant serine proteinase inhibitors. Biochim. Biophys. Acta 2013, 1830, 5087–5094. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Song, L.; Chang, Y.; Xu, W.; Wu, L. Molecular cloning, characterization and expression of a novel serine proteinase inhibitor gene in bay scallops (Argopecten irradians, lamarck 1819). Fish Shellfish Immunol. 2006, 20, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Schechter, I.; Berger, A. On the size of the active site in proteases. I. Papain. Biochem. Biophys. Res. Commun. 1967, 27, 157–162. [Google Scholar] [CrossRef]
- Scacheri, E.; Nitti, G.; Valsasina, B.; Orsini, G.; Visco, C.; Ferrera, M.; Sawyer, R.T.; Sarmientos, P. Novel hirudin variants from the leech Hirudinaria manillensis. Amino acid sequence, cDNA cloning and genomic organization. Eur. J. Biochem. FEBS 1993, 214, 295–304. [Google Scholar] [CrossRef]
- Hao, X.; Tang, X.; Luo, L.; Wang, Y.; Lai, R.; Lu, Q. A novel ranacyclin-like peptide with anti-platelet activity identified from skin secretions of the frog Amolops loloensis. Gene 2016, 576, 171–175. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| 1 | 5′-GAYWSNGARTGYGTNTGYAC-3′ |
| 2 | 5′-AAGCAGTGGTATCAACGCAGAGT-3′ |
| 3 | 5′-CTYACRCANACRTGNTTY-3′ |
| 4 | 5′-ATTCTAGAGGCCGAGGCGGCCGA-3′ |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, Y.; Li, B.; Liu, W.; Wang, G.; Du, C.; Ombati, R.; Lai, R.; Long, C.; Li, H. Purification and Characterization of a Novel Kazal-Type Trypsin Inhibitor from the Leech of Hirudinaria manillensis. Toxins 2016, 8, 229. https://doi.org/10.3390/toxins8080229
Lai Y, Li B, Liu W, Wang G, Du C, Ombati R, Lai R, Long C, Li H. Purification and Characterization of a Novel Kazal-Type Trypsin Inhibitor from the Leech of Hirudinaria manillensis. Toxins. 2016; 8(8):229. https://doi.org/10.3390/toxins8080229
Chicago/Turabian StyleLai, Yanmei, Bowen Li, Weihui Liu, Gan Wang, Canwei Du, Rose Ombati, Ren Lai, Chengbo Long, and Hongyuan Li. 2016. "Purification and Characterization of a Novel Kazal-Type Trypsin Inhibitor from the Leech of Hirudinaria manillensis" Toxins 8, no. 8: 229. https://doi.org/10.3390/toxins8080229
APA StyleLai, Y., Li, B., Liu, W., Wang, G., Du, C., Ombati, R., Lai, R., Long, C., & Li, H. (2016). Purification and Characterization of a Novel Kazal-Type Trypsin Inhibitor from the Leech of Hirudinaria manillensis. Toxins, 8(8), 229. https://doi.org/10.3390/toxins8080229