The Long Non-Coding RNA RP5-1024C24.1 and Its Associated-Gene MPPED2 Are Down-Regulated in Human Thyroid Neoplasias and Act as Tumour Suppressors

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
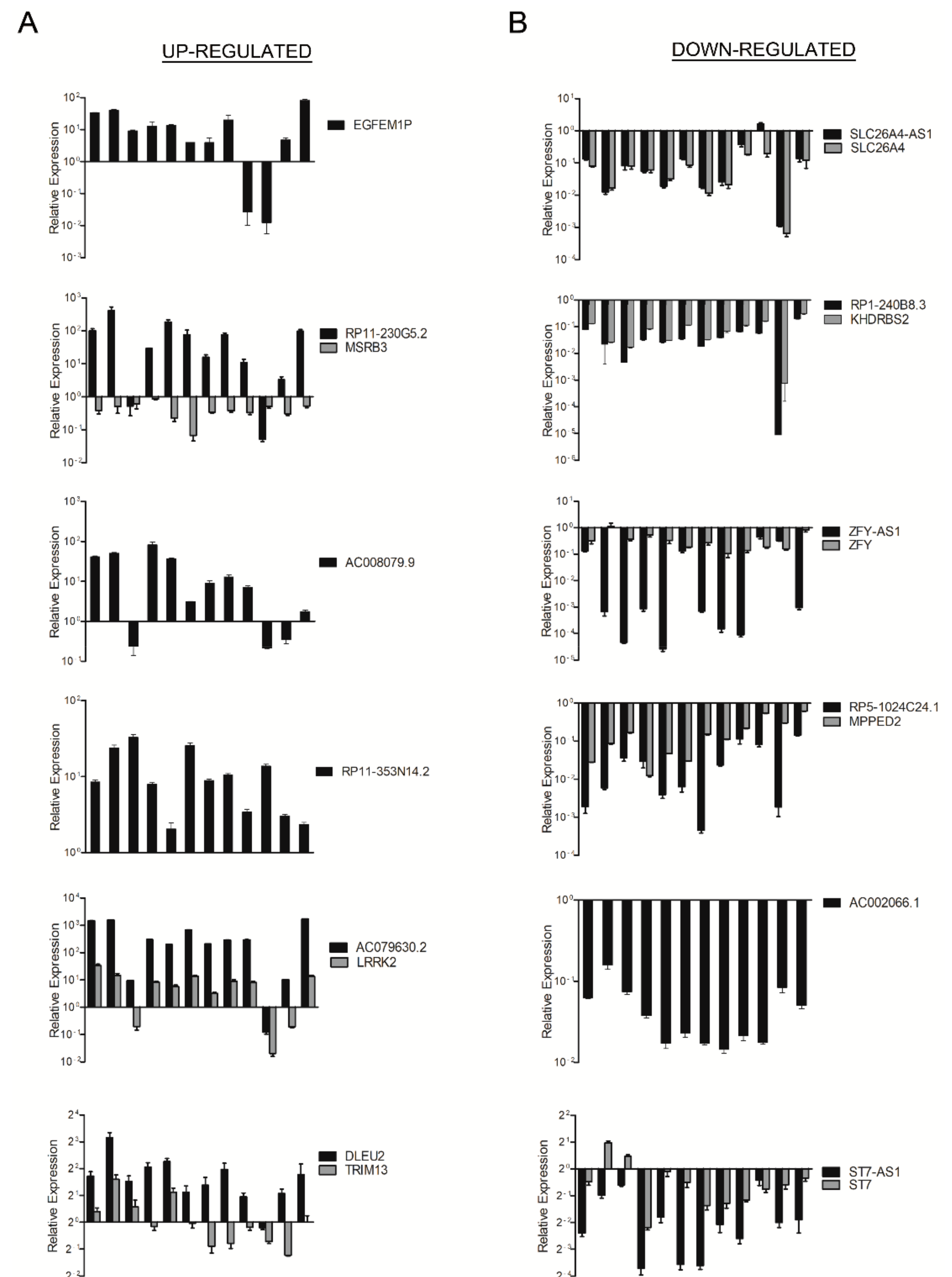
2.1. Identification of LncRNAs Deregulated in Human PTC
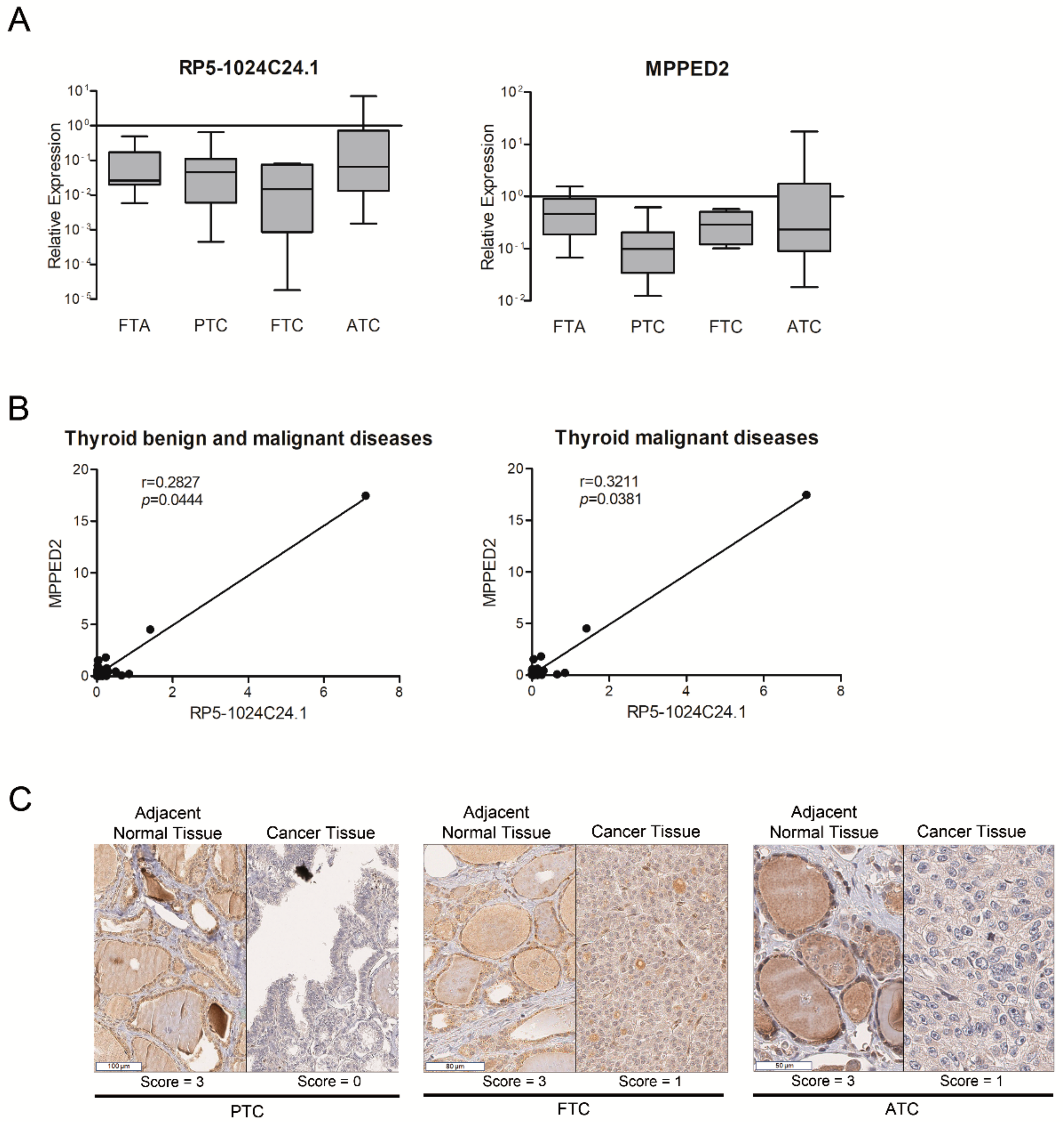
2.2. Analysis of RP5-1024C24.1 and MPPED2 Expression in Neoplastic Thyroid Diseases
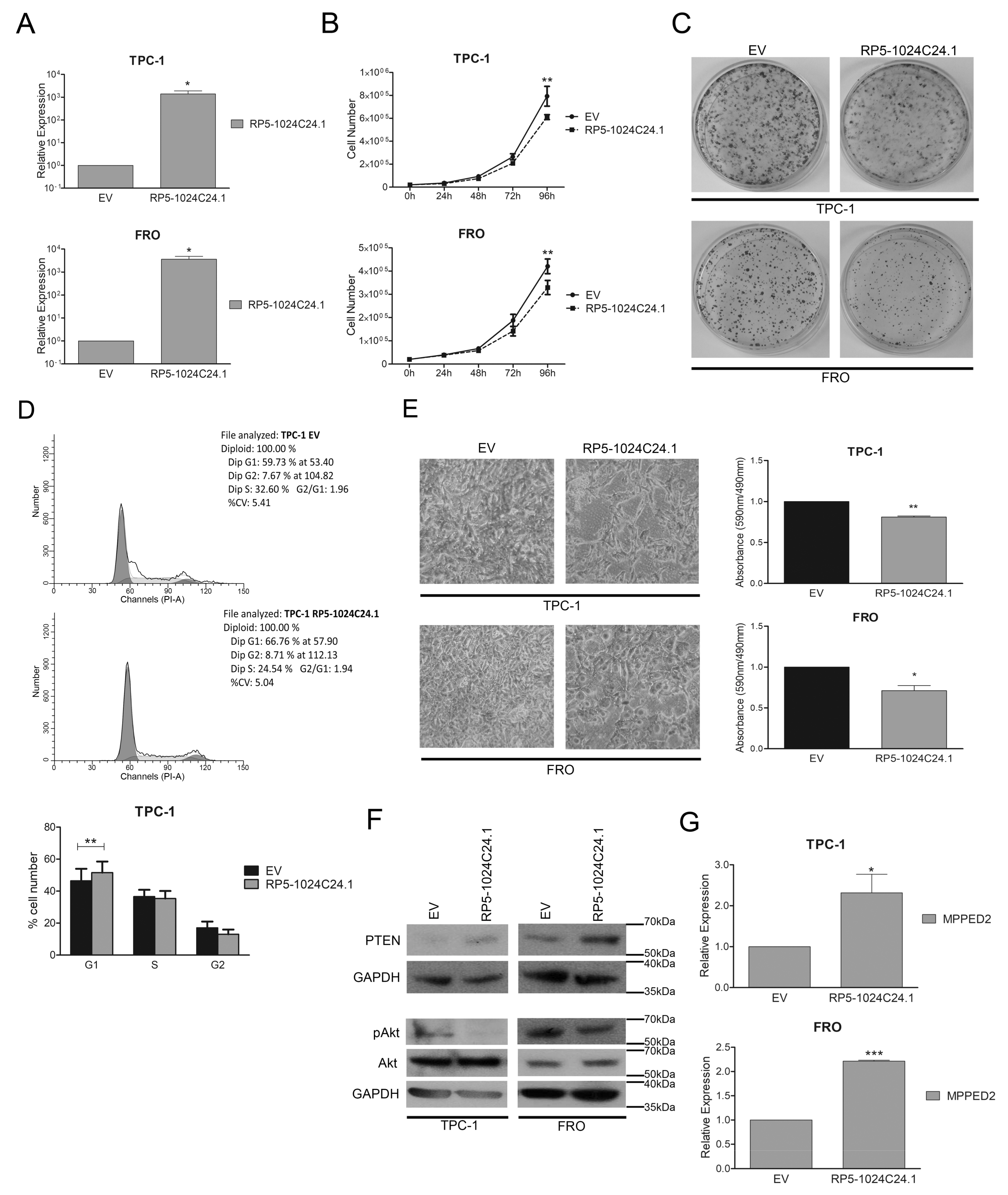
2.3. Down-Regulation of RP5-1024C24.1 Expression Contributes to Thyroid Carcinogenesis by Affecting the PTEN/Akt Pathway
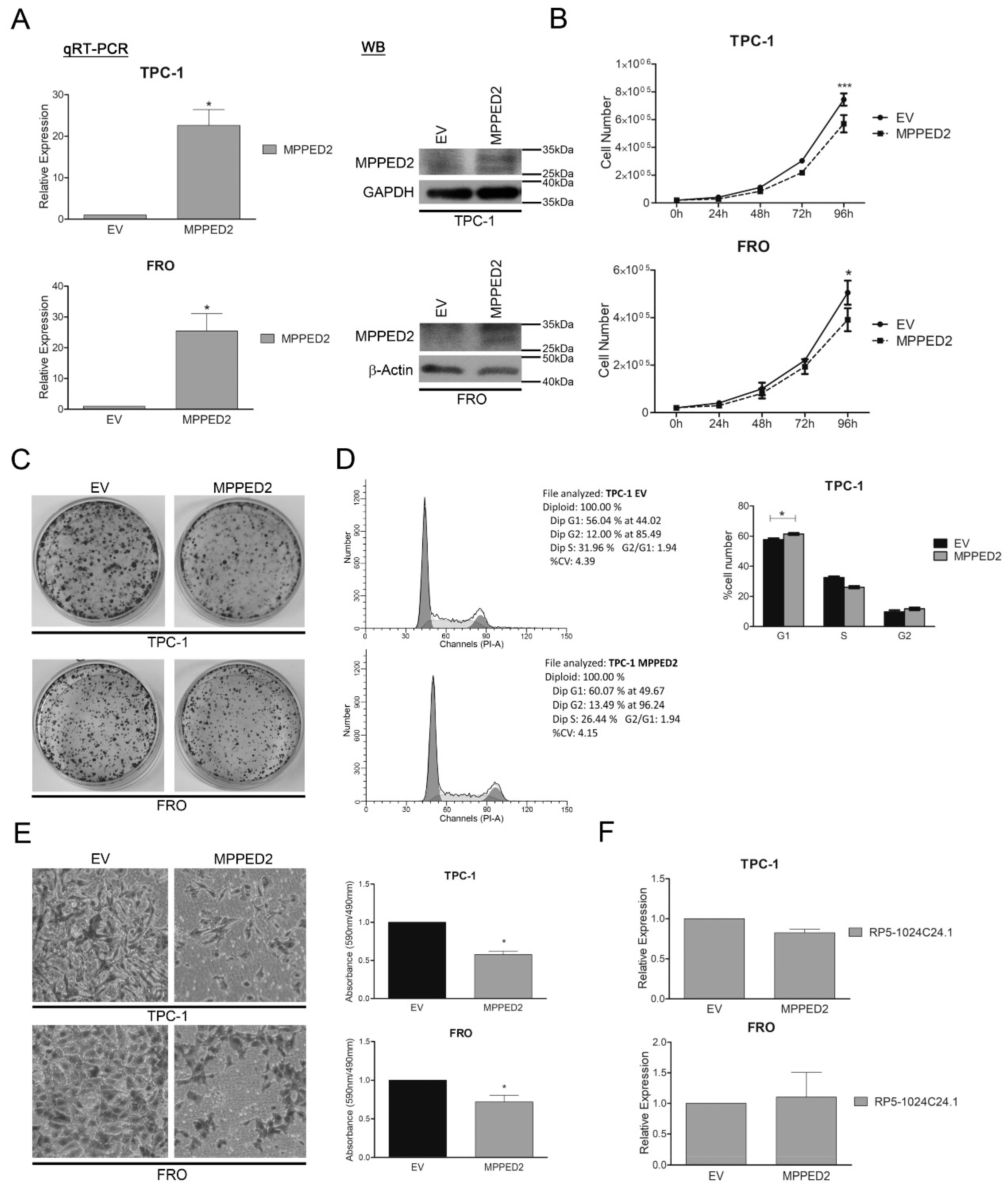
2.4. MPPED2 Is Induced by RP5-1024C24.1 and Negatively Modulates Cell Proliferation and Migration of Thyroid Carcinoma Cell Lines
3. Discussion
4. Materials and Methods
4.1. Human Thyroid Samples
4.2. Long Non-Coding RNA Microarray Analysis
4.3. Cell Lines and Transfection
4.4. Plasmids
4.5. RNA Extraction and qRT-PCR
4.6. Protein Extraction and Western Blot Analysis
4.7. Cell Migration Assays
4.8. Immunohistochemical Analysis
4.9. Flow Cytometry Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Availability of Data and Materials
References
- Parkin, D.M.; Bray, F.; Ferlay, J.; Pisani, P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005, 55, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Hedinger, C.; Williams, E.D.; Sobin, L.H. The who histological classification of thyroid tumors: A commentary on the second edition. Cancer 1989, 63, 908–911. [Google Scholar] [CrossRef]
- Fusco, A.; Grieco, M.; Santoro, M.; Berlingieri, M.T.; Pilotti, S.; Pierotti, M.A.; Della Porta, G.; Vecchio, G. A new oncogene in human thyroid papillary carcinomas and their lymph-nodal metastases. Nature 1987, 328, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Fusco, A.; Santoro, M. 20 years of RET/PTC in thyroid cancer: Clinico-pathological correlations. Arq. Bras. Endocrinol. Metabol. 2007, 51, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Grieco, M.; Santoro, M.; Berlingieri, M.T.; Melillo, R.M.; Donghi, R.; Bongarzone, I.; Pierotti, M.A.; Della Porta, G.; Fusco, A.; Vecchio, G. PTC is a novel rearranged form of the ret proto-oncogene and is frequently detected in vivo in human thyroid papillary carcinomas. Cell 1990, 60, 557–563. [Google Scholar] [CrossRef]
- Jung, C.K.; Little, M.P.; Lubin, J.H.; Brenner, A.V.; Wells, S.A., Jr.; Sigurdson, A.J.; Nikiforov, Y.E. The increase in thyroid cancer incidence during the last four decades is accompanied by a high frequency of BRAF mutations and a sharp increase in RAS mutations. J. Clin. Endocrinol. Metab. 2014, 99, E276–E285. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.; Liu, Z.; Zeng, W.; Maimaiti, Y.; Guo, Y.; Nie, X.; Chen, C.; Zhao, X.; Shi, L.; Liu, C.; et al. Association between BRAF and RAS mutations, and ret rearrangements and the clinical features of papillary thyroid cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 15155–15162. [Google Scholar] [PubMed]
- Penna, G.C.; Vaisman, F.; Vaisman, M.; Sobrinho-Simoes, M.; Soares, P. Molecular markers involved in tumorigenesis of thyroid carcinoma: Focus on aggressive histotypes. Cytogenet. Genome Res. 2017, 150, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Melillo, R.M.; Castellone, M.D.; Guarino, V.; De Falco, V.; Cirafici, A.M.; Salvatore, G.; Caiazzo, F.; Basolo, F.; Giannini, R.; Kruhoffer, M.; et al. The RET/PTC-RAS-BRAF linear signaling cascade mediates the motile and mitogenic phenotype of thyroid cancer cells. J. Clin. Investig. 2005, 115, 1068–1081. [Google Scholar] [CrossRef] [PubMed]
- Pallante, P.; Battista, S.; Pierantoni, G.M.; Fusco, A. Deregulation of microrna expression in thyroid neoplasias. Nat. Rev. Endocrinol. 2013, 10, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Diederichs, S. The hallmarks of cancer: A long non-coding RNA point of view. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Wapinski, O.; Chang, H.Y. Long noncoding RNAs and human disease. Trends Cell Biol. 2011, 21, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.G.; Yin, D.D.; Sun, S.Y.; Han, L. The use of lncRNA analysis for stratification management of prognostic risk in patients with NSCLC. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 115–119. [Google Scholar] [PubMed]
- Zhang, J.; Lin, Z.; Gao, Y.; Yao, T. Downregulation of long noncoding RNA MEG3 is associated with poor prognosis and promoter hypermethylation in cervical cancer. J. Exp. Clin. Cancer Res. 2017, 36, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, B.; Xiao, J.; Yang, Y.; Zhang, Y. A novel seven-long non-coding RNA signature predicts survival in early stage lung adenocarcinoma. Oncotarget 2017, 8, 14876–14886. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Huang, Y.; Chen, J.; Chi, H.; Yu, Z.; Wang, J.; Chen, C. Attenuated ability of bace1 to cleave the amyloid precursor protein via silencing long noncoding RNA bace1as expression. Mol. Med. Rep. 2014, 10, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Kotake, Y.; Nakagawa, T.; Kitagawa, K.; Suzuki, S.; Liu, N.; Kitagawa, M.; Xiong, Y. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15INK4B tumor suppressor gene. Oncogene 2011, 30, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Li, S.; Munoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular interplay of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by polycomb CBX7 in transcriptional silencing of INK4a. Mol. Cell. 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Beckedorff, F.C.; Ayupe, A.C.; Crocci-Souza, R.; Amaral, M.S.; Nakaya, H.I.; Soltys, D.T.; Menck, C.F.; Reis, E.M.; Verjovski-Almeida, S. The intronic long noncoding RNA ANRASSF1 recruits PRC2 to the RASSF1A promoter, reducing the expression of RASSF1A and increasing cell proliferation. PLoS Genet. 2013, 9, e1003705. [Google Scholar] [CrossRef] [PubMed]
- Berghoff, E.G.; Clark, M.F.; Chen, S.; Cajigas, I.; Leib, D.E.; Kohtz, J.D. Evf2 (Dlx6as) lncRNA regulates ultraconserved enhancer methylation and the differential transcriptional control of adjacent genes. Development 2013, 140, 4407–4416. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, S.T.; Boeva, V.; Escamilla-Del-Arenal, M.; Ancelin, K.; Granier, C.; Matias, N.R.; Sanulli, S.; Chow, J.; Schulz, E.; Picard, C.; et al. Jarid2 is implicated in the initial XIST-induced targeting of PRC2 to the inactive X chromosome. Mol. Cell. 2014, 53, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Liu, L.; Ge, L.; Xie, L.; Liu, S.; Sang, L.; Zhan, T.; Li, H. miR-448 downregulates MPPED2 to promote cancer proliferation and inhibit apoptosis in oral squamous cell carcinoma. Exp. Ther. Med. 2016, 12, 2747–2752. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Shen, C.; Zhao, L.; Wang, J.; McCrae, M.; Chen, X.; Lu, F. Dysregulation of host cellular genes targeted by human papillomavirus (HPV) integration contributes to HPV-related cervical carcinogenesis. Int. J. Cancer 2016, 138, 1163–1174. [Google Scholar] [CrossRef] [PubMed]
- Liguori, L.; Andolfo, I.; de Antonellis, P.; Aglio, V.; di Dato, V.; Marino, N.; Orlotti, N.I.; De Martino, D.; Capasso, M.; Petrosino, G.; et al. The metallophosphodiesterase Mpped2 impairs tumorigenesis in neuroblastoma. Cell Cycle 2012, 11, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Gu, J.; Matsumoto, K.; Aota, S.; Parsons, R.; Yamada, K.M. Inhibition of cell migration, spreading, and focal adhesions by tumor suppressor PTEN. Science 1998, 280, 1614–1617. [Google Scholar] [CrossRef] [PubMed]
- Song, M.S.; Salmena, L.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor. Nat. Rev. Mol. Cell Biol. 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lesche, R.; Li, D.M.; Liliental, J.; Zhang, H.; Gao, J.; Gavrilova, N.; Mueller, B.; Liu, X.; Wu, H. PTEN modulates cell cycle progression and cell survival by regulating phosphatidylinositol 3,4,5,-trisphosphate and AKT/protein kinase B signaling pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 6199–6204. [Google Scholar] [CrossRef] [PubMed]
- Stambolic, V.; Suzuki, A.; de la Pompa, J.L.; Brothers, G.M.; Mirtsos, C.; Sasaki, T.; Ruland, J.; Penninger, J.M.; Siderovski, D.P.; Mak, T.W. Negative regulation of PKB/AKT-dependent cell survival by the tumor suppressor PTEN. Cell 1998, 95, 29–39. [Google Scholar] [CrossRef]
- Xu, N.; Lao, Y.; Zhang, Y.; Gillespie, D.A. AKT: A double-edged sword in cell proliferation and genome stability. J. Oncol. 2012, 2012, 951724. [Google Scholar] [CrossRef] [PubMed]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sepe, R.; Formisano, U.; Federico, A.; Forzati, F.; Bastos, A.U.; D'Angelo, D.; Cacciola, N.A.; Fusco, A.; Pallante, P. CBX7 and hmga1b proteins act in opposite way on the regulation of the SPP1 gene expression. Oncotarget 2015, 6, 2680–2692. [Google Scholar] [CrossRef] [PubMed]
- Puca, F.; Colamaio, M.; Federico, A.; Gemei, M.; Tosti, N.; Bastos, A.U.; Del Vecchio, L.; Pece, S.; Battista, S.; Fusco, A. Hmga1 silencing restores normal stem cell characteristics in colon cancer stem cells by increasing p53 levels. Oncotarget 2014, 5, 3234–3245. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Up-Regulated LncRNAs (PTC vs. NT) | |||||||
| Gene Symbol | Seqname | Fold Change | p -Value | Chr. | Strand | Relationship | Associated Gene |
| XLOC_010052 | TCONS_00020760 | 44.796062 | 0.011679098 | chr12 | − | intergenic | |
| EGFEM1P | ENST00000488647 | 29.21525 | 0.003039263 | chr3 | + | intergenic | |
| RP11-230G5.2 | ENST00000538294 | 16.403324 | 0.003381364 | chr12 | − | natural antisense | MSRB3 |
| AC008079.9 | ENST00000434390 | 15.255612 | 0.017524553 | chr22 | − | intergenic | |
| RP11-353N14.2 | ENST00000576963 | 14.603789 | 6.1 × 10−5 | chr17 | + | intergenic | |
| AC079630.2 | ENST00000457989 | 12.705946 | 0.00535464 | chr12 | + | intergenic | LRRK2 |
| CTC-255N20.1 | ENST00000504297 | 6.02792 | 0.001935031 | chr5 | − | bidirectional | STK32A |
| AC003102.3 | ENST00000453562 | 5.626362 | 7.34 × 10−4 | chr17 | − | natural antisense | RUNDC3A |
| DLEU2 | uc001vdo.1 | 4.361354 | 9.4 × 10−4 | chr13 | − | natural antisense | TRIM13 |
| RP11-4C20.4 | ENST00000433110 | 3.8502407 | 0.035742607 | chr10 | + | intron sense-overlapping | PTPRE |
| Down-Regulated LncRNAs (PTC vs. NT) | |||||||
| Gene Symbol | Seqname | Fold Change | p -Value | Chr. | Strand | Relationship | Associated Gene |
| CTB-85P21.2 | ENST00000566630 | 72.72284 | 6.38 × 10−7 | chr5 | + | intergenic | |
| SLC26A4-AS1 | NR_028137 | 27.245485 | 0.006468582 | chr7 | − | natural antisense | SLC26A4 |
| RP11-317P15.5 | ENST00000570153 | 20.019192 | 0.007198248 | chr1 | − | intergenic | |
| RP1-240B8.3 | ENST00000511849 | 17.767982 | 0.001431982 | chr6 | − | exon sense-overlapping | KHDRBS2 |
| ZFY-AS1 | ENST00000417305 | 14.560697 | 0.017612867 | chrY | − | natural antisense | ZFY |
| LA16c-329F2.1 | ENST00000570022 | 12.498036 | 0.031231718 | chr16 | − | intronic antisense | MAPK8IP3 |
| RP5-1024C24.1 | ENST00000531002 | 10.285704 | 7.4 × 10−6 | chr11 | + | intronic antisense | MPPED2 |
| AC002066.1 | ENST00000439070 | 9.467349 | 8.22 × 10−4 | chr7 | − | intergenic | |
| ST7-AS1 | NR_002330 | 8.9516325 | 1.38 × 10−4 | chr7 | − | natural antisense | ST7 |
| RP11-121G22.3 | ENST00000553197 | 8.723473 | 5.89 × 10−6 | chr12 | + | intronic antisense | PPFIA2 |
| Histotype (n) | Reduced Expression 1 (n, %) | Whole Cohort |
|---|---|---|
| FTA 2 (n = 19) | n = 7 (37%) | p < 0.001 |
| PTC 3 (n = 147) | n = 116 (79%) | p < 0.001 |
| FTC 4 (n = 30) | n = 23 (77%) | p < 0.001 |
| PDC 5 (n = 37) | n = 32 (86%) | p < 0.001 |
| ATC 6 (n = 15) | n = 8 (53%) | p = 0.003 |
| Whole cohort p < 0.001 | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepe, R.; Pellecchia, S.; Serra, P.; D’Angelo, D.; Federico, A.; Raia, M.; Cortez Cardoso Penha, R.; Decaussin-Petrucci, M.; Del Vecchio, L.; Fusco, A.; et al. The Long Non-Coding RNA RP5-1024C24.1 and Its Associated-Gene MPPED2 Are Down-Regulated in Human Thyroid Neoplasias and Act as Tumour Suppressors. Cancers 2018, 10, 146. https://doi.org/10.3390/cancers10050146
Sepe R, Pellecchia S, Serra P, D’Angelo D, Federico A, Raia M, Cortez Cardoso Penha R, Decaussin-Petrucci M, Del Vecchio L, Fusco A, et al. The Long Non-Coding RNA RP5-1024C24.1 and Its Associated-Gene MPPED2 Are Down-Regulated in Human Thyroid Neoplasias and Act as Tumour Suppressors. Cancers. 2018; 10(5):146. https://doi.org/10.3390/cancers10050146
Chicago/Turabian StyleSepe, Romina, Simona Pellecchia, Pierre Serra, Daniela D’Angelo, Antonella Federico, Maddalena Raia, Ricardo Cortez Cardoso Penha, Myriam Decaussin-Petrucci, Luigi Del Vecchio, Alfredo Fusco, and et al. 2018. "The Long Non-Coding RNA RP5-1024C24.1 and Its Associated-Gene MPPED2 Are Down-Regulated in Human Thyroid Neoplasias and Act as Tumour Suppressors" Cancers 10, no. 5: 146. https://doi.org/10.3390/cancers10050146