Erbb4 Is Required for Cerebellar Development and Malignant Phenotype of Medulloblastoma

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Models
2.2. Cell Culture
2.3. CGNP Isolation and Culture
2.4. Viral Infections
2.5. Tissue Immunofluorescence
2.6. Cell Immunofluorescence
2.7. Brain Immunohistochemistry
2.8. Tumor Immunohistochemistry
2.9. RNAscope
2.10. Migration Assay
2.11. Cell Viability MTT Assay
2.12. RNA Extraction, Reverse Transcription, and Gene Expression
2.13. Western Blot
2.14. Colony Formation Assay
2.15. Oncosphere Formation Assay
2.16. Cell Cycle
2.17. Cell Apoptosis Assay
2.18. In Vivo Carcinogenesis
2.19. Microarray Analysis
2.20. Data Analysis
3. Results
3.1. ERBB4 is Expressed in The Inner Part of the External Germinal Layer During Cerebellum Development
3.2. ERBB4 Is Expressed in Culture CGNPs, and Its Activation Protects Cells from an Apoptotic Stimulus In Vitro
3.3. ERBB4 Expression Is Higher in Group 4 Medulloblastomas, and Its High Expression Is Associated with Poor Clinical Outcome
3.4. ERBB4 Knock-down Inhibits Cell Viability and Activates Apoptosis in Human Medulloblastoma Cells In Vitro and In Vivo
3.5. MBSCs Express High Levels of ERBB4 and its Knock-Down Inhibits MBSCs Activity
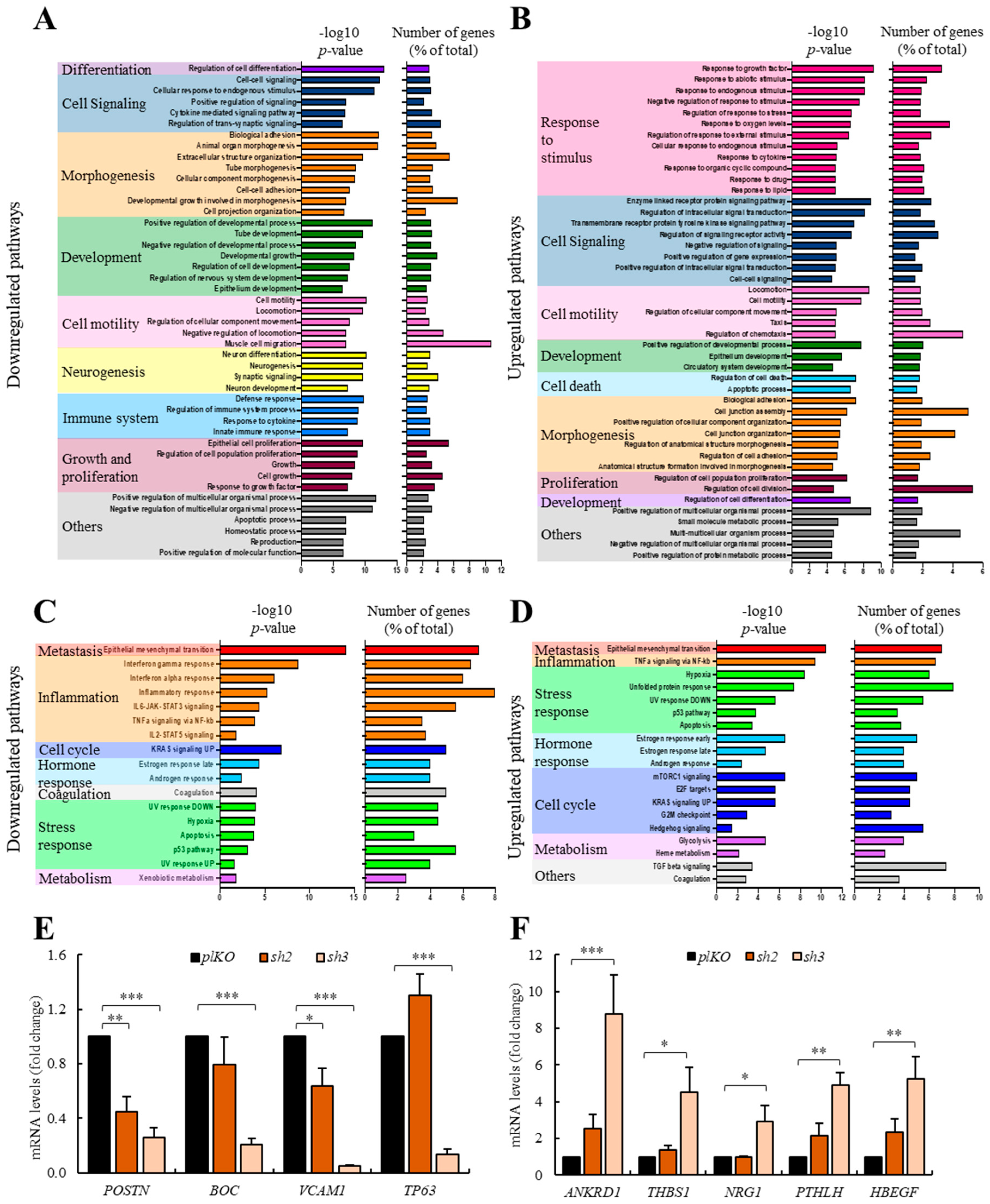
3.6. ERBB4 Knock-Down Alters Multiple Processes and Pathways in Medulloblastoma Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bartlett, F.; Kortmann, R.; Saran, F. Medulloblastoma. Clin. Oncol. R Coll. Radiol. 2013, 25, 36–45. [Google Scholar] [CrossRef]
- Butts, T.; Green, M.J.; Wingate, R.J. Development of the cerebellum: Simple steps to make a ‘little brain’. Development 2014, 141, 4031–4041. [Google Scholar] [CrossRef] [Green Version]
- Rakic, P. Neuron-glia relationship during granule cell migration in developing cerebellar cortex. A golgi and electronmicroscopic study in macacus rhesus. J. Comp. Neurol. 1971, 141, 283–312. [Google Scholar] [CrossRef]
- Grimmer, M.R.; Weiss, W.A. Childhood tumors of the nervous system as disorders of normal development. Curr. Opin. Pediatr. 2006, 18, 634–638. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 world health organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [Green Version]
- Coluccia, D.; Figuereido, C.; Isik, S.; Smith, C.; Rutka, J.T. Medulloblastoma: Tumor biology and relevance to treatment and prognosis paradigm. Curr. Neurol. Neurosci. Rep. 2016, 16, 43. [Google Scholar] [CrossRef]
- Musial-Bright, L.; Fengler, R.; Henze, G.; Hernaiz Driever, P. Carboplatin and ototoxicity: Hearing loss rates among survivors of childhood medulloblastoma. Childs Nerv. Syst. 2011, 27, 407–413. [Google Scholar] [CrossRef]
- Kadota, R.P.; Mahoney, D.H.; Doyle, J.; Duerst, R.; Friedman, H.; Holmes, E.; Kun, L.; Zhou, T.; Pollack, I.F. Dose intensive melphalan and cyclophosphamide with autologous hematopoietic stem cells for recurrent medulloblastoma or germinoma. Pediatr. Blood Cancer 2008, 51, 675–678. [Google Scholar] [CrossRef] [Green Version]
- Saury, J.M.; Emanuelson, I. Cognitive consequences of the treatment of medulloblastoma among children. Pediatr. Neurol. 2011, 44, 21–30. [Google Scholar] [CrossRef]
- Gershon, T.R.; Becher, O.J. Medulloblastoma: Therapy and biologic considerations. Curr. Neurol. Neurosci. Rep. 2006, 6, 200–206. [Google Scholar] [CrossRef]
- Taylor, M.D.; Northcott, P.A.; Korshunov, A.; Remke, M.; Cho, Y.J.; Clifford, S.C.; Eberhart, C.G.; Parsons, D.W.; Rutkowski, S.; Gajjar, A.; et al. Molecular subgroups of medulloblastoma: The current consensus. Acta Neuropathol. 2012, 123, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar]
- Clarke, M.F.; Dick, J.E.; Dirks, P.B.; Eaves, C.J.; Jamieson, C.H.; Jones, D.L.; Visvader, J.; Weissman, I.L.; Wahl, G.M. Cancer stem cells--perspectives on current status and future directions: Aacr workshop on cancer stem cells. Cancer Res. 2006, 66, 9339–9344. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Kumar, V.; McGuire, T.; Coulter, D.W.; Sharp, J.G.; Mahato, R.I. Challenges and recent advances in medulloblastoma therapy. Trends Pharmacol. Sci. 2017, 38, 1061–1084. [Google Scholar] [CrossRef]
- Al Moustafa, A.E.; Achkhar, A.; Yasmeen, A. Egf-receptor signaling and epithelial-mesenchymal transition in human carcinomas. Front Biosci. 2012, 4, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Appert-Collin, A.; Hubert, P.; Cremel, G.; Bennasroune, A. Role of erbb receptors in cancer cell migration and invasion. Front. Pharmacol. 2015, 6, 283. [Google Scholar] [CrossRef] [Green Version]
- Donoghue, J.F.; Kerr, L.T.; Alexander, N.W.; Greenall, S.A.; Longano, A.B.; Gottardo, N.G.; Wang, R.; Tabar, V.; Adams, T.E.; Mischel, P.S.; et al. Activation of erbb4 in glioblastoma can contribute to increased tumorigenicity and influence therapeutic response. Cancers 2018, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Yang, X.; Chu, L.; Zhao, K.; Chen, H.; Xiang, J.; Zhang, Y.; Li, H.; Zhao, W.; Sun, M.; et al. Prognostic value of egfr family expression in lymph node-negative esophageal squamous cell carcinoma patients. Pathol. Res. Pract. 2018, 214, 1017–1023. [Google Scholar] [CrossRef]
- Saglam, O.; Xiong, Y.; Marchion, D.C.; Strosberg, C.; Wenham, R.M.; Johnson, J.J.; Saeed-Vafa, D.; Cubitt, C.; Hakam, A.; Magliocco, A.M. Erbb4 expression in ovarian serous carcinoma resistant to platinum-based therapy. Cancer Control 2017, 24, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Jung, H.H.; Do, I.G.; Bae, S.; Lee, S.K.; Kim, S.W.; Lee, J.E.; Nam, S.J.; Ahn, J.S.; Park, Y.H.; et al. Prognostic value of erbb4 expression in patients with triple negative breast cancer. BMC Cancer 2016, 16, 138. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, K.; Yonezawa, M.; Tatsuguchi, A.; Shinji, S.; Gudis, K.; Tanaka, S.; Fujimori, S.; Sakamoto, C. Localization of phosphorylated erbb1-4 and heregulin in colorectal cancer. BMC Cancer 2014, 14, 863. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, L.; Ni, H.; Sun, L.; Jiao, W.; Chen, L.; Zhou, Q.; Shen, T.; Cui, H.; Gao, T.; et al. Erbb4 acts as a suppressor in the development of hepatocellular carcinoma. Carcinogenesis 2017, 38, 465–473. [Google Scholar] [CrossRef]
- Wang, J.; Yin, J.; Yang, Q.; Ding, F.; Chen, X.; Li, B.; Tian, X. Human epidermal growth factor receptor 4 (her4) is a favorable prognostic marker of breast cancer: A systematic review and meta-analysis. Oncotarget 2016, 7, 76693–76703. [Google Scholar] [CrossRef] [Green Version]
- Machleidt, A.; Buchholz, S.; Diermeier-Daucher, S.; Zeman, F.; Ortmann, O.; Brockhoff, G. The prognostic value of her4 receptor isoform expression in triple-negative and her2 positive breast cancer patients. BMC Cancer 2013, 13, 437. [Google Scholar] [CrossRef] [Green Version]
- Hollmen, M.; Elenius, K. Potential of erbb4 antibodies for cancer therapy. Future Oncol. 2010, 6, 37–53. [Google Scholar] [CrossRef]
- Zhao, W.Y.; Zhuang, C.; Xu, J.; Wang, M.; Zhang, Z.Z.; Tu, L.; Wang, C.J.; Ling, T.L.; Cao, H.; Zhang, Z.G. Her4 is a novel prognostic biomarker in gastrointestinal stromal tumor specifically originated from stomach. Am. J. Cancer Res. 2014, 4, 838–849. [Google Scholar]
- Hua, Y.; Gorshkov, K.; Yang, Y.; Wang, W.; Zhang, N.; Hughes, D.P. Slow down to stay alive: Her4 protects against cellular stress and confers chemoresistance in neuroblastoma. Cancer 2012, 118, 5140–5154. [Google Scholar] [CrossRef]
- Hegde, G.V.; de la Cruz, C.C.; Chiu, C.; Alag, N.; Schaefer, G.; Crocker, L.; Ross, S.; Goldenberg, D.; Merchant, M.; Tien, J.; et al. Blocking nrg1 and other ligand-mediated her4 signaling enhances the magnitude and duration of the chemotherapeutic response of non-small cell lung cancer. Sci. Transl. Med. 2013, 5, 171ra118. [Google Scholar] [CrossRef]
- Bao, J.; Wolpowitz, D.; Role, L.W.; Talmage, D.A. Back signaling by the nrg-1 intracellular domain. J. Cell Biol. 2003, 161, 1133–1141. [Google Scholar] [CrossRef] [Green Version]
- Birchmeier, C. Erbb receptors and the development of the nervous system. Exp. Cell Res. 2009, 315, 611–618. [Google Scholar] [CrossRef]
- Ghashghaei, H.T.; Weber, J.; Pevny, L.; Schmid, R.; Schwab, M.H.; Lloyd, K.C.; Eisenstat, D.D.; Lai, C.; Anton, E.S. The role of neuregulin-erbb4 interactions on the proliferation and organization of cells in the subventricular zone. Proc. Natl. Acad. Sci. USA 2006, 103, 1930–1935. [Google Scholar] [CrossRef] [Green Version]
- Gerecke, K.M.; Wyss, J.M.; Carroll, S.L. Neuregulin-1beta induces neurite extension and arborization in cultured hippocampal neurons. Mol. Cell. Neurosci. 2004, 27, 379–393. [Google Scholar] [CrossRef]
- Min, S.S.; An, J.; Lee, J.H.; Seol, G.H.; Im, J.H.; Kim, H.S.; Baik, T.K.; Woo, R.S. Neuregulin-1 prevents amyloid beta-induced impairment of long-term potentiation in hippocampal slices via erbb4. Neurosci. Lett. 2011, 505, 6–9. [Google Scholar] [CrossRef]
- Vullhorst, D.; Neddens, J.; Karavanova, I.; Tricoire, L.; Petralia, R.S.; McBain, C.J.; Buonanno, A. Selective expression of erbb4 in interneurons, but not pyramidal cells, of the rodent hippocampus. J. Neurosci. 2009, 29, 12255–12264. [Google Scholar] [CrossRef] [Green Version]
- Perez-Garcia, C.G. Erbb4 in laminated brain structures: A neurodevelopmental approach to schizophrenia. Front. Cell. Neurosci. 2015, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- Pinkas-Kramarski, R.; Eilam, R.; Alroy, I.; Levkowitz, G.; Lonai, P.; Yarden, Y. Differential expression of ndf/neuregulin receptors erbb-3 and erbb-4 and involvement in inhibition of neuronal differentiation. Oncogene 1997, 15, 2803–2815. [Google Scholar] [CrossRef] [Green Version]
- Elenius, K.; Corfas, G.; Paul, S.; Choi, C.J.; Rio, C.; Plowman, G.D.; Klagsbrun, M. A novel juxtamembrane domain isoform of her4/erbb4. Isoform-specific tissue distribution and differential processing in response to phorbol ester. J. Biol. Chem. 1997, 272, 26761–26768. [Google Scholar] [CrossRef] [Green Version]
- Rio, C.; Rieff, H.I.; Qi, P.; Khurana, T.S.; Corfas, G. Neuregulin and erbb receptors play a critical role in neuronal migration. Neuron 1997, 19, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Zeng, N.; Liu, L.; McCabe, M.G.; Jones, D.T.; Ichimura, K.; Collins, V.P. Real-time quantitative polymerase chain reaction (qpcr) analysis with fluorescence resonance energy transfer (fret) probes reveals differential expression of the four erbb4 juxtamembrane region variants between medulloblastoma and pilocytic astrocytoma. Neuropathol. Appl. Neurobiol. 2009, 35, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Gilbertson, R.J.; Perry, R.H.; Kelly, P.J.; Pearson, A.D.; Lunec, J. Prognostic significance of her2 and her4 coexpression in childhood medulloblastoma. Cancer Res. 1997, 57, 3272–3280. [Google Scholar]
- Bal, M.M.; Das Radotra, B.; Srinivasan, R.; Sharma, S.C. Expression of c-erbb-4 in medulloblastoma and its correlation with prognosis. Histopathology 2006, 49, 92–93. [Google Scholar] [CrossRef]
- De la Rosa, J.; Saenz Antonanzas, A.; Shahi, M.H.; Melendez, B.; Rey, J.A.; Castresana, J.S. Laminin-adherent versus suspension-non-adherent cell culture conditions for the isolation of cancer stem cells in the daoy medulloblastoma cell line. Tumor Biol. 2016, 37, 12359–12370. [Google Scholar] [CrossRef]
- Garcia, I.; Aldaregia, J.; Marjanovic Vicentic, J.; Aldaz, P.; Moreno-Cugnon, L.; Torres-Bayona, S.; Carrasco-Garcia, E.; Garros-Regulez, L.; Egana, L.; Rubio, A.; et al. Oncogenic activity of sox1 in glioblastoma. Sci. Rep. 2017, 7, 46575. [Google Scholar] [CrossRef]
- Garros-Regulez, L.; Aldaz, P.; Arrizabalaga, O.; Moncho-Amor, V.; Carrasco-Garcia, E.; Manterola, L.; Moreno-Cugnon, L.; Barrena, C.; Villanua, J.; Ruiz, I.; et al. Mtor inhibition decreases sox2-sox9 mediated glioma stem cell activity and temozolomide resistance. Expert Opin. Ther. Targets 2016, 20, 393–405. [Google Scholar] [CrossRef]
- Crowther, A.J.; Gama, V.; Bevilacqua, A.; Chang, S.X.; Yuan, H.; Deshmukh, M.; Gershon, T.R. Tonic activation of bax primes neural progenitors for rapid apoptosis through a mechanism preserved in medulloblastoma. J. Neurosci. 2013, 33, 18098–18108. [Google Scholar] [CrossRef] [Green Version]
- Kornblum, H.I.; Zurcher, S.D.; Werb, Z.; Derynck, R.; Seroogy, K.B. Multiple trophic actions of heparin-binding epidermal growth factor (hb-egf) in the central nervous system. Eur. J. Neurosci. 1999, 11, 3236–3246. [Google Scholar] [CrossRef]
- Gilbertson, R.J.; Clifford, S.C.; MacMeekin, W.; Meekin, W.; Wright, C.; Perry, R.H.; Kelly, P.; Pearson, A.D.; Lunec, J. Expression of the erbb-neuregulin signaling network during human cerebellar development: Implications for the biology of medulloblastoma. Cancer Res. 1998, 58, 3932–3941. [Google Scholar]
- Ben-Arie, N.; Bellen, H.J.; Armstrong, D.L.; McCall, A.E.; Gordadze, P.R.; Guo, Q.; Matzuk, M.M.; Zoghbi, H.Y. Math1 is essential for genesis of cerebellar granule neurons. Nature 1997, 390, 169–172. [Google Scholar] [CrossRef]
- Kirsch, L.; Liscovitch, N.; Chechik, G. Localizing genes to cerebellar layers by classifying ish images. PLoS Comput. Biol. 2012, 8, e1002790. [Google Scholar] [CrossRef] [Green Version]
- Read, T.A.; Fogarty, M.P.; Markant, S.L.; McLendon, R.E.; Wei, Z.; Ellison, D.W.; Febbo, P.G.; Wechsler-Reya, R.J. Identification of cd15 as a marker for tumor-propagating cells in a mouse model of medulloblastoma. Cancer Cell 2009, 15, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Vanner, R.J.; Remke, M.; Gallo, M.; Selvadurai, H.J.; Coutinho, F.; Lee, L.; Kushida, M.; Head, R.; Morrissy, S.; Zhu, X.; et al. Quiescent sox2(+) cells drive hierarchical growth and relapse in sonic hedgehog subgroup medulloblastoma. Cancer Cell 2014, 26, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Zhang, C.; Chen, D.; Chen, S.; Zheng, H. Microrna-98-hmga2-postn signal pathway reverses epithelial-to-mesenchymal transition in laryngeal squamous cell carcinoma. Biomed. Pharmacother. 2019, 117, 108998. [Google Scholar] [CrossRef]
- Mille, F.; Tamayo-Orrego, L.; Levesque, M.; Remke, M.; Korshunov, A.; Cardin, J.; Bouchard, N.; Izzi, L.; Kool, M.; Northcott, P.A.; et al. The shh receptor boc promotes progression of early medulloblastoma to advanced tumors. Dev. Cell 2014, 31, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Liang, L.; Aiken, C.; McClelland, R.; Morrison, L.C.; Tatari, N.; Remke, M.; Ramaswamy, V.; Issaivanan, M.; Ryken, T.; Del Bigio, M.R.; et al. Characterization of novel biomarkers in selecting for subtype specific medulloblastoma phenotypes. Oncotarget 2015, 6, 38881–38900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forster, N.; Saladi, S.V.; van Bragt, M.; Sfondouris, M.E.; Jones, F.E.; Li, Z.; Ellisen, L.W. Basal cell signaling by p63 controls luminal progenitor function and lactation via nrg1. Dev. Cell 2014, 28, 147–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, A.P.; Traum, A.; Boettger, T.; Hackstein, H.; Richter, A.M.; Dammann, R.H. The tumor suppressor rassf1a induces the yap1 target gene ankrd1 that is epigenetically inactivated in human cancers and inhibits tumor growth. Oncotarget 2017, 8, 88437–88452. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Picard, D.; Ra, Y.S.; Li, M.; Northcott, P.A.; Hu, Y.; Stearns, D.; Hawkins, C.; Taylor, M.D.; Rutka, J.; et al. Silencing of thrombospondin-1 is critical for myc-induced metastatic phenotypes in medulloblastoma. Cancer Res. 2010, 70, 8199–8210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, Y.; Katoh, M. Hedgehog target genes: Mechanisms of carcinogenesis induced by aberrant hedgehog signaling activation. Curr. Mol. Med. 2009, 9, 873–886. [Google Scholar] [CrossRef]
- Leto, K.; Arancillo, M.; Becker, E.B.; Buffo, A.; Chiang, C.; Ding, B.; Dobyns, W.B.; Dusart, I.; Haldipur, P.; Hatten, M.E.; et al. Consensus paper: Cerebellar development. Cerebellum 2016, 15, 789–828. [Google Scholar] [CrossRef]
- Zurawel, R.H.; Chiappa, S.A.; Allen, C.; Raffel, C. Sporadic medulloblastomas contain oncogenic beta-catenin mutations. Cancer Res. 1998, 58, 896–899. [Google Scholar]
- Dey, J.; Ditzler, S.; Knoblaugh, S.E.; Hatton, B.A.; Schelter, J.M.; Cleary, M.A.; Mecham, B.; Rorke-Adams, L.B.; Olson, J.M. A distinct smoothened mutation causes severe cerebellar developmental defects and medulloblastoma in a novel transgenic mouse model. Mol. Cell. Biol. 2012, 32, 4104–4115. [Google Scholar] [CrossRef] [Green Version]
- Mei, L.; Xiong, W.C. Neuregulin 1 in neural development, synaptic plasticity and schizophrenia. Nat. Rev. Neurosci. 2008, 9, 437–452. [Google Scholar] [CrossRef]
- Fornasari, B.E.; El Soury, M.; De Marchis, S.; Perroteau, I.; Geuna, S.; Gambarotta, G. Neuregulin1 alpha activates migration of neuronal progenitors expressing erbb4. Mol. Cell. Neurosci. 2016, 77, 87–94. [Google Scholar] [CrossRef]
- Ortega, M.C.; Bribian, A.; Peregrin, S.; Gil, M.T.; Marin, O.; de Castro, F. Neuregulin-1/erbb4 signaling controls the migration of oligodendrocyte precursor cells during development. Exp. Neurol. 2012, 235, 610–620. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, C.; Lai, C.; Riese, D.J., 2nd; Diamonti, A.J.; Cantley, L.C.; Carraway, K.L., 3rd. Ligand discrimination in signaling through an erbb4 receptor homodimer. J. Biol. Chem. 2000, 275, 19803–19807. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, R.; Miller, T.A.; Kuramochi, Y.; Frantz, S.; Kim, Y.D.; Marchionni, M.A.; Kelly, R.A.; Sawyer, D.B. Neuregulin-1 protects ventricular myocytes from anthracycline-induced apoptosis via erbb4-dependent activation of pi3-kinase/akt. J. Mol. Cell. Cardiol. 2003, 35, 1473–1479. [Google Scholar] [CrossRef]
- Yan, F.; Tan, X.; Wan, W.; Dixon, B.J.; Fan, R.; Enkhjargal, B.; Li, Q.; Zhang, J.; Chen, G.; Zhang, J.H. Erbb4 protects against neuronal apoptosis via activation of yap/pik3cb signaling pathway in a rat model of subarachnoid hemorrhage. Exp. Neurol. 2017, 297, 92–100. [Google Scholar] [CrossRef]
- Williams, C.S.; Bernard, J.K.; Demory Beckler, M.; Almohazey, D.; Washington, M.K.; Smith, J.J.; Frey, M.R. Erbb4 is over-expressed in human colon cancer and enhances cellular transformation. Carcinogenesis 2015, 36, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Forget, A.; Martignetti, L.; Puget, S.; Calzone, L.; Brabetz, S.; Picard, D.; Montagud, A.; Liva, S.; Sta, A.; Dingli, F.; et al. Aberrant erbb4-src signaling as a hallmark of group 4 medulloblastoma revealed by integrative phosphoproteomic profiling. Cancer Cell 2018, 34, 379–395.e7. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aldaregia, J.; Errarte, P.; Olazagoitia-Garmendia, A.; Gimeno, M.; Uriz, J.J.; Gershon, T.R.; Garcia, I.; Matheu, A. Erbb4 Is Required for Cerebellar Development and Malignant Phenotype of Medulloblastoma. Cancers 2020, 12, 997. https://doi.org/10.3390/cancers12040997
Aldaregia J, Errarte P, Olazagoitia-Garmendia A, Gimeno M, Uriz JJ, Gershon TR, Garcia I, Matheu A. Erbb4 Is Required for Cerebellar Development and Malignant Phenotype of Medulloblastoma. Cancers. 2020; 12(4):997. https://doi.org/10.3390/cancers12040997
Chicago/Turabian StyleAldaregia, Juncal, Peio Errarte, Ane Olazagoitia-Garmendia, Marian Gimeno, Jose Javier Uriz, Timothy R. Gershon, Idoia Garcia, and Ander Matheu. 2020. "Erbb4 Is Required for Cerebellar Development and Malignant Phenotype of Medulloblastoma" Cancers 12, no. 4: 997. https://doi.org/10.3390/cancers12040997