
The Preparation and Identification of a Monoclonal Antibody against Citrinin and the Development of Detection via Indirect Competitive ELISA

Abstract
:1. Introduction
2. Results
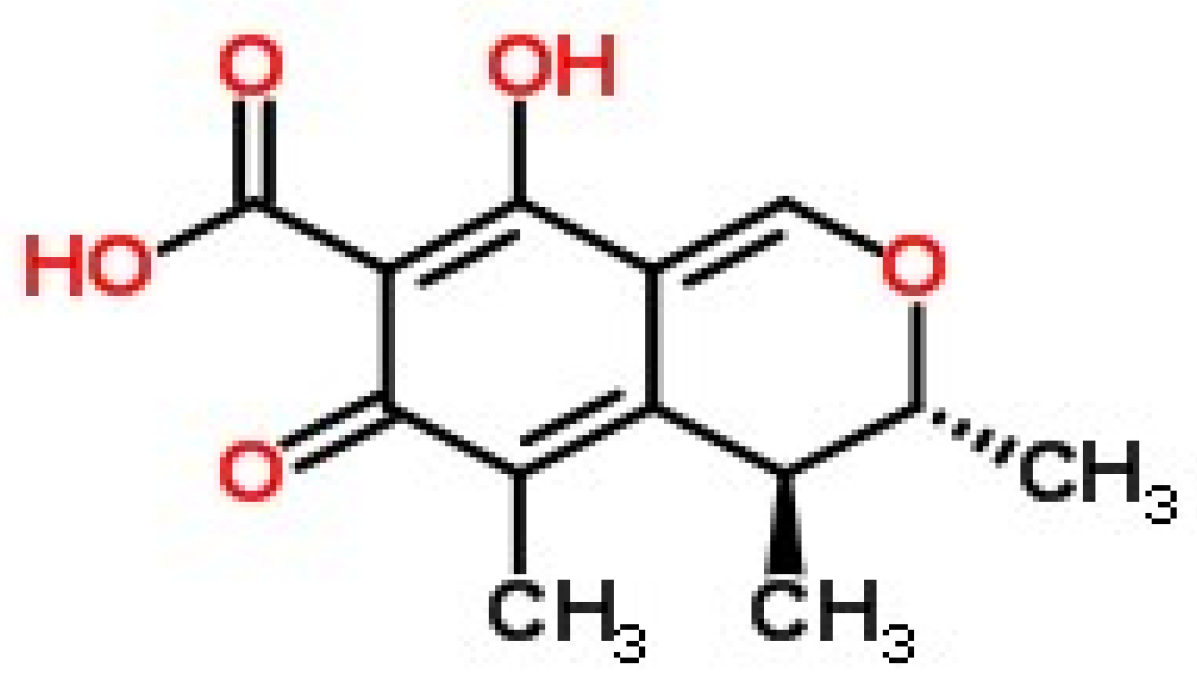
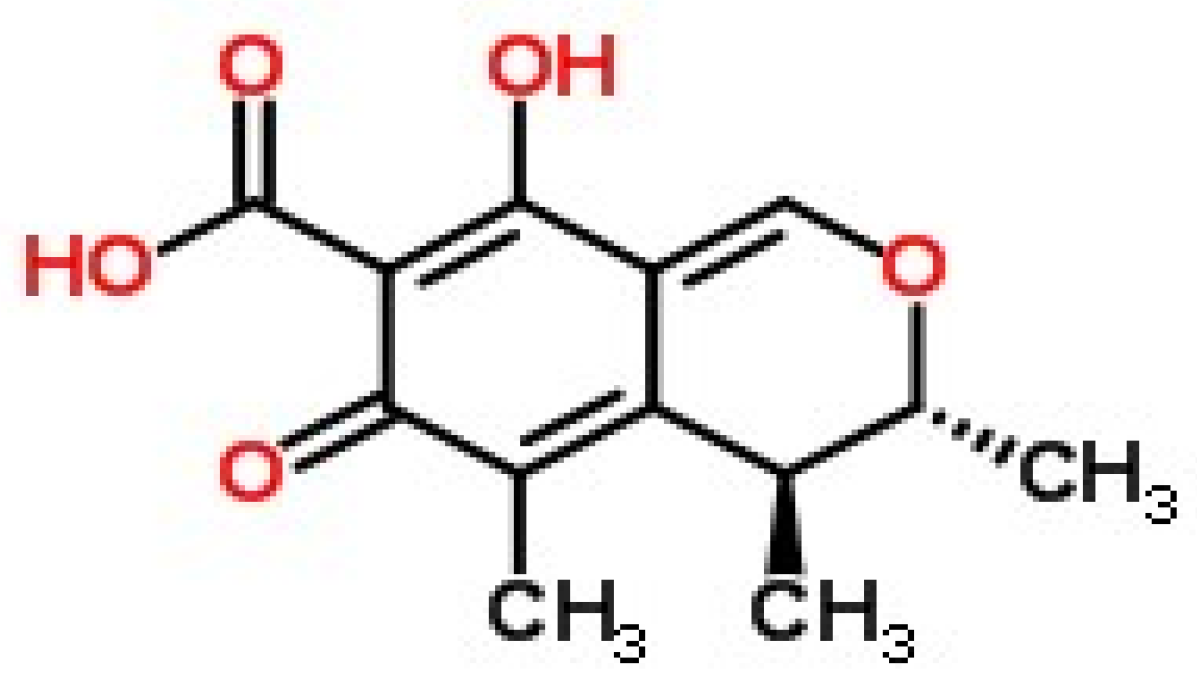
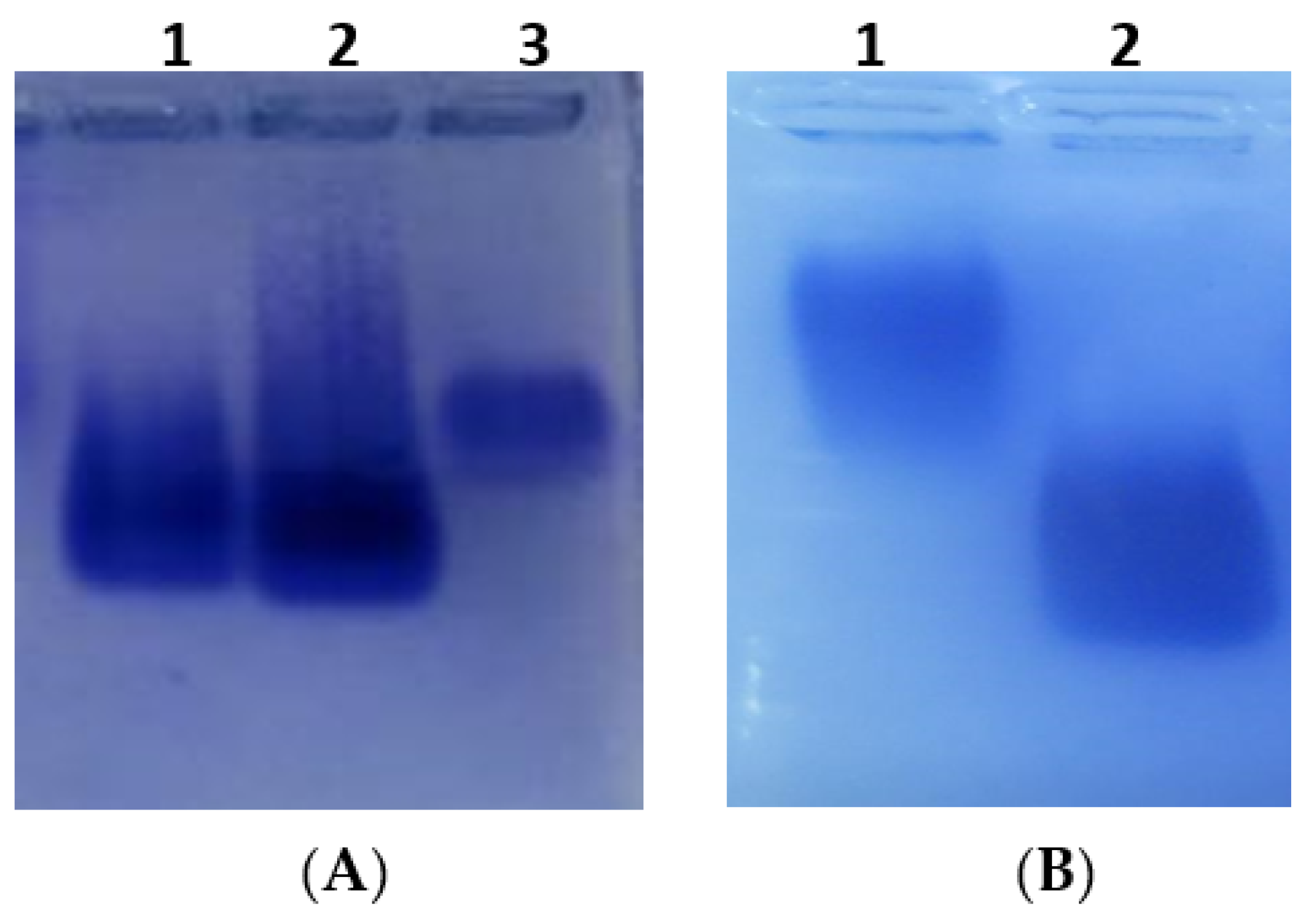
2.1. Synthesis and Identification of CTN-Protein Conjugates
2.2. Anti-Serum ELISA Assay of the Immunized Mice
2.3. Cell Fusion and Screening of Hybridoma Cells
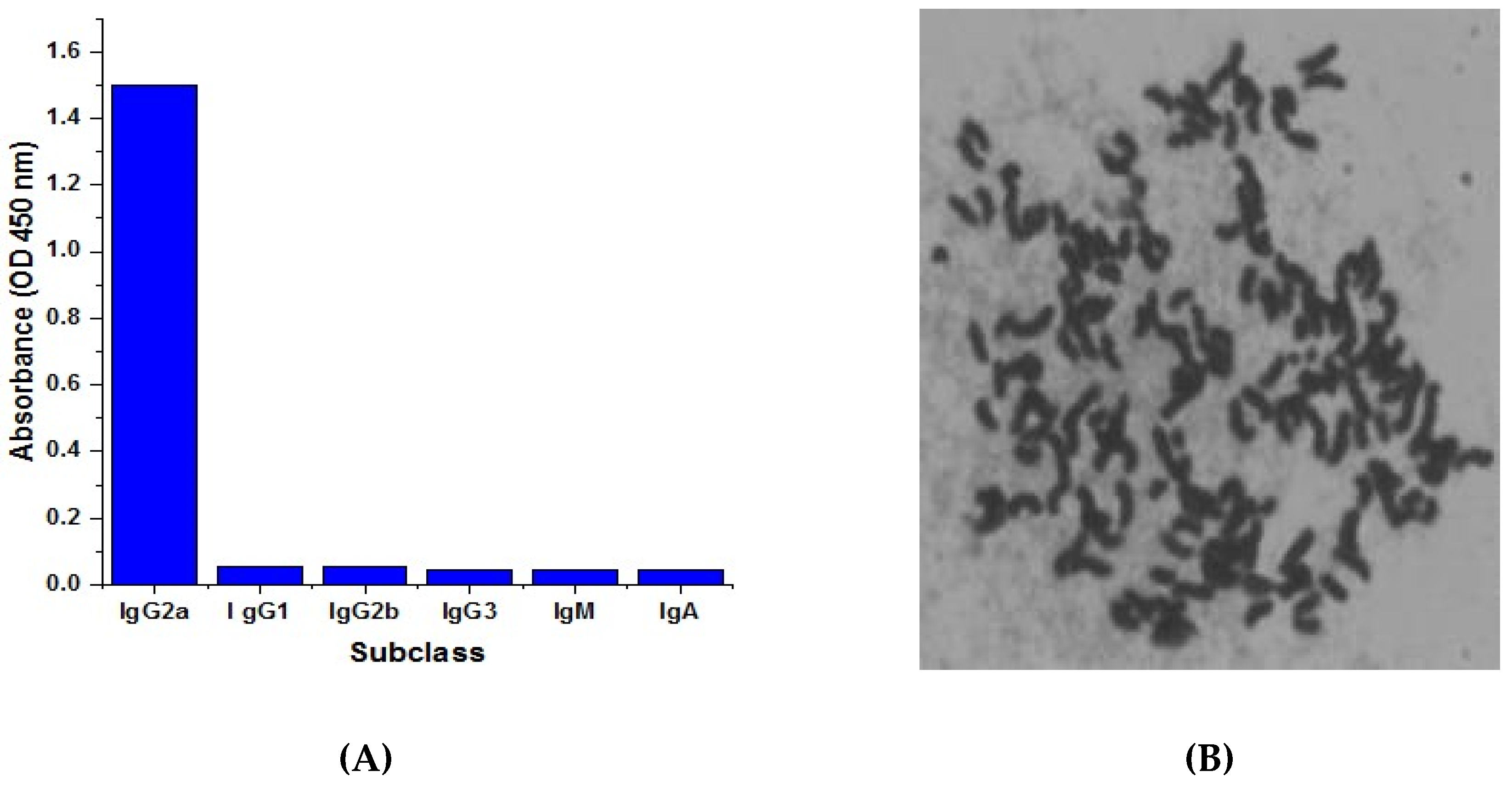
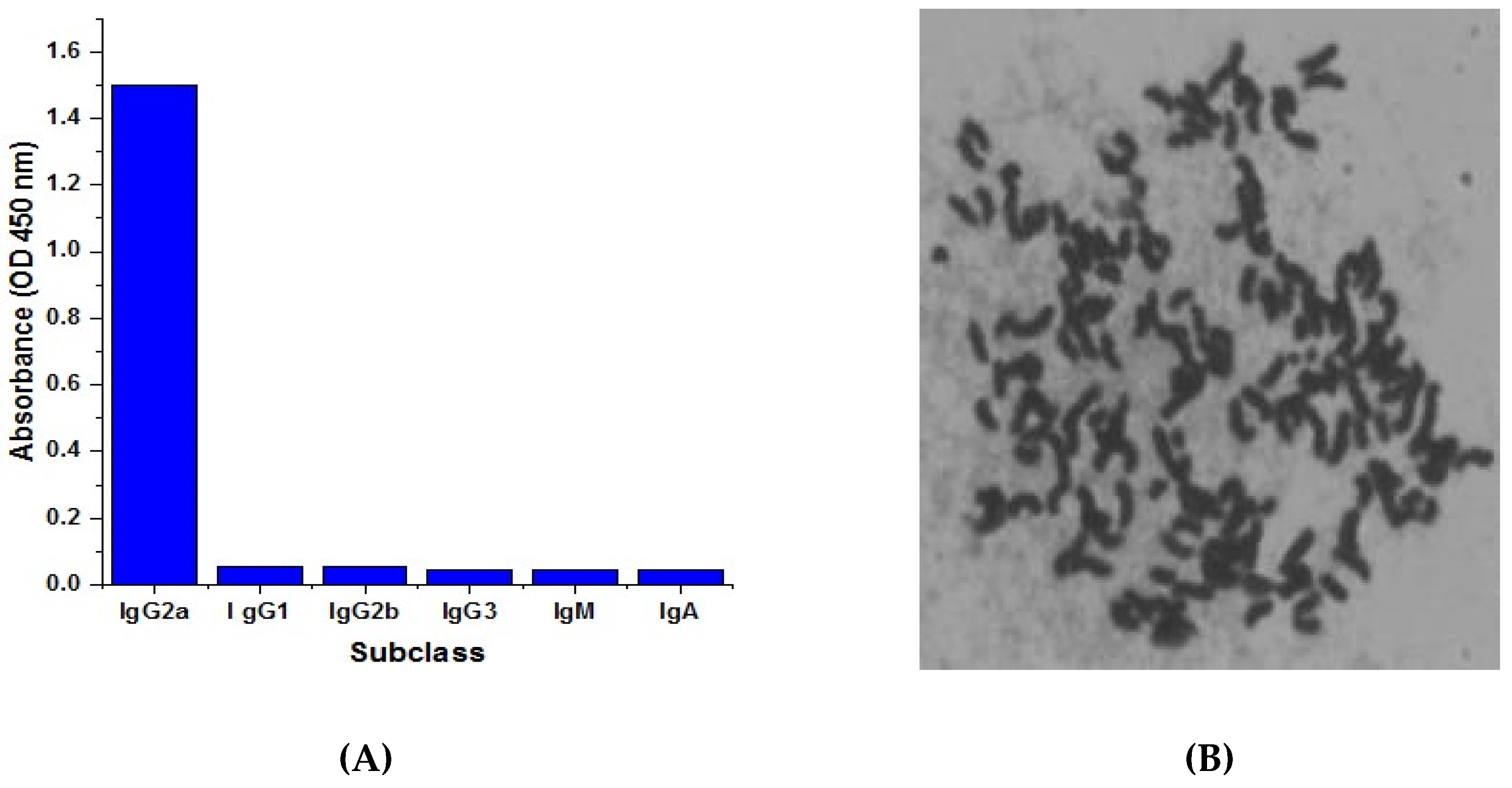
2.4. Isotyping and Chromosome Analysis of Anti-CTN mcAb
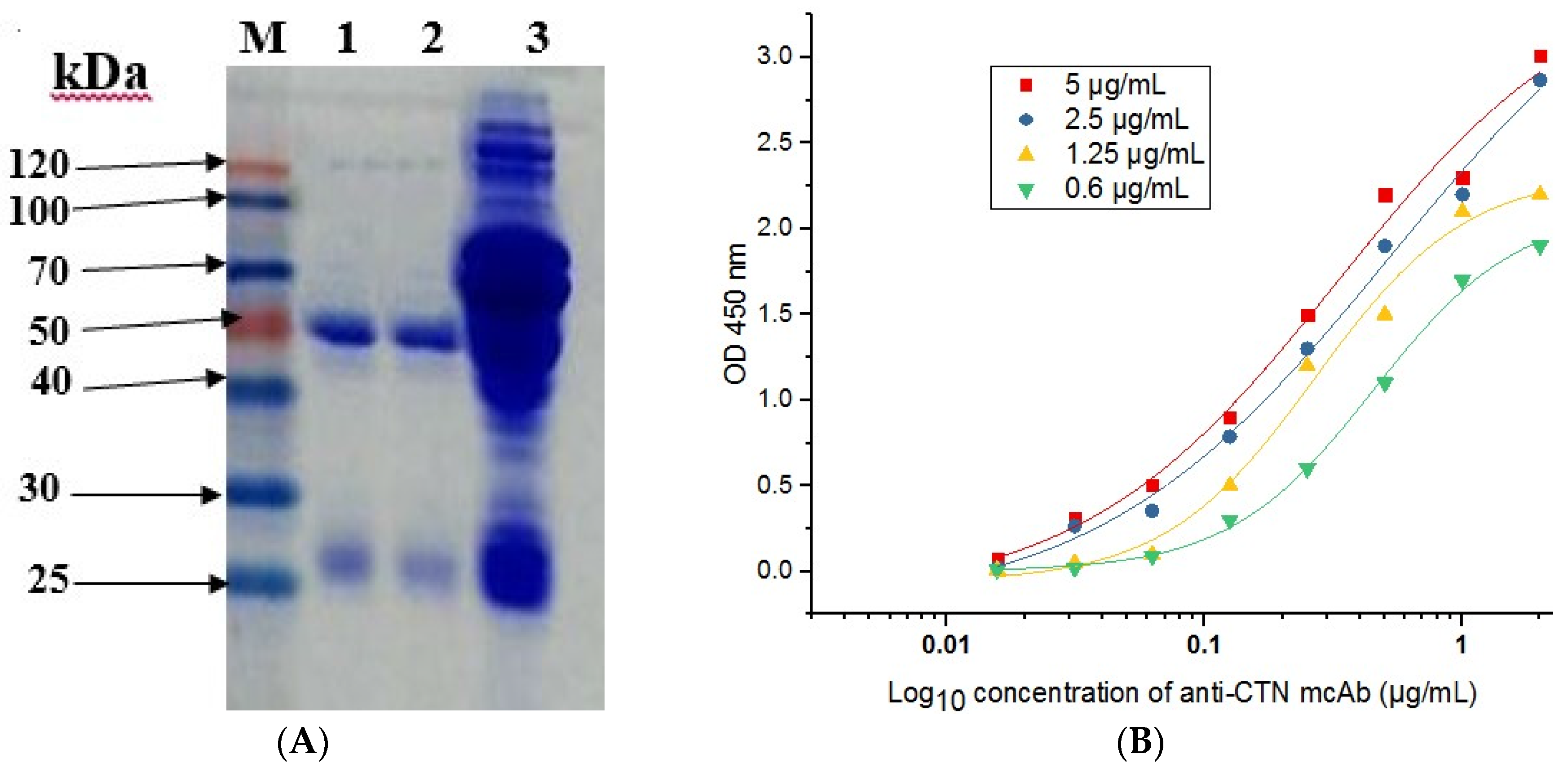
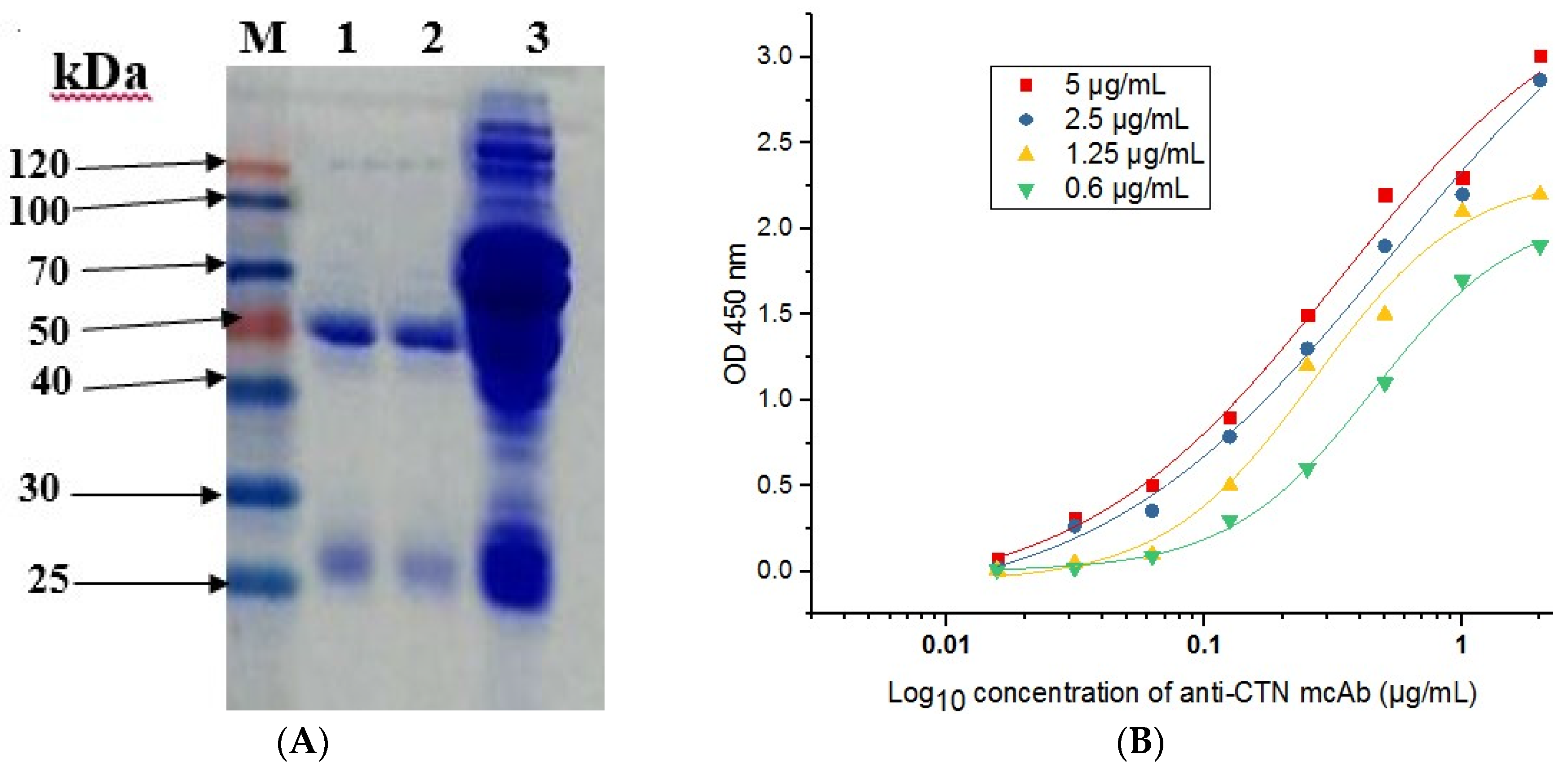
2.5. Purification of Anti-CTN mcAb
2.6. Affinity and Specificity Test of Anti-CTN mcAb
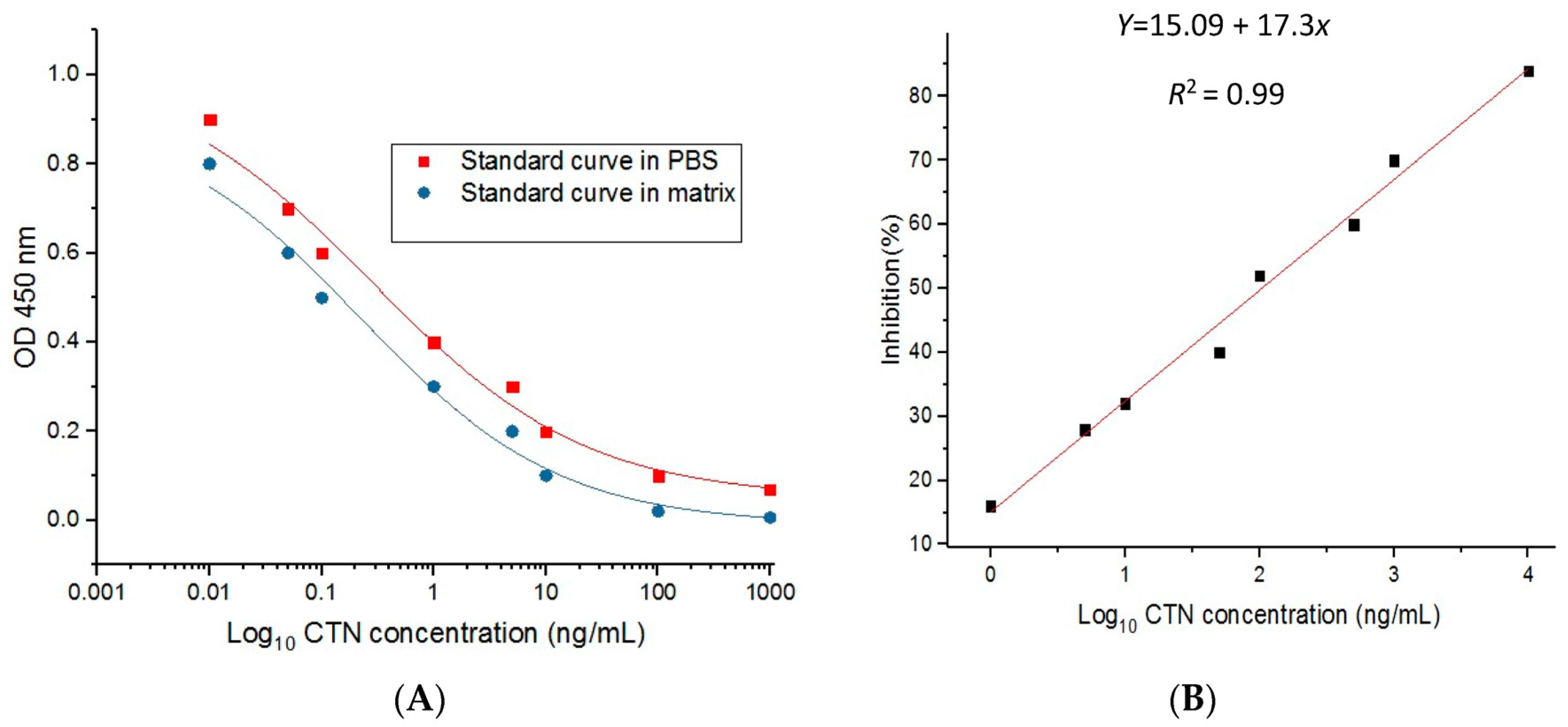
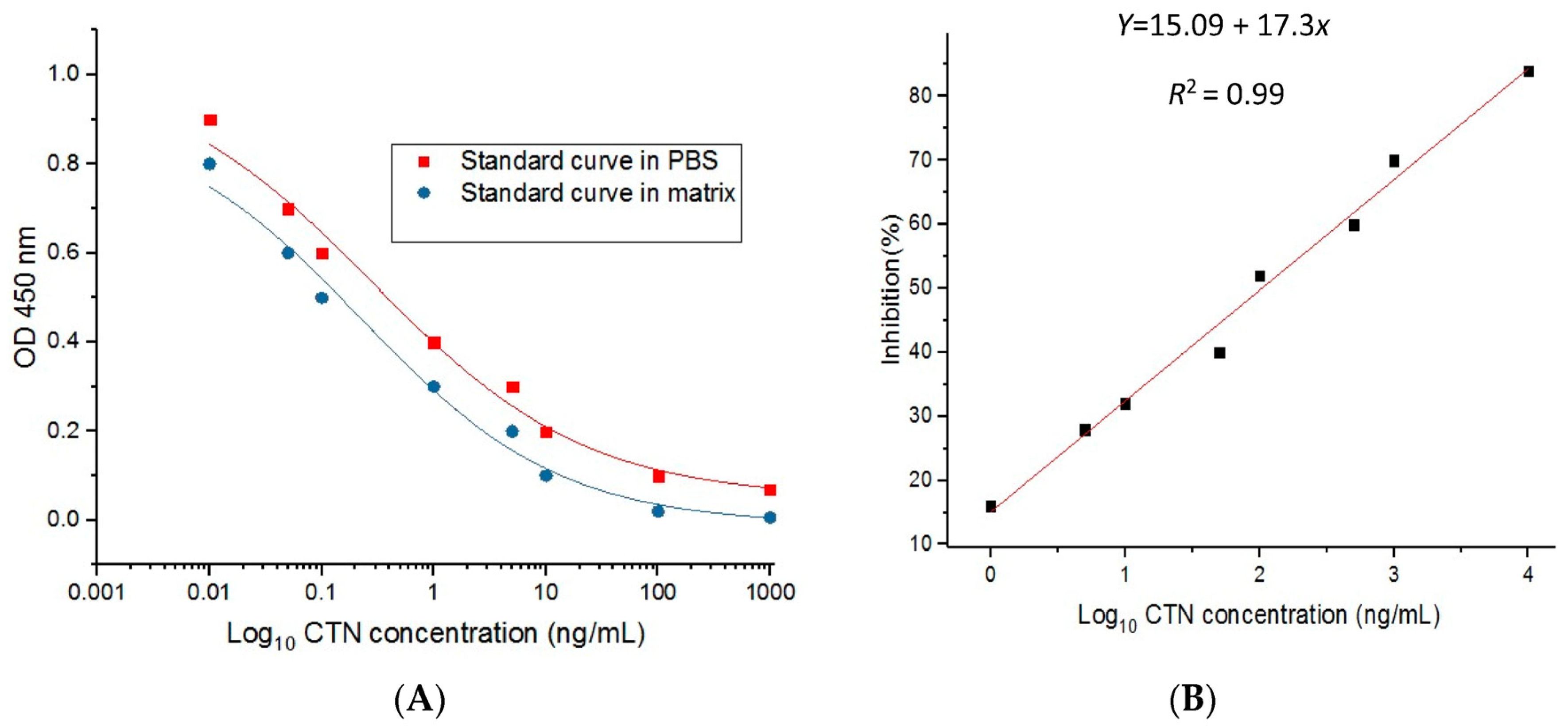
2.7. Standard Curve and Recovery Test
2.8. Samples Detection by ic-ELISA using Anti-CTN mcAb
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis and Characterization of CTN-Protein Conjugates
4.3. Analysis of CTN-Protein Conjugates
4.4. Mice Immunization and Anti-Serum ELISA Assay
4.5. Cell Fusion and Screening of Anti-CTN mcAb
4.6. Isotyping and Chromosome Analysis of the Anti-CTN mcAb
4.7. Production and Purification of Anti-CTN mcAb
4.8. Affinity Assay and Cross-Reactivity of Anti-CTN mcAb
4.9. Standard Curve and Recovery Test
4.10. Ethical Statement and Animal Care
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Da Rocha, M.E.B.; Freire, F.D.C.O.; Erlan Feitosa Maia, F.; Izabel Florindo Guedes, M.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Hetherington, A.C.; Raistrick, H. Studies in the biochemistry of micro-organisms. XIV. On the production and chemical constitution of a new yellow coloring matter, citrinin, produced from glucose by Penicillium citrinum, Thom. Philos. Trans. R. Soc. Ser. B 1931, 220, 269. [Google Scholar] [CrossRef]
- Blanc, P.J.; Laussac, J.P.; Le Bars, J.; Le Bars, P.; Loret, M.O.; Pareilleux, A.; Prome, D.; Prome, J.C.; Santerre, A.L.; Goma, G. Characterization of monascidin a from Monascus as citrinin. Int. J. Food Microbiol. 1995, 27, 201–213. [Google Scholar] [CrossRef]
- Reiss, J. Mycotoxins in foodstuffs. X. Production of citrinin by Penicillium chrysogenum in bread. Food Cosmet. Toxicol. 1977, 15, 303–307. [Google Scholar] [CrossRef]
- Nejati, P.; Chaychi Nosrati, A.; Bayat, M. An investigation on measurement means of citrinin toxin quantity by toxigenic Aspergillus species in biomass, using ELISA. Int. J. Adv. Biol. Biomed. Res. 2014, 2, 2466–2471. [Google Scholar]
- Petkova-Bocharova, T.; Castegnaro, M.; Michelon, J.; Maru, V. Ochratoxin A and Other Mycotoxins in Cereals from an Area of Balkan Endemic Nephropathy and Urinary Tract Tumours in Bulgaria; IARC Scientific Publications: Lyon, France, 1990; pp. 83–87. [Google Scholar]
- Ostry, V.; Malir, F.; Ruprich, J. Producers and important dietary sources of ochratoxin A and citrinin. Toxins 2013, 5, 1574–1586. [Google Scholar] [CrossRef] [PubMed]
- Ciegler, A.; Vesonder, R.; Jackson, L.K. Produciton and biological activity of patulin and citrinin from penicillium expansum. Appl. Environ. Microbiol. 1977, 33, 1004–1006. [Google Scholar] [PubMed]
- Zaied, C.; Zouaoui, N.; Bacha, H.; Abid, S. Natural occurrence of citrinin in tunisian wheat grains. Food Control 2012, 28, 106–109. [Google Scholar] [CrossRef]
- Tölgyesi, Á.; Stroka, J.; Tamosiunas, V.; Zwickel, T. Simultaneous analysis of Alternaria toxins and citrinin in tomato: An optimised method using liquid chromatography-tandem mass spectrometry. Food Addit. Contam. Part A 2015, 32, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Bailly, J.; Querin, A.; Le Bars-Bailly, S.; Benard, G.; Guerre, P. Citrinin production and stability in cheese. J. Food Prot. 2002, 65, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.H.; Lin, T.F. Monascus rice products. In Advances in Food and Nutrition Research; Academic Press: University of Nebraska, Lincoln, NE, USA, 2007; Volume 53, pp. 123–159. [Google Scholar]
- Li, Y.; Zhou, Y.-C.; Yang, M.-H.; Ou-Yang, Z. Natural occurrence of citrinin in widely consumed traditional chinese food red yeast rice, medicinal plants and their related products. Food Chem. 2012, 132, 1040–1045. [Google Scholar] [CrossRef]
- Li, F.; Xu, G.; Li, Y.; Chen, Y.; Ji, R. Natural occurrence of citrinin in Monascus products. Wei Sheng Yan Jiu = J. Hyg. Res. 2005, 34, 451–454. [Google Scholar]
- Sabater-Vilar, M.; Maas, R.F.; Fink-Gremmels, J. Mutagenicity of commercial Monascus fermentation products and the role of citrinin contamination. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 1999, 444, 7–16. [Google Scholar] [CrossRef]
- Flajs, D.; Peraica, M. Toxicological properties of citrinin. Arch. Ind. Hyg. Toxicol. 2009, 60, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Frank, H.K. Citrinin. Zeitschrift für Ernährungswissenschaft 1992, 31, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Sasmal, D.; Bandyopadhyay, S.; Bagchi, G.; Chatterjee, T.; Dey, S. Hematological changes produced in mcie by ochratoxin A and citrinin. Toxicology 1983, 26, 55–62. [Google Scholar] [CrossRef]
- Liu, B.-H.; Wu, T.-S.; Su, M.-C.; Chung, C.P.; Yu, F.-Y. Evaluation of citrinin occurrence and cytotoxicity in Monascus fermentation products. J. Agric. Food Chem. 2005, 53, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Dönmez-Altuntas, H.; Dumlupinar, G.; Imamoglu, N.; Hamurcu, Z.; Liman, B.C. Effects of the mycotoxin citrinin on micronucleus formation in a cytokinesis-block genotoxicity assay in cultured human lymphocytes. J. Appl. Toxicol. 2007, 27, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Qingqing, H.; Linbo, Y.; Yunqian, G.; Shuqiang, L. Toxic effects of citrinin on the male reproductive system in mice. Exp. Toxicol. Pathol. 2012, 64, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sharma, A.; Patil, R.; Rahman, S.; Leishangthem, G.; Kumar, M. Effect of feeding graded doses of citrinin on clinical and teratology in female Wistar rats. Indian J. Exp. Biol. 2014, 52, 159–167. [Google Scholar] [PubMed]
- Böhm, J.; De Saeger, S.; Edler, L.; Fink-Gremmels, J.; Mantle, P.; Peraica, M.; Stetina, R.; Vrabcheva, T. Scientific opinion on the risks for public and animal health related to the presence of citrinin in food and feed. Eur. Food Saf. Auth. J. 2012, 10, 1–82. [Google Scholar]
- Santos, I.M.; Abrunhosa, L.; Venâncio, A.; Lima, N. The effect of culture preservation techniques on patulin and citrinin production by Penicillium expansum link. Lett. Appl. Microbiol. 2002, 35, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Vrabcheva, T.; Usleber, E.; Dietrich, R.; Märtlbauer, E. Co-occurrence of ochratoxin A and citrinin in cereals from Bulgarian villages with a history of Balkan endemic nephropathy. J. Agric. Food Chem. 2000, 48, 2483–2488. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Xu, J.; Wang, X.; Qi, P.; Wei, W.; Chen, X.; Li, R.; Zhou, Y. Citrinin determination in red fermented rice products by optimized extraction method coupled to liquid chromatography tandem mass spectrometry (LC-MS/MS). J. Food Sci. 2015, 80, T1438–T1444. [Google Scholar] [CrossRef] [PubMed]
- Huertas-Pérez, J.F.; Arroyo-Manzanares, N.; García-Campaña, A.M.; Gámiz-Gracia, L. High-throughput determination of citrinin in rice by ultra-high-performance liquid chromatography and fluorescence detection (UHPLC-FL). Food Addit. Contam. Part A 2015, 32, 1352–1357. [Google Scholar] [CrossRef] [PubMed]
- Shu, P.-Y.; Lin, C.-H. Simple and sensitive determination of citrinin in Monascus by GC-selected ion monitoring mass spectrometry. Anal. Sci. 2002, 18, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Abramson, D.; Usleber, E.; Märtlbauer, E. An indirect enzyme immunoassay for the mycotoxin citrinin. Appl. Environ. Microbiol. 1995, 61, 2007–2009. [Google Scholar] [PubMed]
- Xu, B.-J.; Jia, X.-Q.; Gu, L.-J.; Sung, C.-K. Review on the qualitative and quantitative analysis of the mycotoxin citrinin. Food Control 2006, 17, 271–285. [Google Scholar] [CrossRef]
- Cho, Y.-J.; Lee, D.-H.; Kim, D.-O.; Min, W.-K.; Bong, K.-T.; Lee, G.-G.; Seo, J.-H. Production of a monoclonal antibody against ochratoxin A and its application to immunochromatographic assay. J. Agric. Chem. 2005, 53, 8447–8451. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, Y.; Pan, F.; Liu, Z.; Wang, Z. Identification of tetrodotoxin antigens and a monoclonal antibody. Food Chem. 2009, 112, 582–586. [Google Scholar] [CrossRef]
- Gayathri, L.; Dhivya, R.; Dhanasekaran, D.; Periasamy, V.S.; Alshatwi, A.A.; Akbarsha, M.A. Hepatotoxic effect of ochratoxin A and citrinin, alone and in combination, and protective effect of vitamin E: In vitro study in Hep G2 cell. Food Chem. Toxicol. 2015, 83, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.T.; Mora, E.C. Toxicity of Penicillium citrinum AUA-532 contaminated corn and citrinin in broiler chicks. Poult. Sci. 1978, 57, 1221–1226. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; He, Z.; Yuan, H.; Wu, J.; Wen, L.; Li, R.; Zhang, M.; Yuan, L.; Yuan, Z. Generation and characterization of a monoclonal antibody to penicillic acid from Penicillium cyclopium. Afr. J. Biotechnol. 2010, 9, 3026–3031. [Google Scholar]
- Liu, R.; Yu, Z.; He, Q.; Xu, Y. Preparation and identification of a monoclonal antibody against citrinin. Wei Sheng Yan Jiu = J. Hyg. Res. 2007, 36, 190–193. [Google Scholar]
- Ling, S.; Pang, J.; Yu, J.; Wang, R.; Liu, L.; Ma, Y.; Zhang, Y.; Jin, N.; Wang, S. Preparation and identification of monoclonal antibody against fumonisin B 1 and development of detection by ic-ELISA. Toxicon 2014, 80, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Maragos, C.; Busman, M.; Plattner, R. Development of monoclonal antibodies for the fusarin mycotoxins. Food Addit. Contam. 2008, 25, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Guo, Y. Preparation of synthetic antigen and monoclonal antibody for indirect competitive ELISA of citrinin. Food Agric. Immunol. 2012, 23, 145–156. [Google Scholar] [CrossRef]
- Cheng, H.; Chen, Y.; Yang, Y.; Chen, X.; Guo, X.; Du, A. Characterization of anti-citrinin specific scFvs selected from non-immunized mouse splenocytes by eukaryotic ribosome display. PLoS ONE 2015, 10, e0131482. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Yang, W.J.; He, J.X.; Zhao, J.Z.; Justo, G.C.; Zhang, X.Y. Preparation of artificial antigen and egg yolk-derived immunoglobulin (IgY) of citrinin for enzyme-linked immunosorbent assay. Biomed. Environ. Sci. 2009, 22, 237–243. [Google Scholar]
- Jin, N.; Ling, S.; Yang, C.; Wang, S. Preparation and identification of monoclonal antibody against citreoviridin and development of detection by ic-ELISA. Toxicon 2014, 90, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Kozak, C.A.; Lawrence, J.B.; Ruddle, F.H. A sequential staining technique for the chromosomal analysis of interspecific mouse/hamster and mouse/human somatic cell hybrids. Exp. Cell Res. 1977, 105, 109–117. [Google Scholar] [CrossRef]
- Bai, L.; Qian, J.-H.; Wang, J. Purification of mouse IgG from ascites fluid and serum by caprylic acid and ammonium sulfate. J. Dali Med. Coll. 2000, 4, 001. [Google Scholar]
- Chang, Y.-C.; Gottlieb, D.I. Characterization of the proteins purified with monoclonal antibodies to glutamic acid decarboxylase. J. Neurosci. 1988, 8, 2123–2130. [Google Scholar] [PubMed]
- Beatty, J.D.; Beatty, B.G.; Vlahos, W.G. Measurement of monoclonal antibody affinity by non-competitive enzyme immunoassay. J. Immunol. Methods 1987, 100, 173–179. [Google Scholar] [CrossRef]
- Kido, K.; Edakuni, K.; Morinaga, O.; Tanaka, H.; Shoyama, Y. An enzyme-linked immunosorbent assay for aconitine-type alkaloids using an anti-aconitine monoclonal antibody. Anal. Chim. Acta 2008, 616, 109–114. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxins | Cross-Reactivity (%) |
|---|---|
| ZEA | <0.01% |
| PT | 0.01% |
| STG | <0.01% |
| T-2 | <0.01% |
| OTA | <0.01% |
| OVA | <0.01% |
| BSA | <0.01% |
| Spiked Level (ng/mL) | Measured Concentration (ng/mL) | Recovery (%) | CV (%) |
|---|---|---|---|
| 0.5 | 0.47 ± 0.01 | 94.5 ± 1.1 | 1.5 |
| 5 | 4.84 ± 0.06 | 96.7 ± 1.6 | 2.2 |
| 100 | 93.9 ± 3.0 | 93.9 ± 3.0 | 4.3 |
| 1000 | 902.0 ± 7.2 | 90.2 ± 0.7 | 1.0 |
| Average | 93.8 ± 1.6 | 2.3 |
| Samples | OD 450 nm | Detection Results |
|---|---|---|
| Control PBS (B0) | 1.07 ± 0.11 | ND |
| Wheat bread | 1.1 ± 0.07 | ND |
| Cheese | 1.03 ± 0.04 | ND |
| Apple | 1.00 ± 0.04 | ND |
| Tomato | 1.13 ± 0.09 | ND |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalayu Yirga, S.; Ling, S.; Yang, Y.; Yuan, J.; Wang, S. The Preparation and Identification of a Monoclonal Antibody against Citrinin and the Development of Detection via Indirect Competitive ELISA. Toxins 2017, 9, 110. https://doi.org/10.3390/toxins9030110
Kalayu Yirga S, Ling S, Yang Y, Yuan J, Wang S. The Preparation and Identification of a Monoclonal Antibody against Citrinin and the Development of Detection via Indirect Competitive ELISA. Toxins. 2017; 9(3):110. https://doi.org/10.3390/toxins9030110
Chicago/Turabian StyleKalayu Yirga, Shimuye, Sumei Ling, Yanling Yang, Jun Yuan, and Shihua Wang. 2017. "The Preparation and Identification of a Monoclonal Antibody against Citrinin and the Development of Detection via Indirect Competitive ELISA" Toxins 9, no. 3: 110. https://doi.org/10.3390/toxins9030110