Dysregulated HAI-2 Plays an Important Role in Renal Cell Carcinoma Bone Metastasis through Ligand-Dependent MET Phosphorylation

 and
and
Abstract
:1. Introduction
2. Results
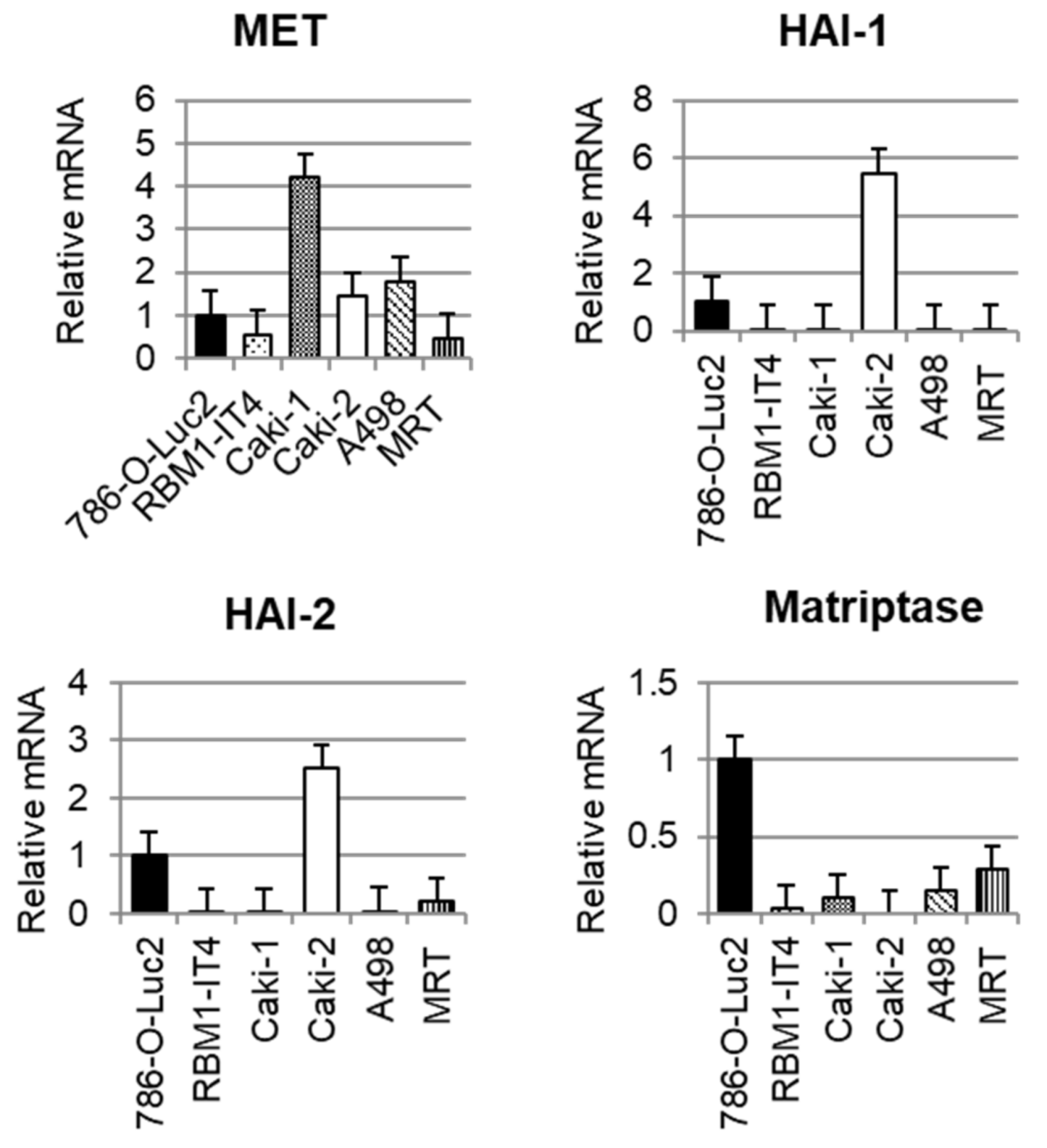
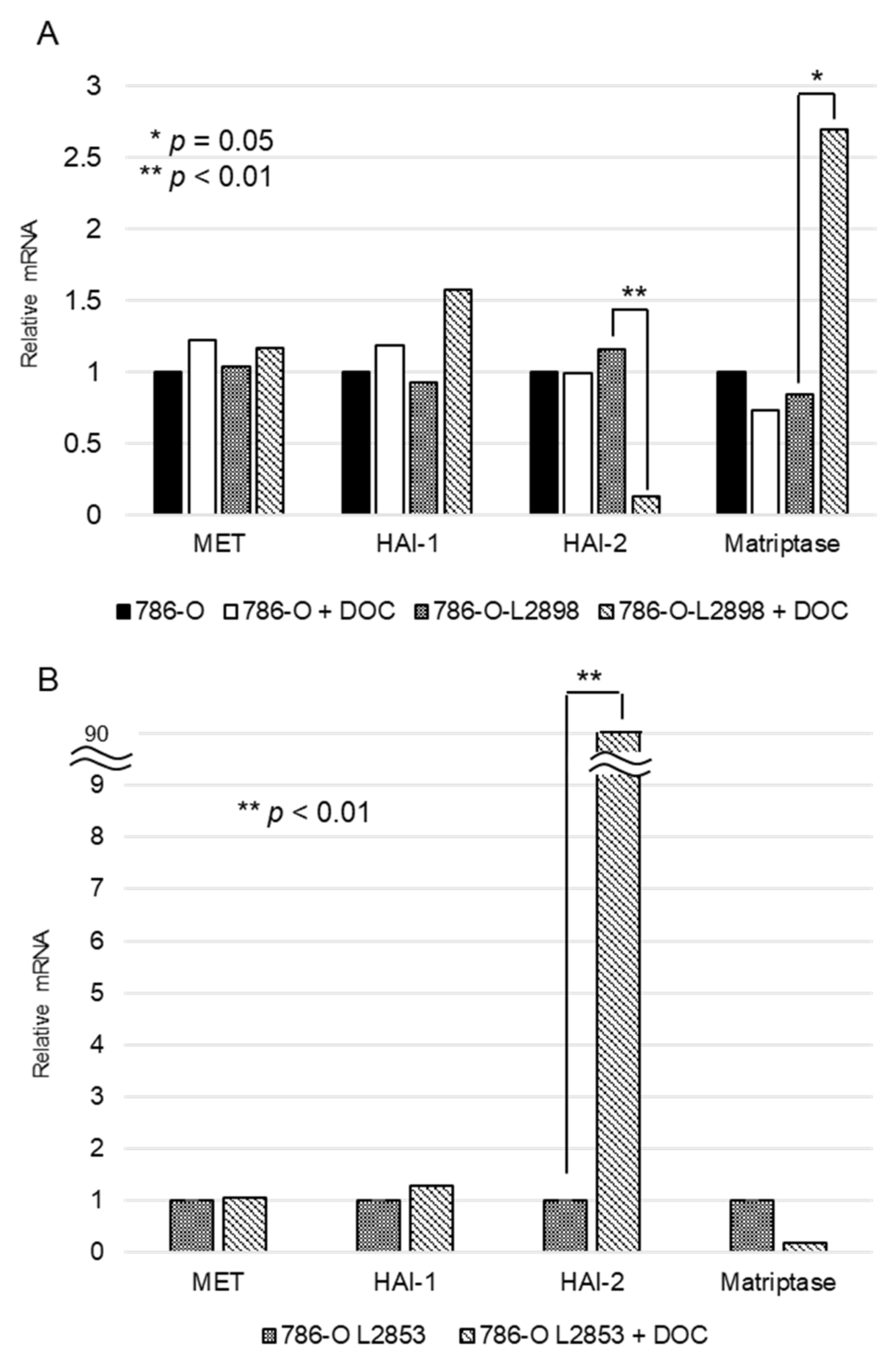
2.1. Expression of Each Molecule in RCC Cell Lines
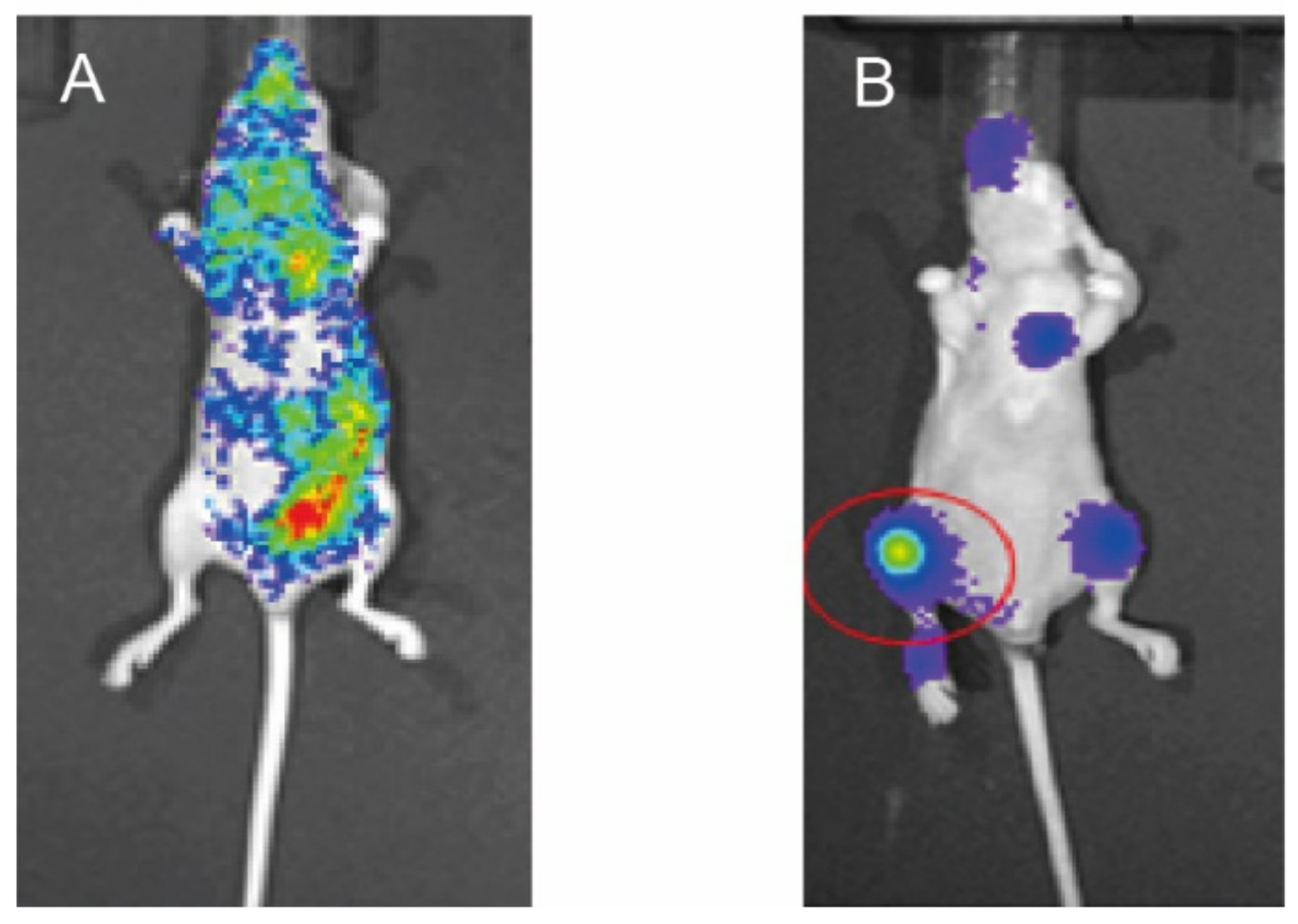
2.2. Preparation of Mouse Model of Bone Metastasis
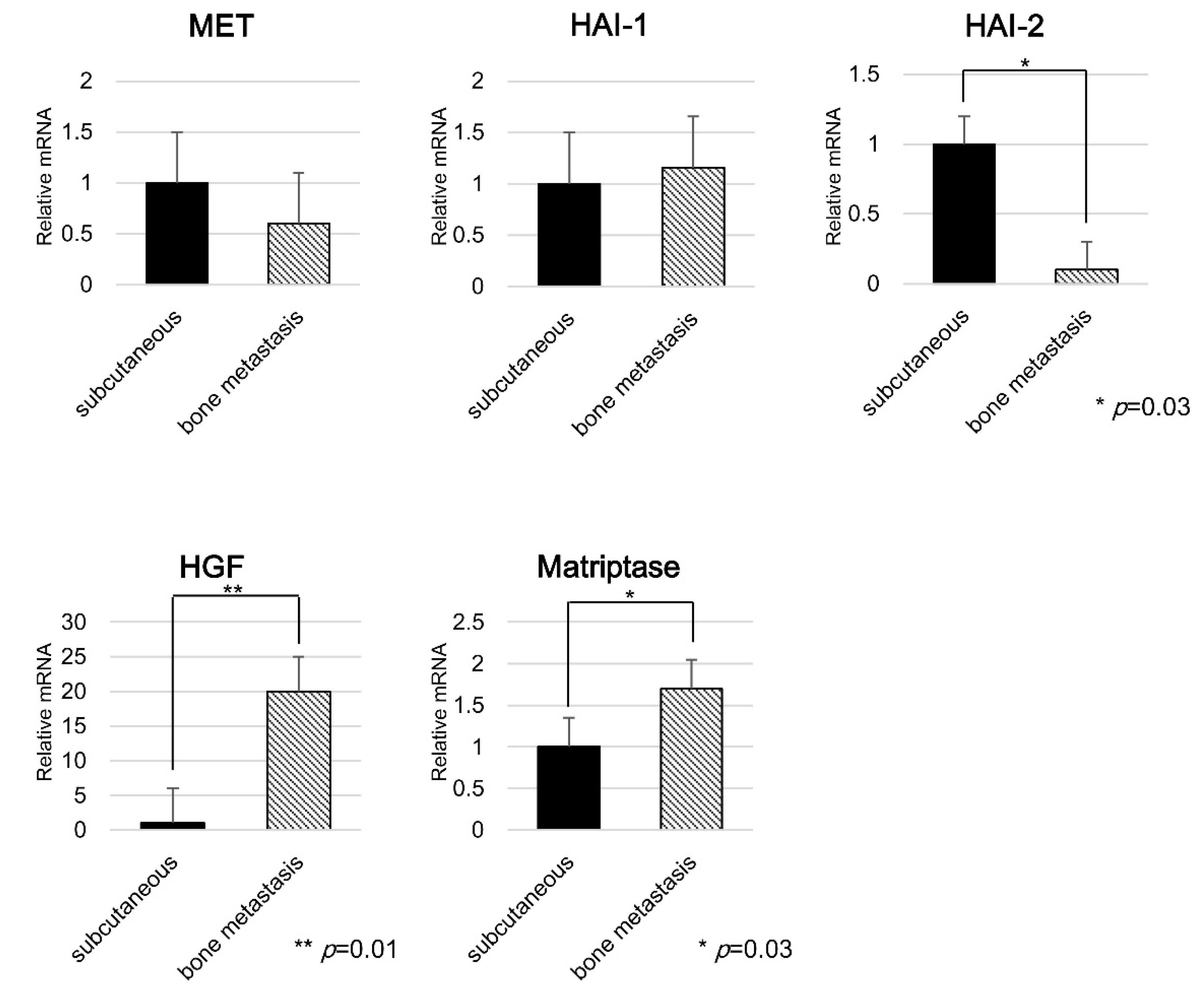
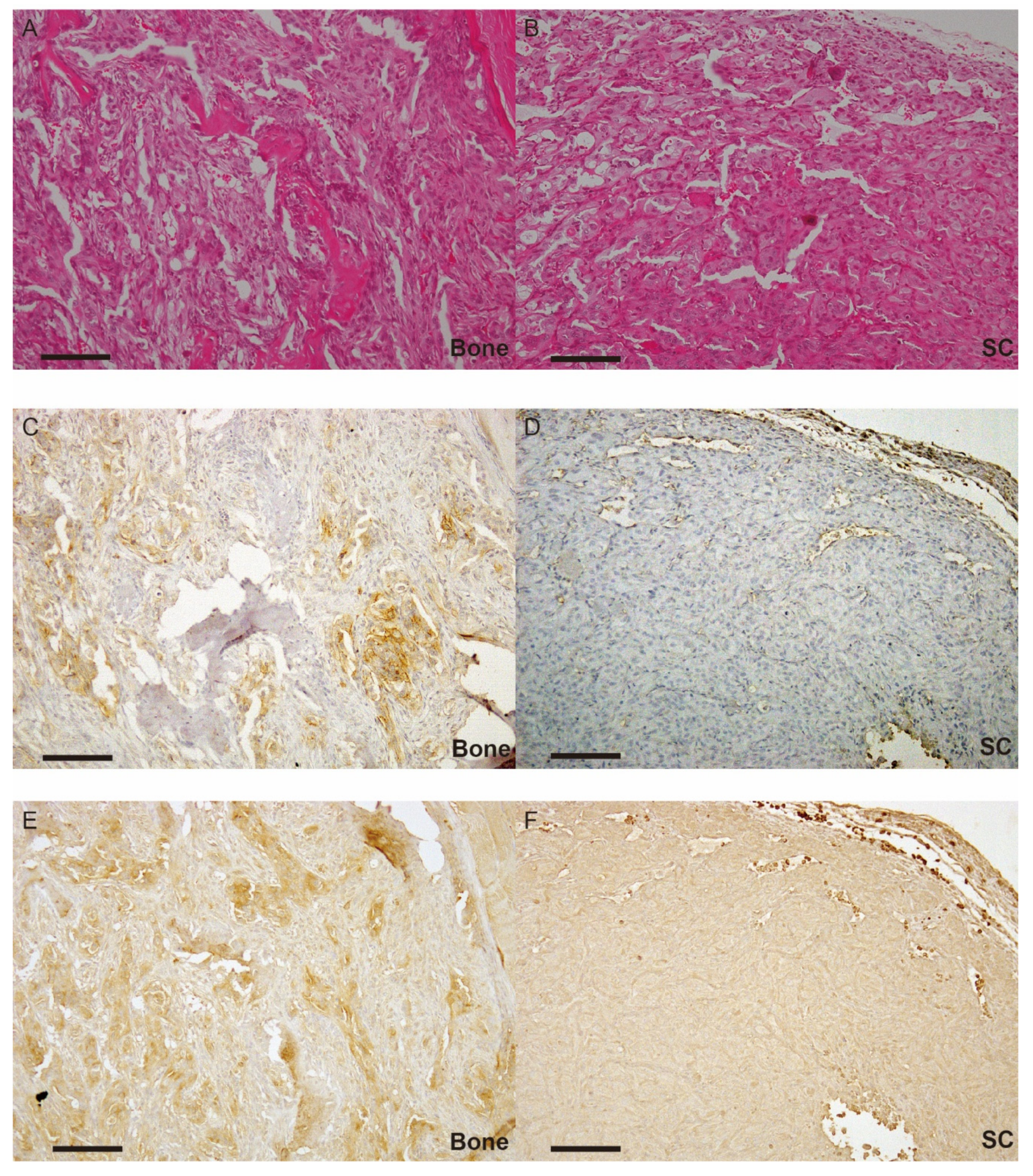
2.3. Expression of Each Molecule in Bone Metastasis
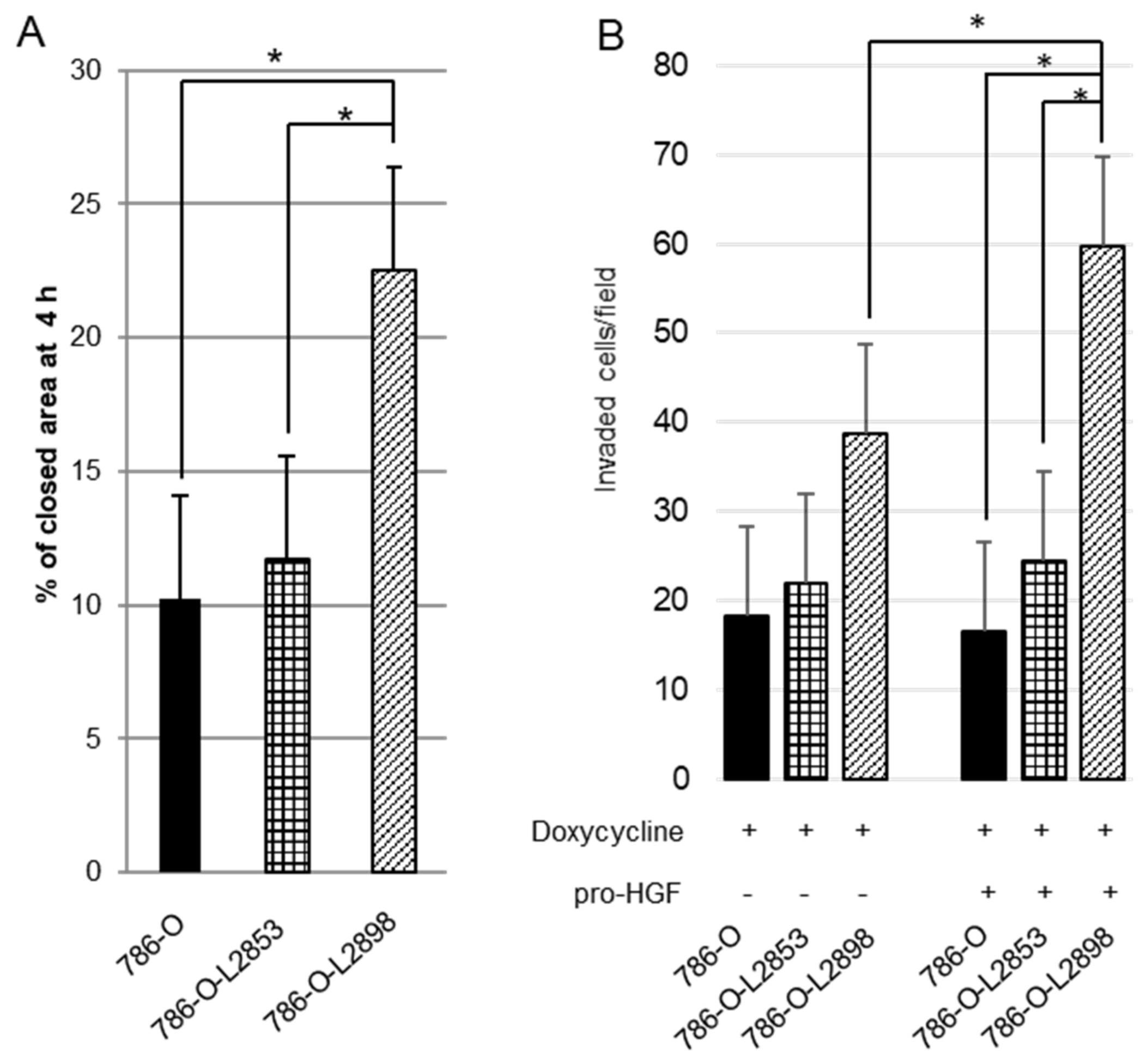
2.4. In Vitro Study Using HAI-2 Stable Knockdown and Overexpressed RCC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animal Experiments
4.3. RNA Extraction and RT-PCR
4.4. Real-Time Quantitative PCR
4.5. Immunohistochemistry
4.6. Construction of Lentiviral Vectors and Viral Transduction into 786-O-Luc2 Cells to Produce Doxycycline-Induced HAI-2 Stable Knockdown and Overexpression Cells
4.7. Wound Healing Assay
4.8. Invasion Assay
4.9. PCR Array
4.10. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, S.C.; Kuo, P.L. Bone metastasis from renal cell carcinoma. Int. J. Mol. Sci. 2016, 17, 987–1005. [Google Scholar] [CrossRef] [PubMed]
- Motzer, R.J.; Hutson, T.E.; Tomczak, P.; Michaelson, M.D.; Bukowski, R.M.; Rixe, O.; Oudard, S.; Negrier, S.; Szczylik, C.; Kim, S.T.; et al. Sunitinib versus interferon alfa in metastatic renal-cell carcinoma. N. Engl. J. Med. 2007, 356, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Rini, B.I.; Halabi, S.; Rosenberg, J.E.; Stadler, W.M.; Vaena, D.A.; Ou, S.S.; Archer, L.; Atkins, J.N.; Picus, J.; Czaykowski, P.; et al. Bevacizumab plus interferon alfa compared with interferon alfa monotherapy in patients with metastatic renal cell carcinoma: Calgb 90206. J. Clin. Oncol. 2008, 26, 5422–5428. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Figlin, R.A.; Hutson, T.E.; Michaelson, M.D.; Negrier, S.; Kim, S.T.; Huang, X.; Motzer, R.J. Prognostic factors for progression-free and overall survival with sunitinib targeted therapy and with cytokine as first-line therapy in patients with metastatic renal cell carcinoma. Ann. Oncol. 2011, 22, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.; Doucet, M.; Kominsky, S. Renal cell carcinoma bone metastasis--elucidating the molecular targets. Cancer Metastasis Rev. 2007, 26, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Rini, B.I.; Wilding, G.; Hudes, G.; Stadler, W.M.; Kim, S.; Tarazi, J.; Rosbrook, B.; Trask, P.C.; Wood, L.; Dutcher, J.P. Phase ii study of axitinib in sorafenib-refractory metastatic renal cell carcinoma. J. Clin. Oncol. 2009, 27, 4462–4468. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, T.K.; Escudier, B.; Powles, T.; Tannir, N.M.; Mainwaring, P.N.; Rini, B.I.; Hammers, H.J.; Donskov, F.; Roth, B.J.; Peltola, K.; et al. Cabozantinib versus everolimus in advanced renal cell carcinoma (meteor): Final results from a randomised, open-label, phase 3 trial. Lancet Oncol. 2016, 17, 917–927. [Google Scholar] [CrossRef]
- Escudier, B.; Powles, T.; Motzer, R.J.; Olencki, T.; Aren Frontera, O.; Oudard, S.; Rolland, F.; Tomczak, P.; Castellano, D.; Appleman, L.J.; et al. Cabozantinib, a new standard of care for patients with advanced renal cell carcinoma and bone metastases? Subgroup analysis of the meteor trial. J. Clin. Oncol. 2018, 36, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Giubellino, A.; Linehan, W.M.; Bottaro, D.P. Targeting the met signaling pathway in renal cancer. Expert Rev. Anticancer Ther. 2009, 9, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Sweeney, C.J.; Corn, P.G.; Rathkopf, D.E.; Smith, D.C.; Hussain, M.; George, D.J.; Higano, C.S.; Harzstark, A.L.; Sartor, A.O.; et al. Cabozantinib in chemotherapy-pretreated metastatic castration-resistant prostate cancer: Results of a phase ii nonrandomized expansion study. J. Clin. Oncol. 2014, 32, 3391–3399. [Google Scholar] [CrossRef] [PubMed]
- Betsunoh, H.; Mukai, S.; Akiyama, Y.; Fukushima, T.; Minamiguchi, N.; Hasui, Y.; Osada, Y.; Kataoka, H. Clinical relevance of hepsin and hepatocyte growth factor activator inhibitor type 2 expression in renal cell carcinoma. Cancer Sci. 2007, 98, 491–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, H.; Kawaguchi, M.; Fukushima, T.; Shimomura, T. Hepatocyte growth factor activator inhibitors (HAI-1 and HAI-2): Emerging key players in epithelial integrity and cancer. Pathol. Int. 2018, 68, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Kanetake, H.; Kanda, S. Presence of phosphorylated hepatocyte growth factor receptor/c-met is associated with tumor progression and survival in patients with conventional renal cell carcinoma. Clin. Cancer Res. 2006, 12, 4876–4881. [Google Scholar] [CrossRef] [PubMed]
- Mukai, S.; Yorita, K.; Kawagoe, Y.; Katayama, Y.; Nakahara, K.; Kamibeppu, T.; Sugie, S.; Tukino, H.; Kamoto, T.; Kataoka, H. Matriptase and met are prominently expressed at the site of bone metastasis in renal cell carcinoma: Immunohistochemical analysis. Hum. Cell 2015, 28, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Mukai, S.; Yorita, K.; Yamasaki, K.; Nagai, T.; Kamibeppu, T.; Sugie, S.; Kida, K.; Onizuka, C.; Tsukino, H.; Kamimura, T.; et al. Expression of human kallikrein 1-related peptidase 4 (KLK4) and met phosphorylation in prostate cancer tissue: Immunohistochemical analysis. Hum. Cell 2015, 28, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Comoglio, P.M.; Giordano, S.; Trusolino, L. Drug development of met inhibitors: Targeting oncogene addiction and expedience. Nat. Rev. Drug Discov. 2008, 7, 504–516. [Google Scholar] [CrossRef] [PubMed]
- List, K.; Bugge, T.H.; Szabo, R. Matriptase: Potent proteolysis on the cell surface. Mol. Med. 2006, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T. Hepatocyte growth factor as mitogen, motogen and morphogen, and its roles in organ regeneration. Princess Takamatsu Symp. 1994, 24, 195–213. [Google Scholar] [PubMed]
- Bergum, C.; Zoratti, G.; Boerner, J.; List, K. Strong expression association between matriptase and its substrate prostasin in breast cancer. J. Cell. Physiol. 2012, 227, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.S.; Chen, A.; Hsieh, D.S.; Yao, C.W.; Cheng, M.F.; Lin, Y.F. Expression of serine protease matriptase in renal cell carcinoma: Correlation of tissue microarray immunohistochemical expression analysis results with clinicopathological parameters. Int. J. Surg. Pathol. 2006, 14, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, H.; Miyata, S.; Uchinokura, S.; Itoh, H. Roles of hepatocyte growth factor (HGF) activator and hgf activator inhibitor in the pericellular activation of HGF/scatter factor. Cancer Metastasis Rev. 2003, 22, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Najy, A.J.; Won, J.J.; Movilla, L.S.; Kim, H.R. Differential tumorigenic potential and matriptase activation between PDGF B versus PDGF D in prostate cancer. Mol. Cancer Res. 2012, 10, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.H.; Teng, C.H.; Tu, Y.T.; Cheng, T.S.; Wu, S.R.; Ko, C.J.; Shyu, H.Y.; Lan, S.W.; Huang, H.P.; Tzeng, S.F.; et al. HAI-2 suppresses the invasive growth and metastasis of prostate cancer through regulation of matriptase. Oncogene 2014, 33, 4643–4652. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, B.S.; Gmyrek, G.A.; Inra, J.; Scherr, D.S.; Vaughan, E.D.; Nanus, D.M.; Kattan, M.W.; Gerald, W.L.; Vande Woude, G.F. High expression of the met receptor in prostate cancer metastasis to bone. Urology 2002, 60, 1113–1117. [Google Scholar] [CrossRef]
- Previdi, S.; Maroni, P.; Matteucci, E.; Broggini, M.; Bendinelli, P.; Desiderio, M.A. Interaction between human-breast cancer metastasis and bone microenvironment through activated hepatocyte growth factor/met and beta-catenin/wnt pathways. Eur. J. Cancer 2010, 46, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- D'Amico, L.; Belisario, D.; Migliardi, G.; Grange, C.; Bussolati, B.; D’Amelio, P.; Perera, T.; Dalmasso, E.; Dalle Carbonare, L.; Godio, L.; et al. C-met inhibition blocks bone metastasis development induced by renal cancer stem cells. Oncotarget 2016, 7, 45525–45537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trusolino, L.; Bertotti, A.; Comoglio, P.M. Met signalling: Principles and functions in development, organ regeneration and cancer. Nat. Rev. Mol. Cell Biol. 2010, 11, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, S.; Comoglio, P.M. The met receptor tyrosine kinase in invasion and metastasis. J. Cell. Physiol. 2007, 213, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, T.; Denda, K.; Kitamura, A.; Kawaguchi, T.; Kito, M.; Kondo, J.; Kagaya, S.; Qin, L.; Takata, H.; Miyazawa, K.; et al. Hepatocyte growth factor activator inhibitor, a novel kunitz-type serine protease inhibitor. J. Biol. Chem. 1997, 272, 6370–6376. [Google Scholar] [CrossRef] [PubMed]
- Wyler, L.; Napoli, C.U.; Ingold, B.; Sulser, T.; Heikenwalder, M.; Schraml, P.; Moch, H. Brain metastasis in renal cancer patients: Metastatic pattern, tumour-associated macrophages and chemokine/chemoreceptor expression. Br. J. Cancer 2014, 110, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Behnam Azad, B.; Nimmagadda, S. The intricate role of CXCR4 in cancer. Adv. Cancer Res. 2014, 124, 31–82. [Google Scholar] [PubMed]
- Wu, C.J.; Feng, X.; Lu, M.; Morimura, S.; Udey, M.C. Matriptase-mediated cleavage of epcam destabilizes claudins and dysregulates intestinal epithelial homeostasis. J. Clin. Investig. 2017, 127, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Suda, K.; Mizuuchi, H.; Maehara, Y.; Mitsudomi, T. Acquired resistance mechanisms to tyrosine kinase inhibitors in lung cancer with activating epidermal growth factor receptor mutation—Diversity, ductility, and destiny. Cancer Metastasis Rev. 2012, 31, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Pennacchietti, S.; Cazzanti, M.; Bertotti, A.; Rideout, W.M., 3rd; Han, M.; Gyuris, J.; Perera, T.; Comoglio, P.M.; Trusolino, L.; Michieli, P. Microenvironment-derived HGF overcomes genetically determined sensitivity to anti-met drugs. Cancer Res. 2014, 74, 6598–6609. [Google Scholar] [CrossRef] [PubMed]
- Joffre, C.; Barrow, R.; Menard, L.; Calleja, V.; Hart, I.R.; Kermorgant, S. A direct role for met endocytosis in tumorigenesis. Nat. Cell Biol. 2011, 13, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.C.; Lyman, S.K.; McCauley, S.; Kovalenko, M.; Spangler, R.; Liu, C.; Lee, M.; O’Sullivan, C.; Barry-Hamilton, V.; Ghermazien, H.; et al. Selective allosteric inhibition of MMP9 is efficacious in preclinical models of ulcerative colitis and colorectal cancer. PLoS ONE 2015, 10, e0127063. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, K.; Kataoka, H.; Nabeshima, K.; Seguchi, K.; Moriyama, T.; Osada, Y.; Koono, M. Establishment and characterization of a human renal carcinoma cell line MRT-1, with special reference to the production of serine proteinase inhibitors. Hum. Cell 1995, 8, 11–18. [Google Scholar] [PubMed]
- Schefe, J.H.; Lehmann, K.E.; Buschhmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT-PCR data analysis: Current concepts and novel “gene expression’s CT difference” formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | 786-O-L2898 Fold Change | 786-O-L2853 Fold Change |
|---|---|---|---|
| CCL7 | Chemokine (C-C motif) ligand 7 | 4.29 | 1.33 |
| CDH1 | Cadherin 1, type 1, E-cadherin | −1.08 | 1.72 |
| CXCR4 | Chemokine (C-X-C motif) receptor 4 | 2.57 | −2.45 |
| MMP9 | Matrix metalloprotease 9 | 1.69 | −7.29 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamasaki, K.; Mukai, S.; Sugie, S.; Nagai, T.; Nakahara, K.; Kamibeppu, T.; Sakamoto, H.; Shibasaki, N.; Terada, N.; Toda, Y.; et al. Dysregulated HAI-2 Plays an Important Role in Renal Cell Carcinoma Bone Metastasis through Ligand-Dependent MET Phosphorylation. Cancers 2018, 10, 190. https://doi.org/10.3390/cancers10060190
Yamasaki K, Mukai S, Sugie S, Nagai T, Nakahara K, Kamibeppu T, Sakamoto H, Shibasaki N, Terada N, Toda Y, et al. Dysregulated HAI-2 Plays an Important Role in Renal Cell Carcinoma Bone Metastasis through Ligand-Dependent MET Phosphorylation. Cancers. 2018; 10(6):190. https://doi.org/10.3390/cancers10060190
Chicago/Turabian StyleYamasaki, Koji, Shoichiro Mukai, Satoru Sugie, Takahiro Nagai, Kozue Nakahara, Toyoharu Kamibeppu, Hiromasa Sakamoto, Noboru Shibasaki, Naoki Terada, Yoshinobu Toda, and et al. 2018. "Dysregulated HAI-2 Plays an Important Role in Renal Cell Carcinoma Bone Metastasis through Ligand-Dependent MET Phosphorylation" Cancers 10, no. 6: 190. https://doi.org/10.3390/cancers10060190
APA StyleYamasaki, K., Mukai, S., Sugie, S., Nagai, T., Nakahara, K., Kamibeppu, T., Sakamoto, H., Shibasaki, N., Terada, N., Toda, Y., Kataoka, H., & Kamoto, T. (2018). Dysregulated HAI-2 Plays an Important Role in Renal Cell Carcinoma Bone Metastasis through Ligand-Dependent MET Phosphorylation. Cancers, 10(6), 190. https://doi.org/10.3390/cancers10060190