
Distinct Neurotoxicity Profile of Listeriolysin O from Listeria monocytogenes

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
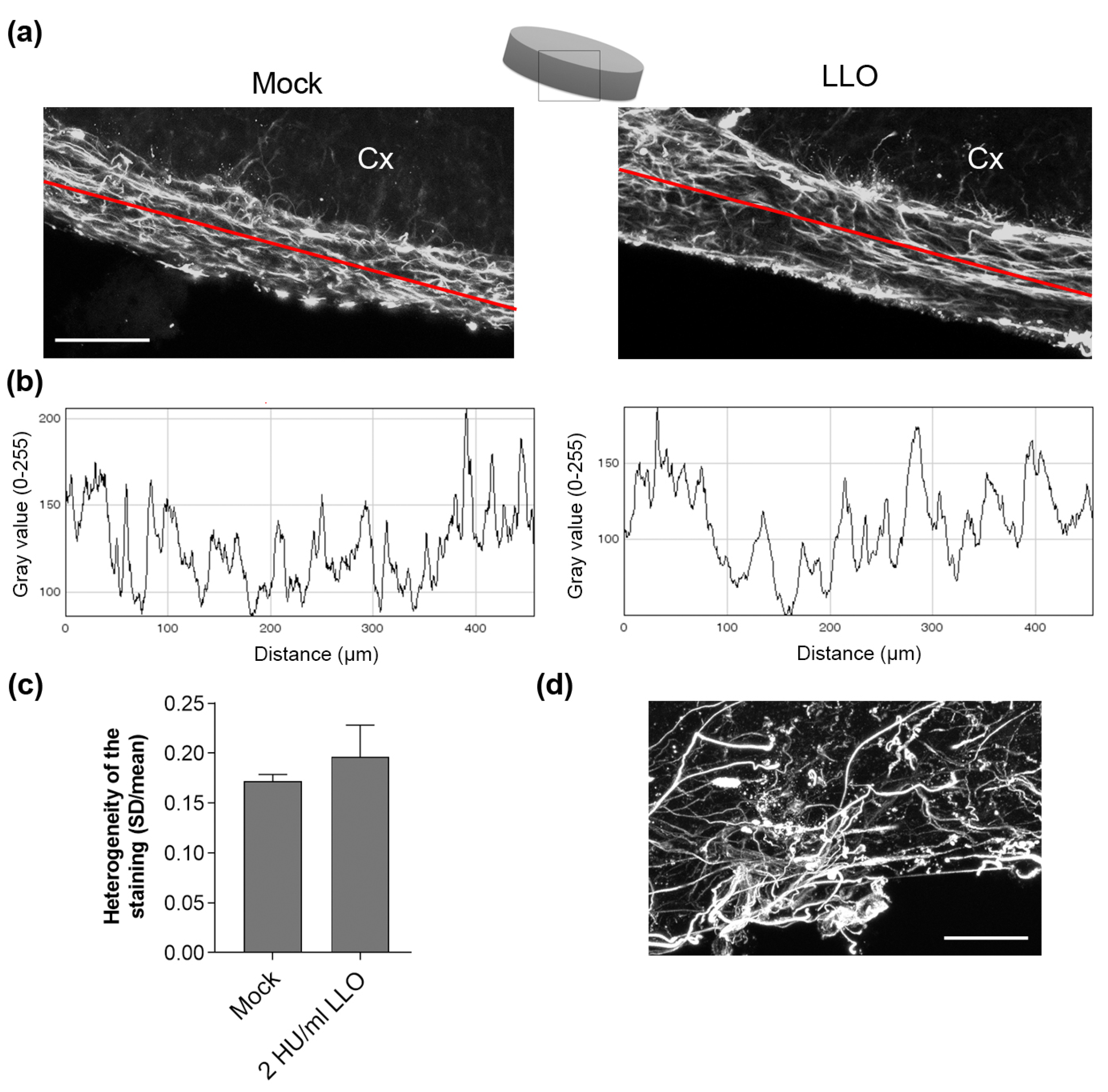
2. Results
3. Discussion
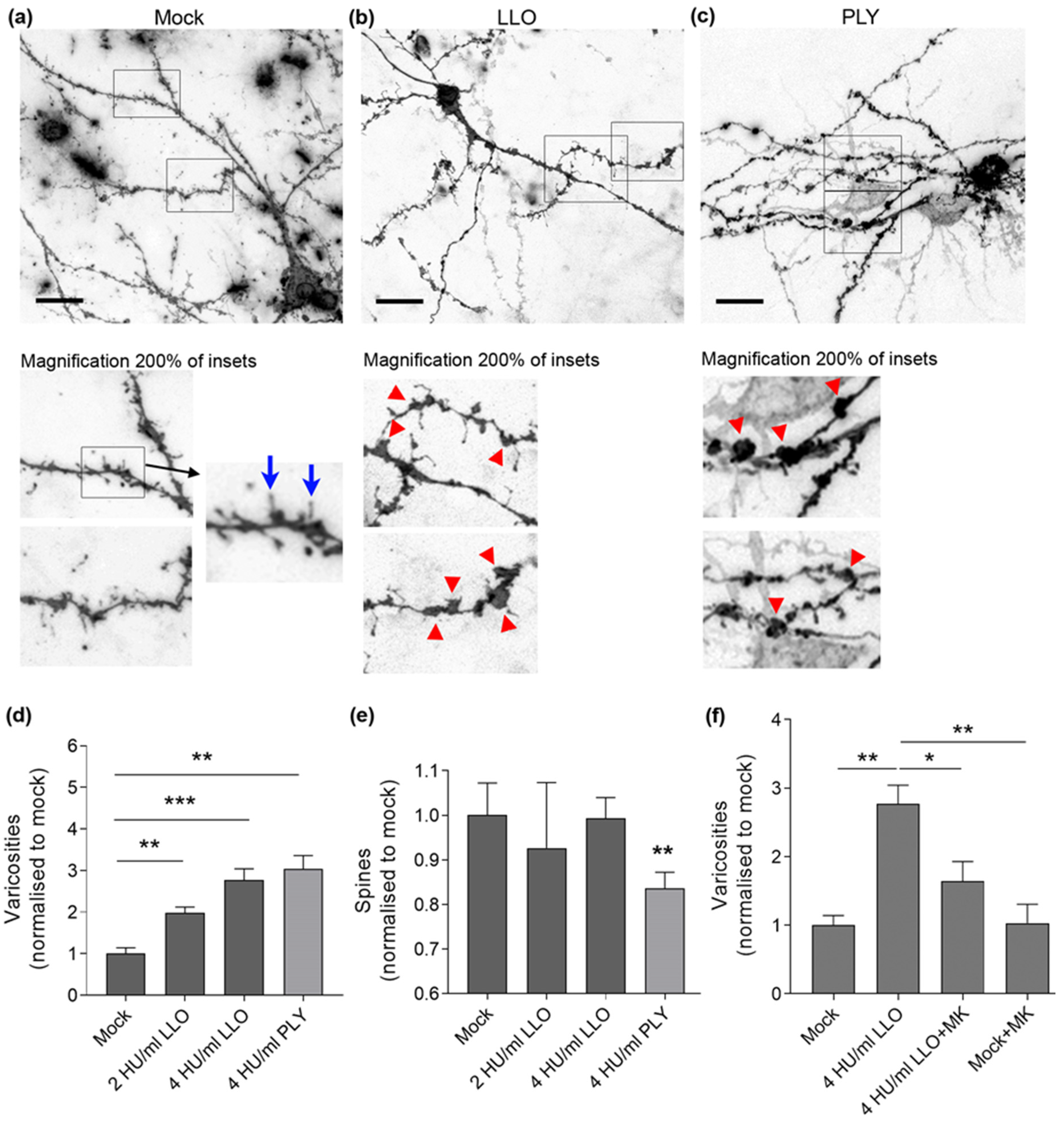
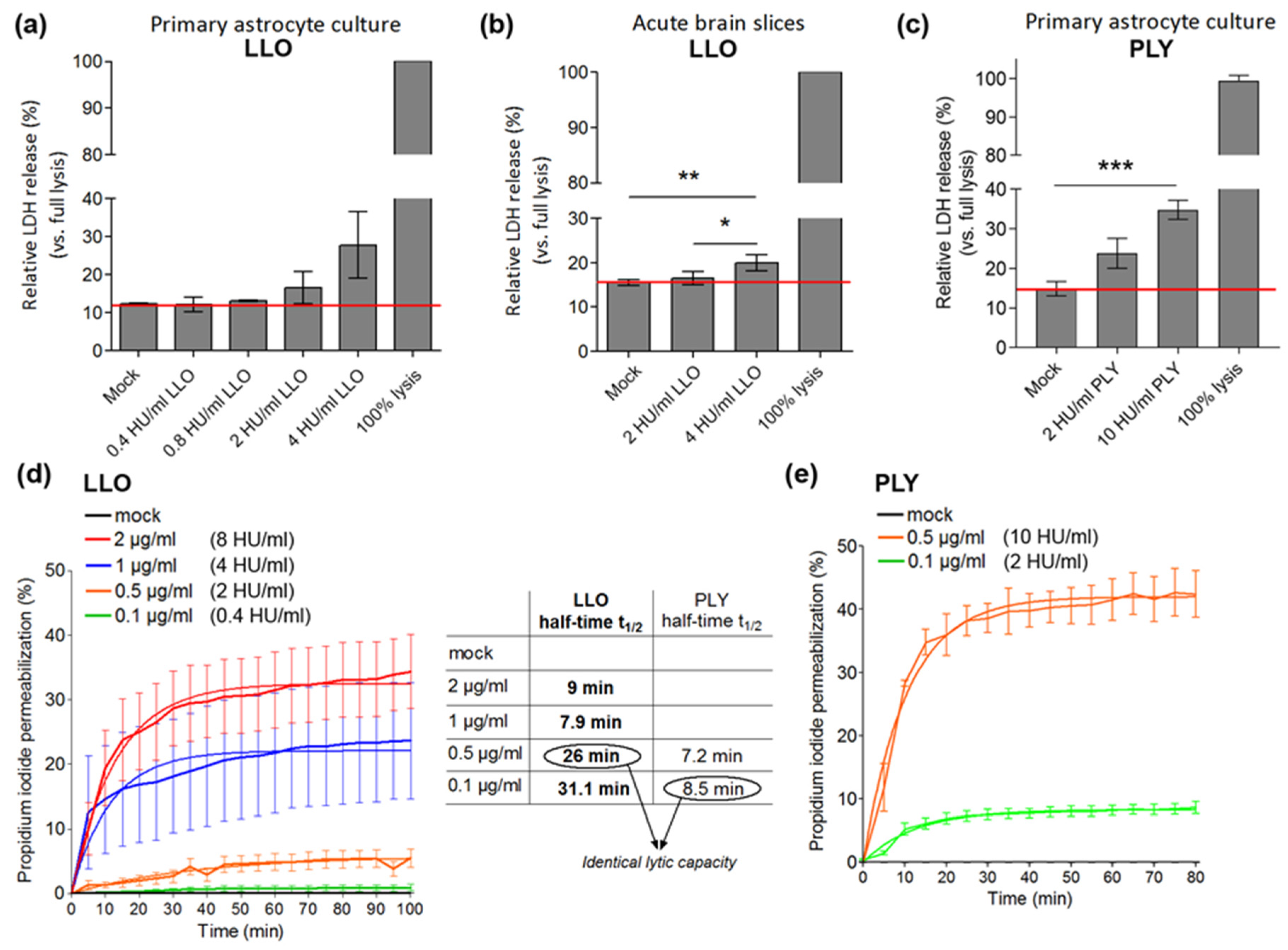
- Dendritic varicosity formation (swelling of the dendrites that are mostly NMDA glutamate receptor-dependent). A similar phenomenon is observed in ischemic brain damage [25]. Studies have described the presence of dendritic varicosities as an element of neural damage in models of diabetes and prion disease [26,27].
- Dendritic spine collapse, which represents a morphological substrate of synaptic loss [28]. Dendritic spines represent actin-filled dendritic protrusions, which are very dynamic at the beginning of their formation. A synapse is formed on the top of a dendritic spine, in contact with neighboring neurites. The spine stabilizes and becomes mature [29].
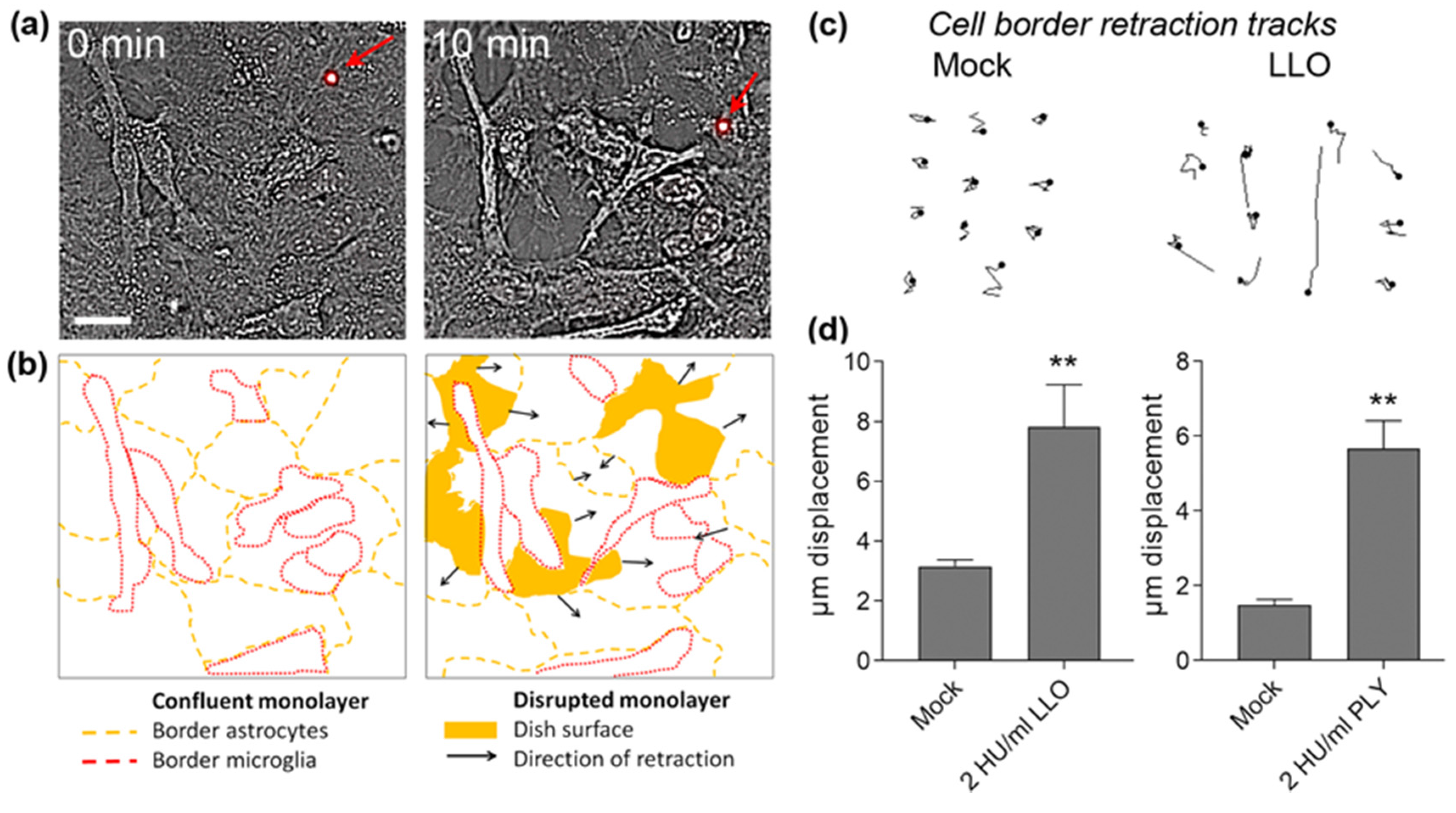
- the magnitude and mechanical strength of cell shape changes differ from toxin to toxin;
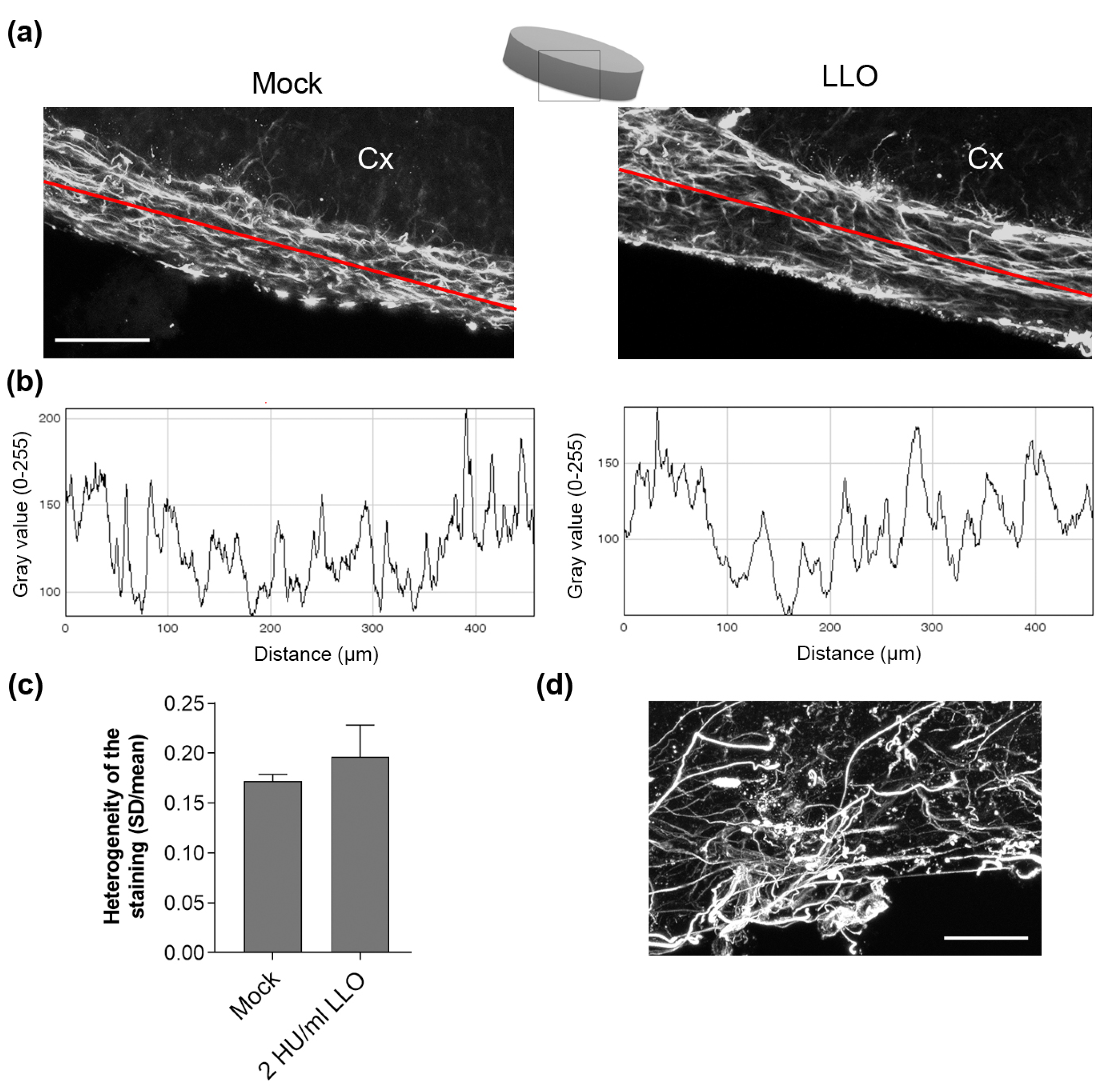
- the time frame of cell shape changes by LLO may be slower compared to PLY, thus allowing more effective tissue adaptation;
- continuous exposure of tissue to LLO in normal pH conditions may lead to toxin inactivation; or
- inhibitory cellular metabolites specifically inhibit LLO.
4. Materials and Methods
4.1. LLO and PLY Preparation
4.2. Cell and Slice Cultures and Culture Treatments
4.3. Live Imaging Experiments
4.4. Lactate Dehydrogenase (LDH) Test
4.5. Neural Staining in Slices and Immunohistochemistry
4.6. Microscopy
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dal Peraro, M.; van der Goot, F.G. Pore-forming toxins: Ancient, but never really out of fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Hotze, E.M.; Tweten, R.K. Membrane assembly of the cholesterol-dependent cytolysin pore complex. Biochim. Biophys. Acta 2012, 1818, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- Rosado, C.J.; Kondos, S.; Bull, T.E.; Kuiper, M.J.; Law, R.H.; Buckle, A.M.; Voskoboinik, I.; Bird, P.I.; Trapani, J.A.; Whisstock, J.C.; et al. The MACPF/CDC family of pore-forming toxins. Cell Microbiol. 2008, 10, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Brooks, N.J.; Smith, R.A.; Lea, S.M.; Bubeck, D. Structural basis for recognition of the pore-forming toxin intermedilysin by human complement receptor CD59. Cell Rep. 2013, 3, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Bavdek, A.; Gekara, N.O.; Priselac, D.; Gutierrez Aguirre, I.; Darji, A.; Chakraborty, T.; Macek, P.; Lakey, J.H.; Weiss, S.; Anderluh, G. Sterol and pH interdependence in the binding, oligomerization, and pore formation of listeriolysin O. Biochemistry 2007, 46, 4425–4437. [Google Scholar] [CrossRef] [PubMed]
- Marriott, H.M.; Mitchell, T.J.; Dockrell, D.H. Pneumolysin: A double-edged sword during the host-pathogen interaction. Curr. Mol. Med. 2008, 8, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.M.; Ellemor, D.M.; Boyd, R.L.; Emmins, J.J.; Rood, J.I. Synergistic effects of alpha-toxin and perfringolysin O in clostridium perfringens-mediated gas gangrene. Infect. Immun. 2001, 69, 7904–7910. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.L.; Calderwood, S.B.; Weber, D.J.; Miller, S.I.; Southwick, F.S.; Caviness, V.S., Jr.; Swartz, M.N. Acute bacterial meningitis in adults. A review of 493 episodes. N. Engl. J. Med. 1993, 328, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Meli, D.N.; Christen, S.; Leib, S.L.; Tauber, M.G. Current concepts in the pathogenesis of meningitis caused by streptococcus pneumoniae. Curr. Opin. Infect. Dis. 2002, 15, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Hirst, R.A.; Gosai, B.; Rutman, A.; Guerin, C.J.; Nicotera, P.; Andrew, P.W.; O’Callaghan, C. Streptococcus pneumoniae deficient in pneumolysin or autolysin has reduced virulence in meningitis. J. Infect. Dis. 2008, 197, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Wippel, C.; Maurer, J.; Förtsch, C.; Hupp, S.; Bohl, A.; Ma, J.; Mitchell, T.J.; Bunkowski, S.; Brück, W.; Nau, R.; et al. Bacterial cytolysin during meningitis disrupts the regulation of glutamate in the brain, leading to synaptic damage. PLoS Pathog. 2013, 9, e1003380. [Google Scholar] [CrossRef] [PubMed]
- Hupp, S.; Heimeroth, V.; Wippel, C.; Fortsch, C.; Ma, J.; Mitchell, T.J.; Iliev, A.I. Astrocytic tissue remodeling by the meningitis neurotoxin pneumolysin facilitates pathogen tissue penetration and produces interstitial brain edema. Glia 2012, 60, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Förtsch, C.; Hupp, S.; Ma, J.; Mitchell, T.J.; Maier, E.; Benz, R.; Iliev, A.I. Changes in astrocyte shape induced by sublytic concentrations of the cholesterol-dependent cytolysin pneumolysin still require pore-forming capacity. Toxins 2011, 3, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Stringaris, A.K.; Geisenhainer, J.; Bergmann, F.; Balshusemann, C.; Lee, U.; Zysk, G.; Mitchell, T.J.; Keller, B.U.; Kuhnt, U.; Gerber, J.; et al. Neurotoxicity of pneumolysin, a major pneumococcal virulence factor, involves calcium influx and depends on activation of p38 mitogen-activated protein kinase. Neurobiol. Dis. 2002, 11, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Ebert, S.; Gerber, J.; Bader, S.; Muhlhauser, F.; Brechtel, K.; Mitchell, T.J.; Nau, R. Dose-dependent activation of microglial cells by toll-like receptor agonists alone and in combination. J. Neuroimmunol. 2005, 159, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Althaus, D.; Lehner, A.; Brisse, S.; Maury, M.; Tasara, T.; Stephan, R. Characterization of Listeria monocytogenes strains isolated during 2011–2013 from human infections in switzerland. Foodborne Pathog. Dis. 2014, 11, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Barbuddhe, S.B.; Chakraborty, T. Listeria as an enteroinvasive gastrointestinal pathogen. Curr. Top. Microbiol. Immunol. 2009, 337, 173–195. [Google Scholar] [PubMed]
- Hamon, M.A.; Ribet, D.; Stavru, F.; Cossart, P. Listeriolysin O: The swiss army knife of Listeria. Trends Microbiol. 2012, 20, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Seveau, S. Multifaceted activity of listeriolysin O, the cholesterol-dependent cytolysin of Listeria monocytogenes. Subcell. Biochem. 2014, 80, 161–195. [Google Scholar] [PubMed]
- Wippel, C.; Fortsch, C.; Hupp, S.; Maier, E.; Benz, R.; Ma, J.; Mitchell, T.J.; Iliev, A.I. Extracellular calcium reduction strongly increases the lytic capacity of pneumolysin from streptococcus pneumoniae in brain tissue. J. Infect. Dis. 2011, 204, 930–936. [Google Scholar] [CrossRef] [PubMed]
- Glomski, I.J.; Decatur, A.L.; Portnoy, D.A. Listeria monocytogenes mutants that fail to compartmentalize listerolysin O activity are cytotoxic, avirulent, and unable to evade host extracellular defenses. Infect. Immun. 2003, 71, 6754–6765. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.F.; Gitter, A.H.; Gunzel, D.; Weiss, S.; Mohamed, W.; Chakraborty, T.; Fromm, M.; Schulzke, J.D. Listeriolysin O affects barrier function and induces chloride secretion in HT-29/B6 colon epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1350–G1359. [Google Scholar] [CrossRef] [PubMed]
- Schuerch, D.W.; Wilson-Kubalek, E.M.; Tweten, R.K. Molecular basis of listeriolysin O pH dependence. Proc. Natl. Acad. Sci. USA 2005, 102, 12537–12542. [Google Scholar] [CrossRef] [PubMed]
- Spreer, A.; Kerstan, H.; Bottcher, T.; Gerber, J.; Siemer, A.; Zysk, G.; Mitchell, T.J.; Eiffert, H.; Nau, R. Reduced release of pneumolysin by streptococcus pneumoniae in vitro and in vivo after treatment with nonbacteriolytic antibiotics in comparison to ceftriaxone. Antimicrob. Agents Chemother. 2003, 47, 2649–2654. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, S.M.; Connolly, C.N. Dendritic and mitochondrial changes during glutamate excitotoxicity. Neuropharmacology 2007, 53, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Luo, Y.; Dai, J. Axonal and dendritic changes are associated with diabetic encephalopathy in rats: An important risk factor for alzheimer’s disease. J. Alzheimer's Dis. 2013, 34, 937–947. [Google Scholar]
- Fuhrmann, M.; Mitteregger, G.; Kretzschmar, H.; Herms, J. Dendritic pathology in prion disease starts at the synaptic spine. J. Neurosci. 2007, 27, 6224–6233. [Google Scholar] [CrossRef] [PubMed]
- Smart, F.M.; Halpain, S. Regulation of dendritic spine stability. Hippocampus 2000, 10, 542–554. [Google Scholar] [CrossRef]
- Lippman, J.; Dunaevsky, A. Dendritic spine morphogenesis and plasticity. J. Neurobiol. 2005, 64, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Ikegaya, Y.; Kim, J.A.; Baba, M.; Iwatsubo, T.; Nishiyama, N.; Matsuki, N. Rapid and reversible changes in dendrite morphology and synaptic efficacy following NMDA receptor activation: Implication for a cellular defense against excitotoxicity. J. Cell Sci. 2001, 114, 4083–4093. [Google Scholar] [PubMed]
- Velge, P.; Roche, S.M. Variability of Listeria monocytogenes virulence: A result of the evolution between saprophytism and virulence? Future Microbiol. 2010, 5, 1799–1821. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Hohmann, E.L.; Calderwood, S.B. Central nervous system infection with Listeria monocytogenes. 33 years’ experience at a general hospital and review of 776 episodes from the literature. Medicine 1998, 77, 313–336. [Google Scholar]
- Bojanowski, M.W.; Seizeur, R.; Effendi, K.; Bourgouin, P.; Magro, E.; Letourneau-Guillon, L. Spreading of multiple Listeria monocytogenes abscesses via central nervous system fiber tracts: Case report. J. Neurosurg. 2015, 123, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Zhang, C.; Dong, H.; Niu, Z.; Pan, Z.; Huang, J.; Jiao, X. Protective immunity induced by a LLO-deficient Listeria monocytogenes. Microbiol. Immunol. 2010, 54, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Schnupf, P.; Hofmann, J.; Norseen, J.; Glomski, I.J.; Schwartzstein, H.; Decatur, A.L. Regulated translation of listeriolysin O controls virulence of Listeria monocytogenes. Mol. Microbiol. 2006, 61, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Hupp, S.; Fortsch, C.; Wippel, C.; Ma, J.; Mitchell, T.J.; Iliev, A.I. Direct transmembrane interaction between actin and the pore-competent, cholesterol-dependent cytolysin pneumolysin. J. Mol. Biol. 2013, 425, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Iliev, A.I.; Djannatian, J.R.; Opazo, F.; Gerber, J.; Nau, R.; Mitchell, T.J.; Wouters, F.S. Rapid microtubule bundling and stabilization by the Streptococcus pneumoniae neurotoxin pneumolysin in a cholesterol-dependent, non-lytic and Src-kinase dependent manner inhibits intracellular trafficking. Mol. Microbiol. 2009, 71, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Iliev, A.I.; Djannatian, J.R.; Nau, R.; Mitchell, T.J.; Wouters, F.S. Cholesterol-dependent actin remodeling via RhoA and Rac1 activation by the Streptococcus pneumoniae toxin pneumolysin. Proc. Natl. Acad. Sci. USA 2007, 104, 2897–2902. [Google Scholar] [CrossRef] [PubMed]
- Schluter, D.; Domann, E.; Buck, C.; Hain, T.; Hof, H.; Chakraborty, T.; Deckert-Schluter, M. Phosphatidylcholine-specific phospholipase C from Listeria monocytogenes is an important virulence factor in murine cerebral listeriosis. Infect. Immun. 1998, 66, 5930–5938. [Google Scholar] [PubMed]
- Xiong, C.; Yang, G.; Kumar, S.; Aggarwal, S.; Leustik, M.; Snead, C.; Hamacher, J.; Fischer, B.; Umapathy, N.S.; Hossain, H.; et al. The lectin-like domain of TNF protects from listeriolysin-induced hyperpermeability in human pulmonary microvascular endothelial cells—A crucial role for protein kinase C-alpha inhibition. Vascul. Pharmacol. 2010, 52, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Douce, G.; Ross, K.; Cowan, G.; Ma, J.; Mitchell, T.J. Novel mucosal vaccines generated by genetic conjugation of heterologous proteins to pneumolysin (PLY) from Streptococcus pneumoniae. Vaccine 2010, 28, 3231–3237. [Google Scholar] [CrossRef] [PubMed]
- Klauer, S. The corticotectal projection of the rat established in organotypic culture. Neuroreport 1991, 2, 569–572. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurer, J.; Hupp, S.; Bischoff, C.; Foertsch, C.; Mitchell, T.J.; Chakraborty, T.; Iliev, A.I. Distinct Neurotoxicity Profile of Listeriolysin O from Listeria monocytogenes. Toxins 2017, 9, 34. https://doi.org/10.3390/toxins9010034
Maurer J, Hupp S, Bischoff C, Foertsch C, Mitchell TJ, Chakraborty T, Iliev AI. Distinct Neurotoxicity Profile of Listeriolysin O from Listeria monocytogenes. Toxins. 2017; 9(1):34. https://doi.org/10.3390/toxins9010034
Chicago/Turabian StyleMaurer, Jana, Sabrina Hupp, Carolin Bischoff, Christina Foertsch, Timothy J. Mitchell, Trinad Chakraborty, and Asparouh I. Iliev. 2017. "Distinct Neurotoxicity Profile of Listeriolysin O from Listeria monocytogenes" Toxins 9, no. 1: 34. https://doi.org/10.3390/toxins9010034