F-Box/WD Repeat Domain-Containing 7 Induces Chemotherapy Resistance in Colorectal Cancer Stem Cells

 , and
, and
Abstract
:1. Introduction
2. Results
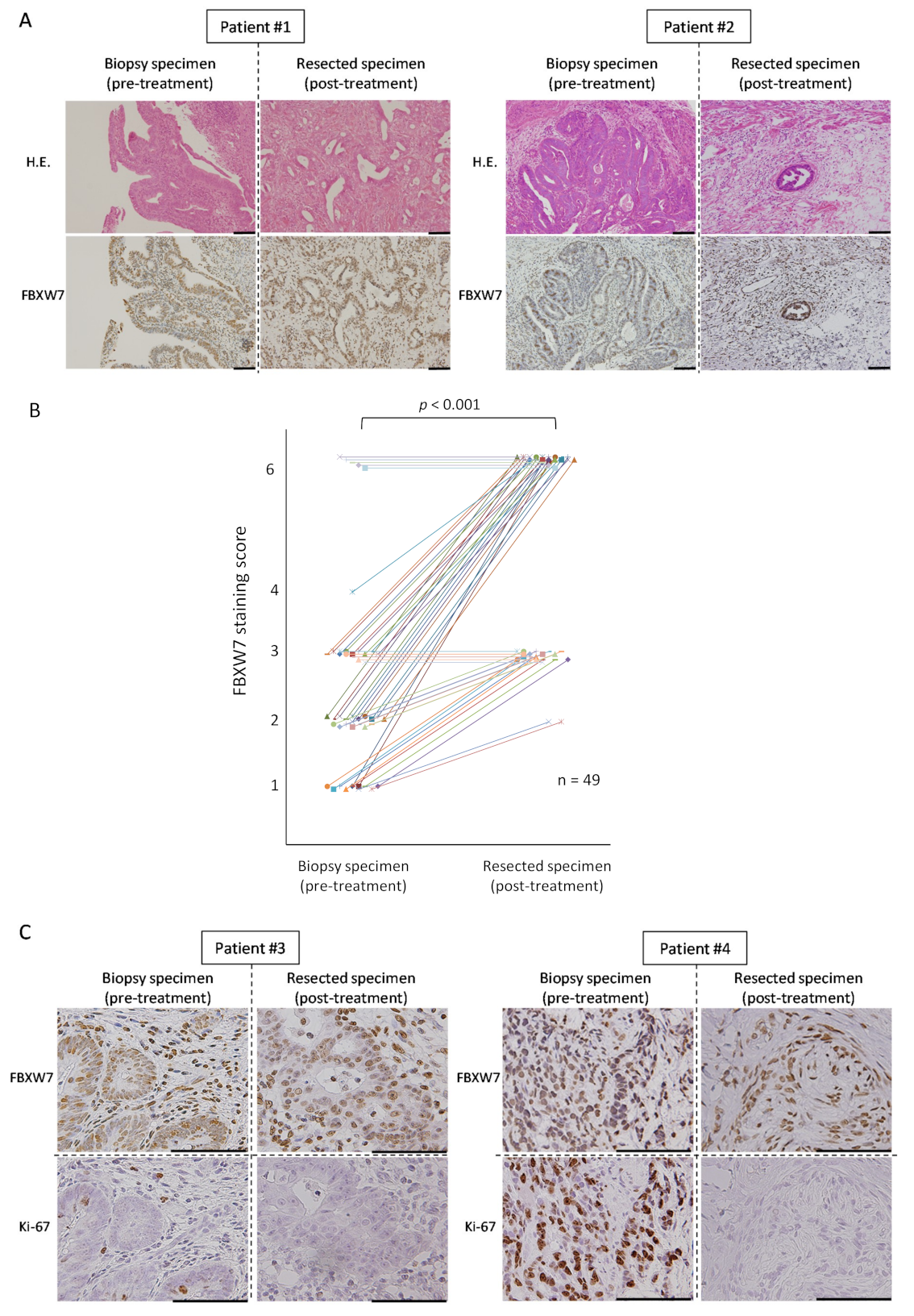
2.1. High FBXW7 Expression in Pre-Treatment Biopsy Specimens Is Related to Poor Pathological Theraperutic Effect
2.2. FBXW7 Is Highly Expressed in the Post-Treatement Resected Specimens
2.3. FBXW7 Expression in the EpCAMhigh/CD44high Population Was Significantly Higher Than That in The EpCAMhigh/CD44low Population
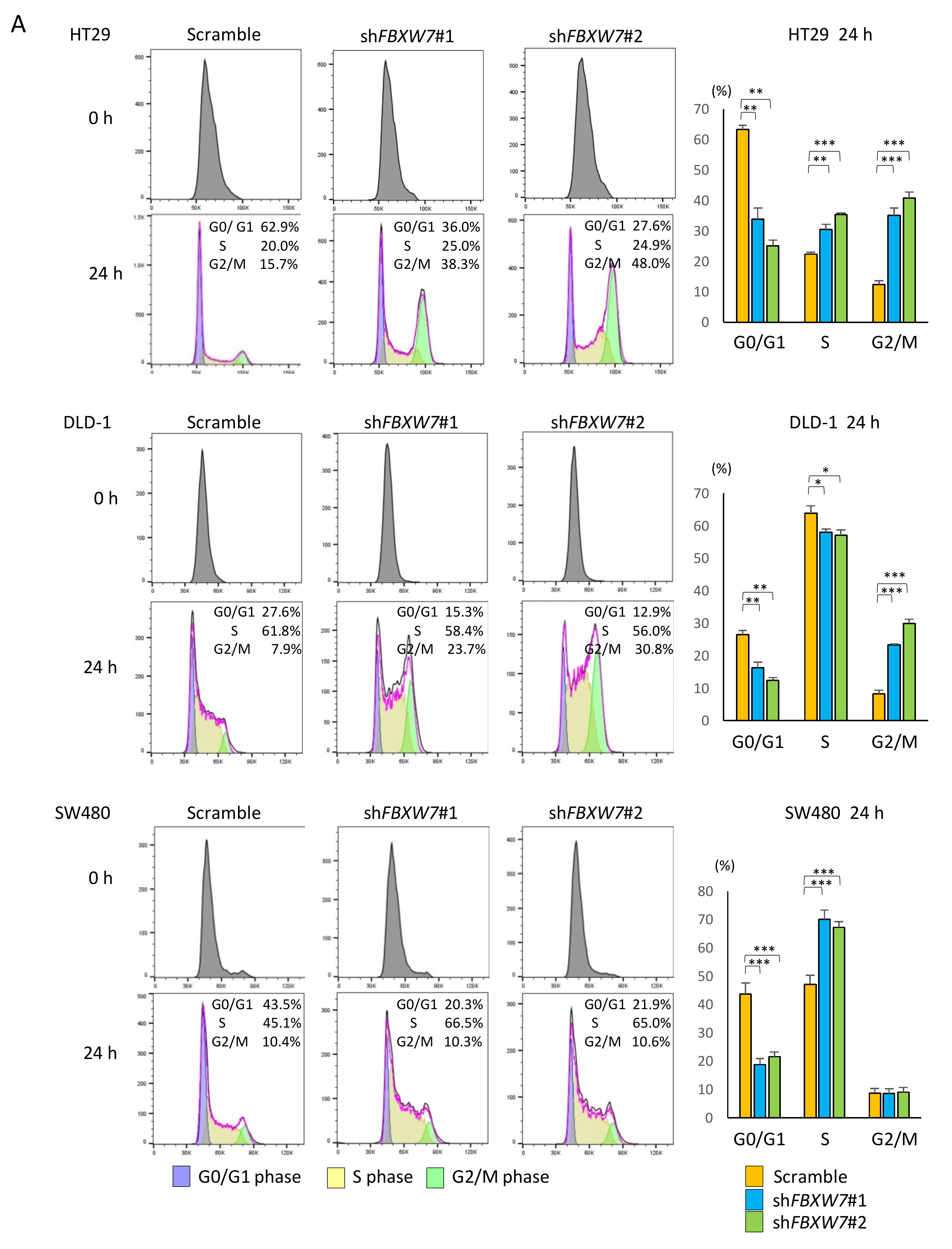
2.4. FBXW7 Knockdown Accelerates Cell cycle and Cell Proliferation, Resulting in Increased Sensitivity to Anti-Cancer Drugs in CRC Cells
2.5. FBXW7 Knockdown Inhibits Tumor Growth Rates upon Chemotherapy Treatment in Vivo
3. Discussion
4. Materials and Methods
4.1. Patient Population
4.2. Cell Lines and Reagents
4.3. IHC Studies
4.4. Xenograft Analysis
4.5. RNA Isolation and qPCR
4.6. Cell Synchronization and Cell Cycle Analysis
4.7. Luminescence-Based Proliferation and Cytotoxicity Assays
4.8. FBXW7 Protein Analysis
4.9. Western Blotting Analysis
4.10. In vivo Xenograft Studies
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- Bosset, J.F.; Collette, L.; Calais, G.; Mineur, L.; Maingon, P.; Radosevic-Jelic, L.; Daban, A.; Bardet, E.; Beny, A.; Ollier, J.C. Chemotherapy with preoperative radiotherapy in rectal cancer. N. Engl. J. Med. 2006, 355, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Gerard, J.P.; Conroy, T.; Bonnetain, F.; Bouche, O.; Chapet, O.; Closon-Dejardin, M.T.; Untereiner, M.; Leduc, B.; Francois, E.; Maurel, J.; et al. Preoperative radiotherapy with or without concurrent fluorouracil and leucovorin in T3-4 rectal cancers: Results of FFCD 9203. J. Clin. Oncol. 2006, 24, 4620–4625. [Google Scholar] [CrossRef]
- Andre, T.; Boni, C.; Mounedji-Boudiaf, L.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Zaninelli, M.; Clingan, P.; Bridgewater, J.; et al. Oxaliplatin, fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Dalerba, P.; Cho, R.W.; Clarke, M.F. Cancer stem cells: Models and concepts. Annu. Rev. Med. 2007, 58, 267–284. [Google Scholar] [CrossRef]
- Sugihara, E.; Saya, H. Complexity of cancer stem cells. Int. J. Cancer 2013, 132, 1249–1259. [Google Scholar] [CrossRef]
- Jordan, C.T.; Guzman, M.L.; Noble, M. Cancer stem cells. N. Engl. J. Med. 2006, 355, 1253–1261. [Google Scholar] [CrossRef]
- Dalerba, P.; Dylla, S.J.; Park, I.K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, C.A.; Pollett, A.; Gallinger, S.; Dick, J.E. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature 2007, 445, 106–110. [Google Scholar] [CrossRef]
- Ricci-Vitiani, L.; Lombardi, D.G.; Pilozzi, E.; Biffoni, M.; Todaro, M.; Peschle, C.; De Maria, R. Identification and expansion of human colon-cancer-initiating cells. Nature 2007, 445, 111–115. [Google Scholar] [CrossRef]
- Levin, T.G.; Powell, A.E.; Davies, P.S.; Silk, A.D.; Dismuke, A.D.; Anderson, E.C.; Swain, J.R.; Wong, M.H. Characterization of the intestinal cancer stem cell marker CD166 in the human and mouse gastrointestinal tract. Gastroenterology 2010, 139, 2072–2082.e2075. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Fojo, T.; Bates, S. Tumour stem cells and drug resistance. Nat. Rev. Cancer 2005, 5, 275–284. [Google Scholar] [CrossRef]
- Oshima, N.; Yamada, Y.; Nagayama, S.; Kawada, K.; Hasegawa, S.; Okabe, H.; Sakai, Y.; Aoi, T. Induction of cancer stem cell properties in colon cancer cells by defined factors. PLoS ONE 2014, 9, e101735. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, G.J.; Saya, H. Therapeutic strategies targeting cancer stem cells. Cancer Sci. 2016, 107, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Yada, M.; Hatakeyama, S.; Kamura, T.; Nishiyama, M.; Tsunematsu, R.; Imaki, H.; Ishida, N.; Okumura, F.; Nakayama, K.; Nakayama, K.I. Phosphorylation-dependent degradation of c-Myc is mediated by the F-box protein Fbw7. EMBO J. 2004, 23, 2116–2125. [Google Scholar] [CrossRef] [Green Version]
- Koepp, D.M.; Schaefer, L.K.; Ye, X.; Keyomarsi, K.; Chu, C.; Harper, J.W.; Elledge, S.J. Phosphorylation-dependent ubiquitination of cyclin E by the SCFFbw7 ubiquitin ligase. Science 2001, 294, 173–177. [Google Scholar] [CrossRef]
- Inuzuka, H.; Shaik, S.; Onoyama, I.; Gao, D.; Tseng, A.; Maser, R.S.; Zhai, B.; Wan, L.; Gutierrez, A.; Lau, A.W.; et al. SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for ubiquitylation and destruction. Nature 2011, 471, 104–109. [Google Scholar] [CrossRef]
- Nakayama, K.I.; Nakayama, K. Ubiquitin ligases: cell-cycle control and cancer. Nat. Rev. Cancer 2006, 6, 369–381. [Google Scholar] [CrossRef]
- Welcker, M.; Clurman, B.E. FBW7 ubiquitin ligase: A tumour suppressor at the crossroads of cell division, growth and differentiation. Nat. Rev. Cancer 2008, 8, 83–93. [Google Scholar] [CrossRef]
- Wertz, I.E.; Kusam, S.; Lam, C.; Okamoto, T.; Sandoval, W.; Anderson, D.J.; Helgason, E.; Ernst, J.A.; Eby, M.; Liu, J.; et al. Sensitivity to antitubulin chemotherapeutics is regulated by MCL1 and FBW7. Nature 2011, 471, 110–114. [Google Scholar] [CrossRef]
- Takeishi, S.; Matsumoto, A.; Onoyama, I.; Naka, K.; Hirao, A.; Nakayama, K.I. Ablation of Fbxw7 eliminates leukemia-initiating cells by preventing quiescence. Cancer Cell 2013, 23, 347–361. [Google Scholar] [CrossRef]
- Izumi, D.; Ishimoto, T.; Miyake, K.; Eto, T.; Arima, K.; Kiyozumi, Y.; Uchihara, T.; Kurashige, J.; Iwatsuki, M.; Baba, Y.; et al. Colorectal Cancer Stem Cells Acquire Chemoresistance Through the Upregulation of F-Box/WD Repeat-Containing Protein 7 and the Consequent Degradation of c-Myc. Stem Cells 2017, 35, 2027–2036. [Google Scholar] [CrossRef]
- Takeishi, S.; Nakayama, K.I. To wake up cancer stem cells, or to let them sleep, that is the question. Cancer Sci. 2016, 107, 875–881. [Google Scholar] [CrossRef]
- Anderson, E.C.; Hessman, C.; Levin, T.G.; Monroe, M.M.; Wong, M.H. The role of colorectal cancer stem cells in metastatic disease and therapeutic response. Cancers (Basel) 2011, 3, 319–339. [Google Scholar] [CrossRef]
- Dalerba, P.; Kalisky, T.; Sahoo, D.; Rajendran, P.S.; Rothenberg, M.E.; Leyrat, A.A.; Sim, S.; Okamoto, J.; Johnston, D.M.; Qian, D.; et al. Single-cell dissection of transcriptional heterogeneity in human colon tumors. Nat. Biotechnol. 2011, 29, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Fukushima, H.; Gao, D.; Inuzuka, H.; Wan, L.; Lau, A.W.; Liu, P.; Wei, W. The two faces of FBW7 in cancer drug resistance. Bioessays 2011, 33, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Yokobori, T.; Yokoyama, Y.; Mogi, A.; Endoh, H.; Altan, B.; Kosaka, T.; Yamaki, E.; Yajima, T.; Tomizawa, K.; Azuma, Y.; et al. FBXW7 mediates chemotherapeutic sensitivity and prognosis in NSCLCs. Mol. Cancer Res. 2014, 12, 32–37. [Google Scholar] [CrossRef]
- Ye, M.; Zhang, Y.; Zhang, X.; Zhang, J.; Jing, P.; Cao, L.; Li, N.; Li, X.; Yao, L. Targeting FBW7 as a Strategy to Overcome Resistance to Targeted Therapy in Non-Small Cell Lung Cancer. Cancer Res. 2017, 77, 3527–3539. [Google Scholar] [CrossRef]
- Arita, H.; Nagata, M.; Yoshida, R.; Matsuoka, Y.; Hirosue, A.; Kawahara, K.; Sakata, J.; Nakashima, H.; Kojima, T.; Toya, R.; et al. FBXW7 expression affects the response to chemoradiotherapy and overall survival among patients with oral squamous cell carcinoma: A single-center retrospective study. Tumour Biol. 2017, 39, 1010428317731771. [Google Scholar] [CrossRef]
- Gombodorj, N.; Yokobori, T.; Tanaka, N.; Suzuki, S.; Kuriyama, K.; Kumakura, Y.; Yoshida, T.; Sakai, M.; Sohda, M.; Baatar, S.; et al. Correlation between high FBXW7 expression in pretreatment biopsy specimens and good response to chemoradiation therapy in patients with locally advanced esophageal cancer: A retrospective study. J. Surg. Oncol. 2018, 118, 101–108. [Google Scholar] [CrossRef]
- Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [CrossRef]
- Japanese Society for Cancer of the Colon and Rectum. Japanese Classification of Colorectal Carcinoma, 8th ed.; Kanehara & Co., LTD: Tokyo, Japan, 2013; p. 34. [Google Scholar]
- Itatani, Y.; Kawada, K.; Fujishita, T.; Kakizaki, F.; Hirai, H.; Matsumoto, T.; Iwamoto, M.; Inamoto, S.; Hatano, E.; Hasegawa, S.; et al. Loss of SMAD4 from colorectal cancer cells promotes CCL15 expression to recruit CCR1+ myeloid cells and facilitate liver metastasis. Gastroenterology 2013, 145, 1064–1075.e1011. [Google Scholar] [CrossRef]
- Westrate, L.M.; Sayfie, A.D.; Burgenske, D.M.; MacKeigan, J.P. Persistent mitochondrial hyperfusion promotes G2/M accumulation and caspase-dependent cell death. PLoS ONE 2014, 9, e91911. [Google Scholar] [CrossRef]
- Miyoshi, H.; Maekawa, H.; Kakizaki, F.; Yamaura, T.; Kawada, K.; Sakai, Y.; Taketo, M.M. An improved method for culturing patient-derived colorectal cancer spheroids. Oncotarget 2018, 9, 21950–21964. [Google Scholar] [CrossRef] [Green Version]
- Deguchi, Y.; Okabe, H.; Oshima, N.; Hisamori, S.; Minamiguchi, S.; Muto, M.; Sakai, Y. PTEN loss is associated with a poor response to trastuzumab in HER2-overexpressing gastroesophageal adenocarcinoma. Gastric. Cancer 2017, 20, 416–427. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | FBXW7 Expression | |||
|---|---|---|---|---|
| Total | High | Low | p Value | |
| n = 55 | n = 21 | n = 34 | ||
| Age | 0.464 | |||
| mean ± SD | 60.6 ± 13.4 | 62.6 ± 7.0 | ||
| range | 26–79 | 50–73 | ||
| Gender | 0.956 | |||
| Male | 43 | 16 | 27 | |
| Female | 12 | 5 | 7 | |
| Location | 0.684 | |||
| Right hemicolon | 2 | 2 | 0 | |
| Left hemicolon | 1 | 1 | 0 | |
| Sigmoid colon | 7 | 3 | 4 | |
| Rectum | 45 | 15 | 30 | |
| Chemotherapy regimen | 0.98 | |||
| L-OHP base | 40 | 16 | 24 | |
| CPT-11 base | 11 | 4 | 7 | |
| others | 4 | 1 | 3 | |
| Molecular target drug | 0.153 | |||
| with | 29 | 8 | 21 | |
| without | 26 | 13 | 13 | |
| Radiation therapy | 0.502 | |||
| RT (+) | 11 | 3 | 8 | |
| RT (−) | 44 | 18 | 26 | |
| Histology (biopsy) | 0.903 | |||
| well, moderate | 49 | 19 | 30 | |
| por, sig, muc | 5 | 1 | 4 | |
| others | 1 | 1 | 0 | |
| clinical N status | 1 | |||
| cN (+) | 51 | 20 | 31 | |
| cN (−) | 4 | 1 | 3 | |
| clinical M status | 0.19 | |||
| cM (+) | 19 | 10 | 9 | |
| cM (−) | 36 | 11 | 25 | |
| clinical Stage | 0.462 | |||
| II | 2 | 0 | 2 | |
| III | 34 | 11 | 23 | |
| IV | 19 | 10 | 9 | |
| Pathological therapeutic effect | 0.019 | |||
| grade 0, 1 | 36 | 18 | 18 | |
| grade 2, 3 | 19 | 3 | 16 | |
| No | Biopsy Specimen | Resected Specimen | ΔValue * | ||||
|---|---|---|---|---|---|---|---|
| Percentage | Intensity | Score | Percentage | Intensity | Score | ||
| 1 | 1 | 1 | 1 | 3 | 2 | 6 | 5 |
| 2 | 1 | 1 | 1 | 3 | 2 | 6 | 5 |
| 3 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 4 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 5 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 6 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 7 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 8 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 9 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 10 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 11 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 12 | 1 | 2 | 2 | 3 | 2 | 6 | 4 |
| 13 | 2 | 1 | 2 | 3 | 2 | 6 | 4 |
| 14 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 15 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 16 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 17 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 18 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 19 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 20 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 21 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 22 | 3 | 1 | 3 | 3 | 2 | 6 | 3 |
| 23 | 2 | 2 | 4 | 3 | 2 | 6 | 2 |
| 24 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 25 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 26 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 27 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 28 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 29 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 30 | 1 | 1 | 1 | 3 | 1 | 3 | 2 |
| 31 | 1 | 1 | 1 | 2 | 1 | 2 | 1 |
| 32 | 1 | 1 | 1 | 2 | 1 | 2 | 1 |
| 33 | 2 | 1 | 2 | 3 | 1 | 3 | 1 |
| 34 | 2 | 1 | 2 | 3 | 1 | 3 | 1 |
| 35 | 1 | 2 | 2 | 3 | 1 | 3 | 1 |
| 36 | 2 | 1 | 2 | 3 | 1 | 3 | 1 |
| 37 | 2 | 1 | 2 | 3 | 1 | 3 | 1 |
| 38 | 2 | 1 | 2 | 3 | 1 | 3 | 1 |
| 39 | 2 | 1 | 2 | 3 | 1 | 3 | 1 |
| 40 | 3 | 2 | 6 | 3 | 2 | 6 | 0 |
| 41 | 3 | 1 | 3 | 3 | 1 | 3 | 0 |
| 42 | 3 | 1 | 3 | 3 | 1 | 3 | 0 |
| 43 | 3 | 2 | 6 | 3 | 2 | 6 | 0 |
| 44 | 3 | 1 | 3 | 3 | 1 | 3 | 0 |
| 45 | 3 | 2 | 6 | 3 | 2 | 6 | 0 |
| 46 | 3 | 2 | 6 | 3 | 2 | 6 | 0 |
| 47 | 3 | 2 | 6 | 3 | 2 | 6 | 0 |
| 48 | 3 | 1 | 3 | 3 | 1 | 3 | 0 |
| 49 | 3 | 1 | 3 | 3 | 1 | 3 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honma, S.; Hisamori, S.; Nishiuchi, A.; Itatani, Y.; Obama, K.; Shimono, Y.; Sakai, Y. F-Box/WD Repeat Domain-Containing 7 Induces Chemotherapy Resistance in Colorectal Cancer Stem Cells. Cancers 2019, 11, 635. https://doi.org/10.3390/cancers11050635
Honma S, Hisamori S, Nishiuchi A, Itatani Y, Obama K, Shimono Y, Sakai Y. F-Box/WD Repeat Domain-Containing 7 Induces Chemotherapy Resistance in Colorectal Cancer Stem Cells. Cancers. 2019; 11(5):635. https://doi.org/10.3390/cancers11050635
Chicago/Turabian StyleHonma, Shusaku, Shigeo Hisamori, Aya Nishiuchi, Yoshiro Itatani, Kazutaka Obama, Yohei Shimono, and Yoshiharu Sakai. 2019. "F-Box/WD Repeat Domain-Containing 7 Induces Chemotherapy Resistance in Colorectal Cancer Stem Cells" Cancers 11, no. 5: 635. https://doi.org/10.3390/cancers11050635