
Inhibition of Human Lung Cancer Cell Proliferation and Survival by Post-Exercise Serum Is Associated with the Inhibition of Akt, mTOR, p70 S6K, and Erk1/2



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
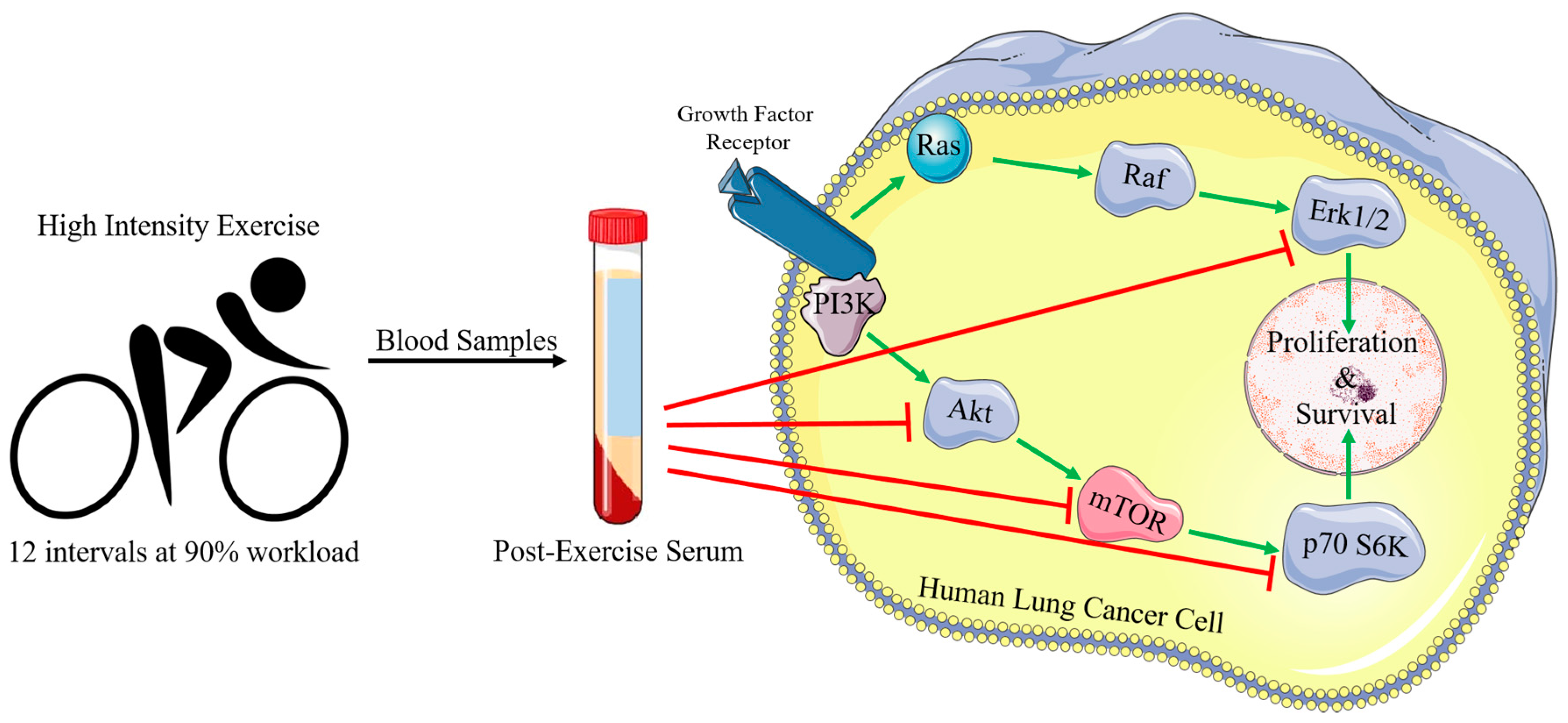
:1. Introduction
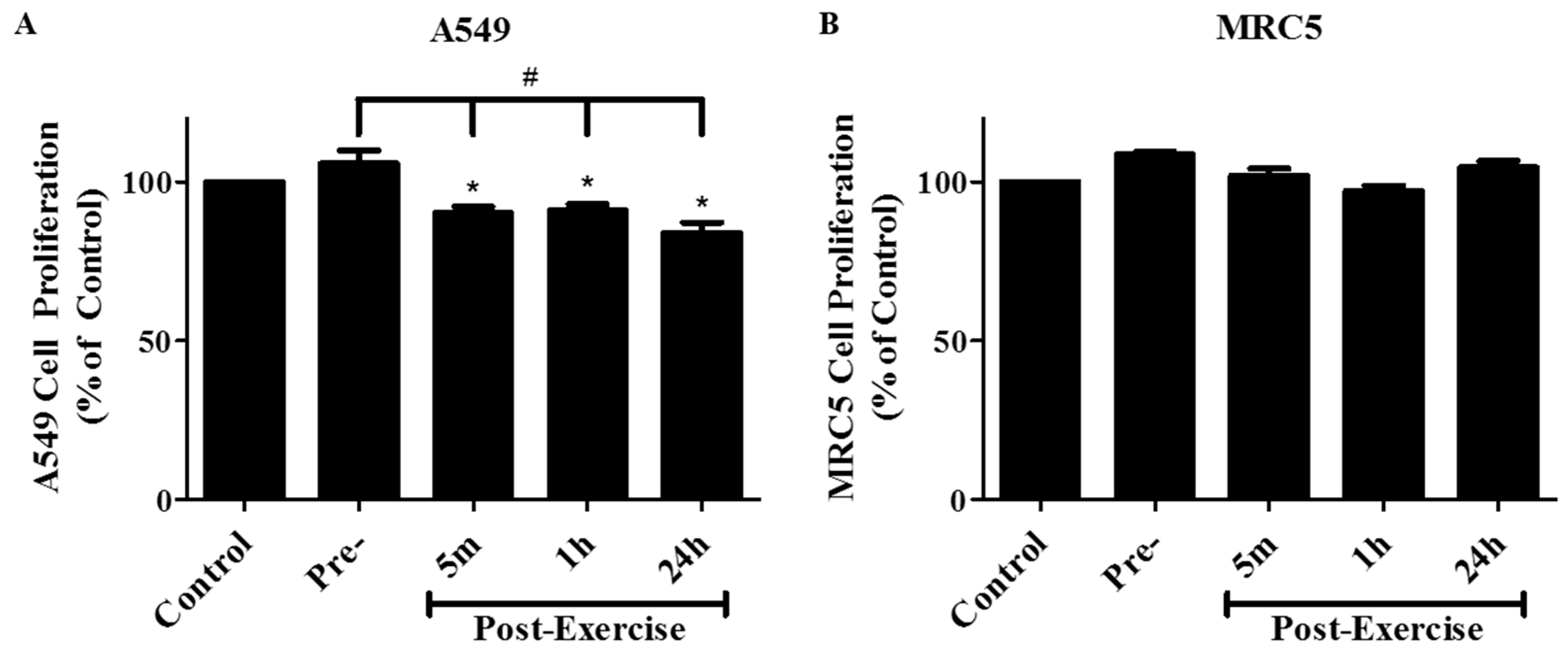
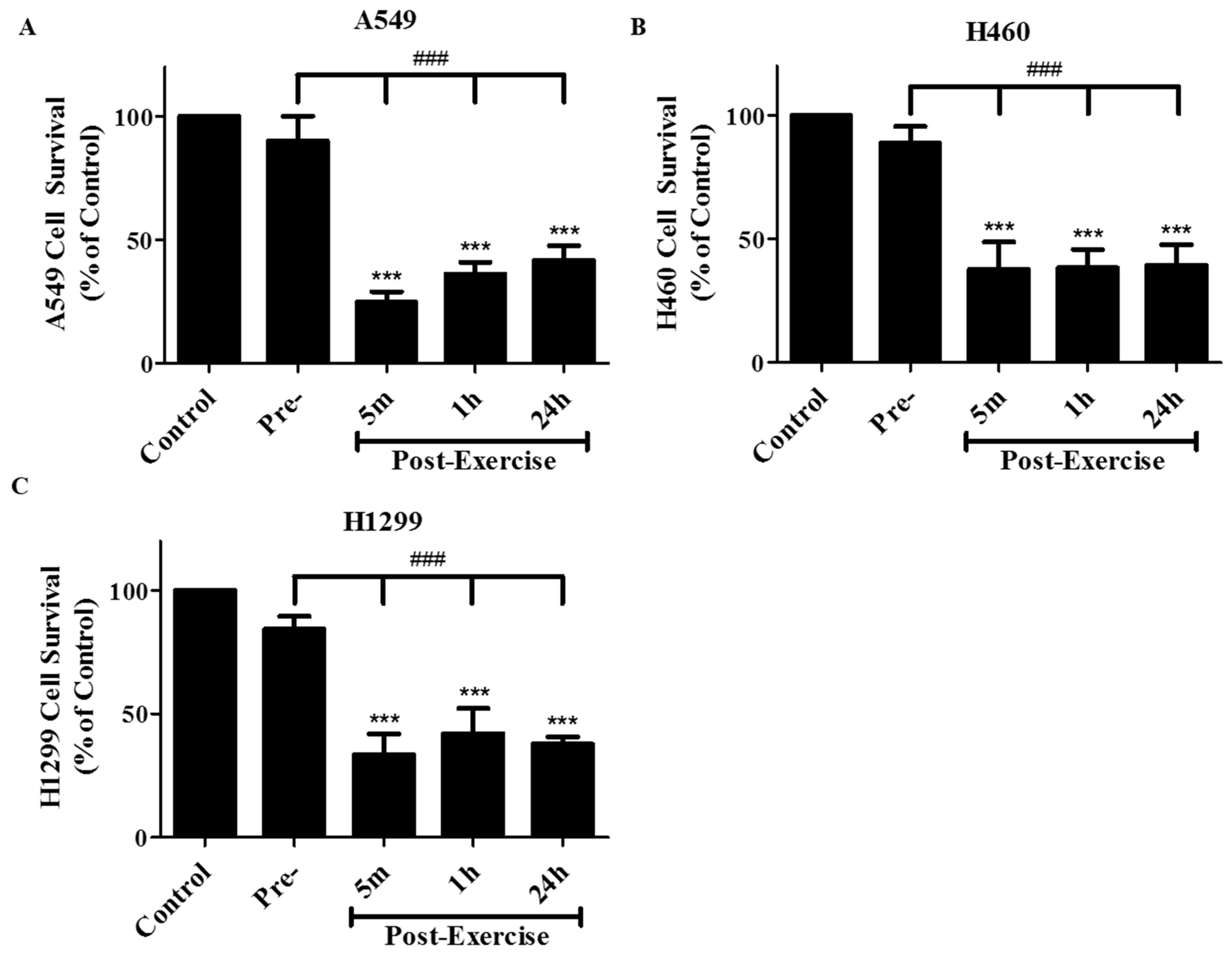
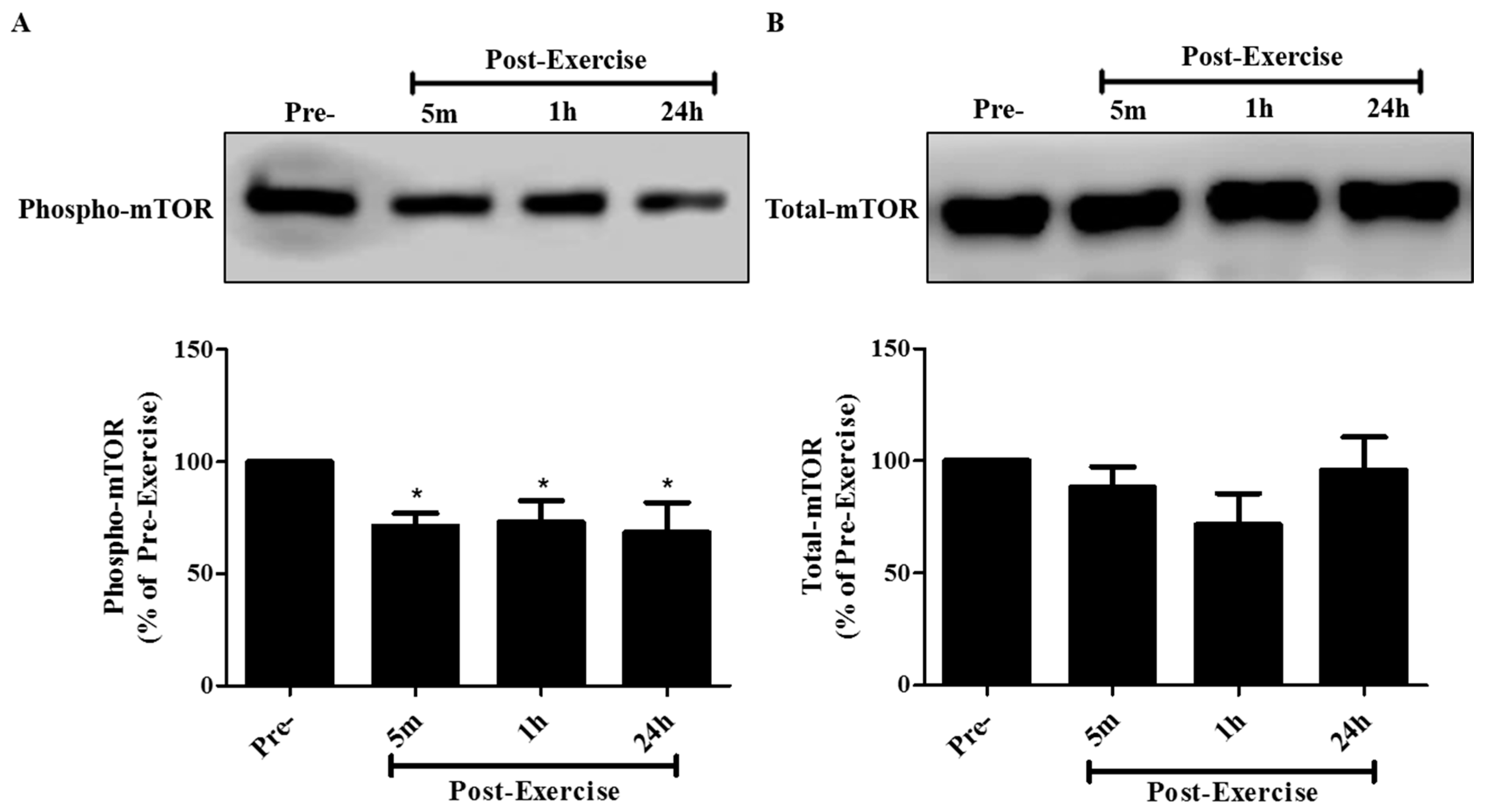
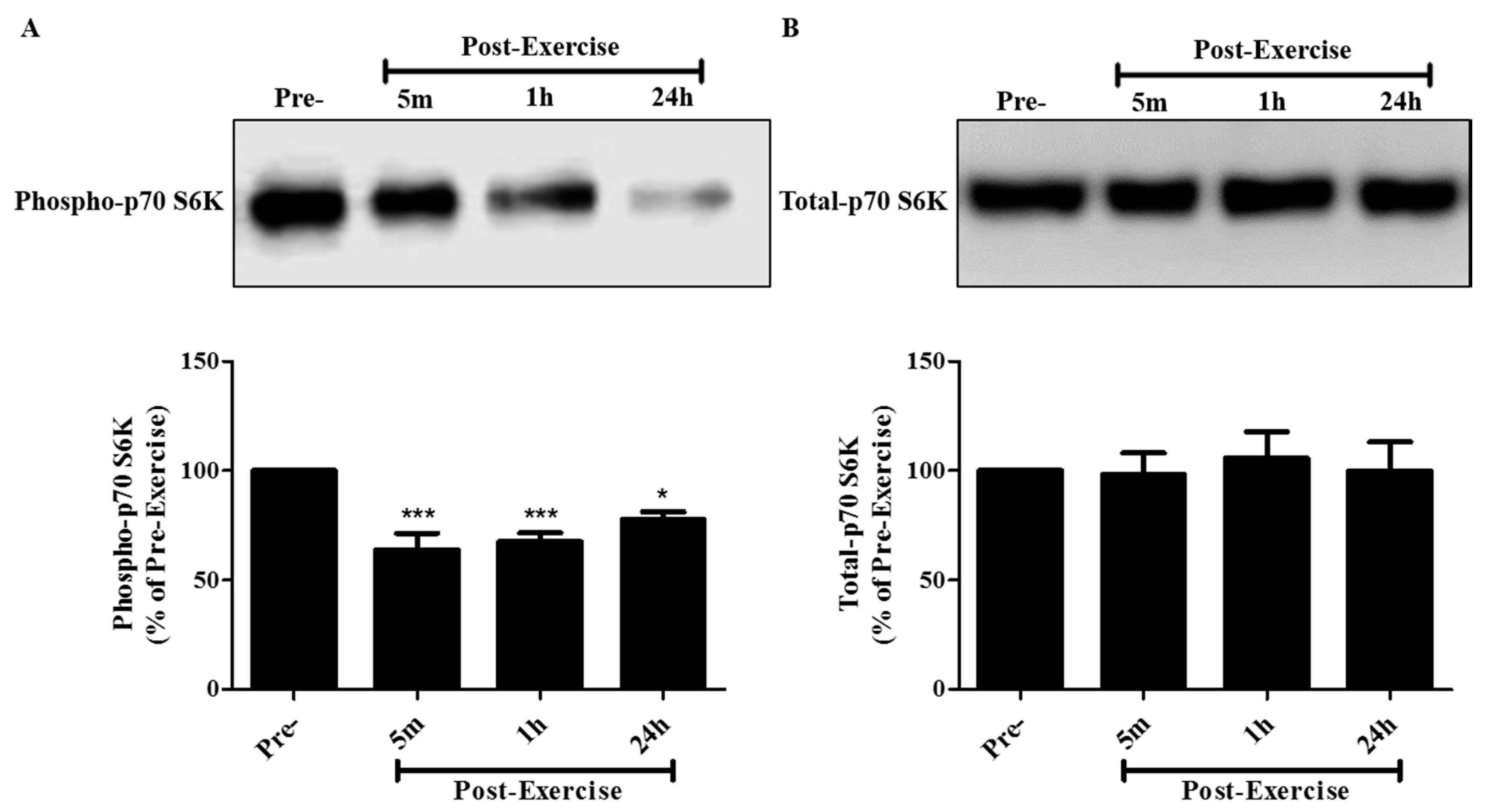
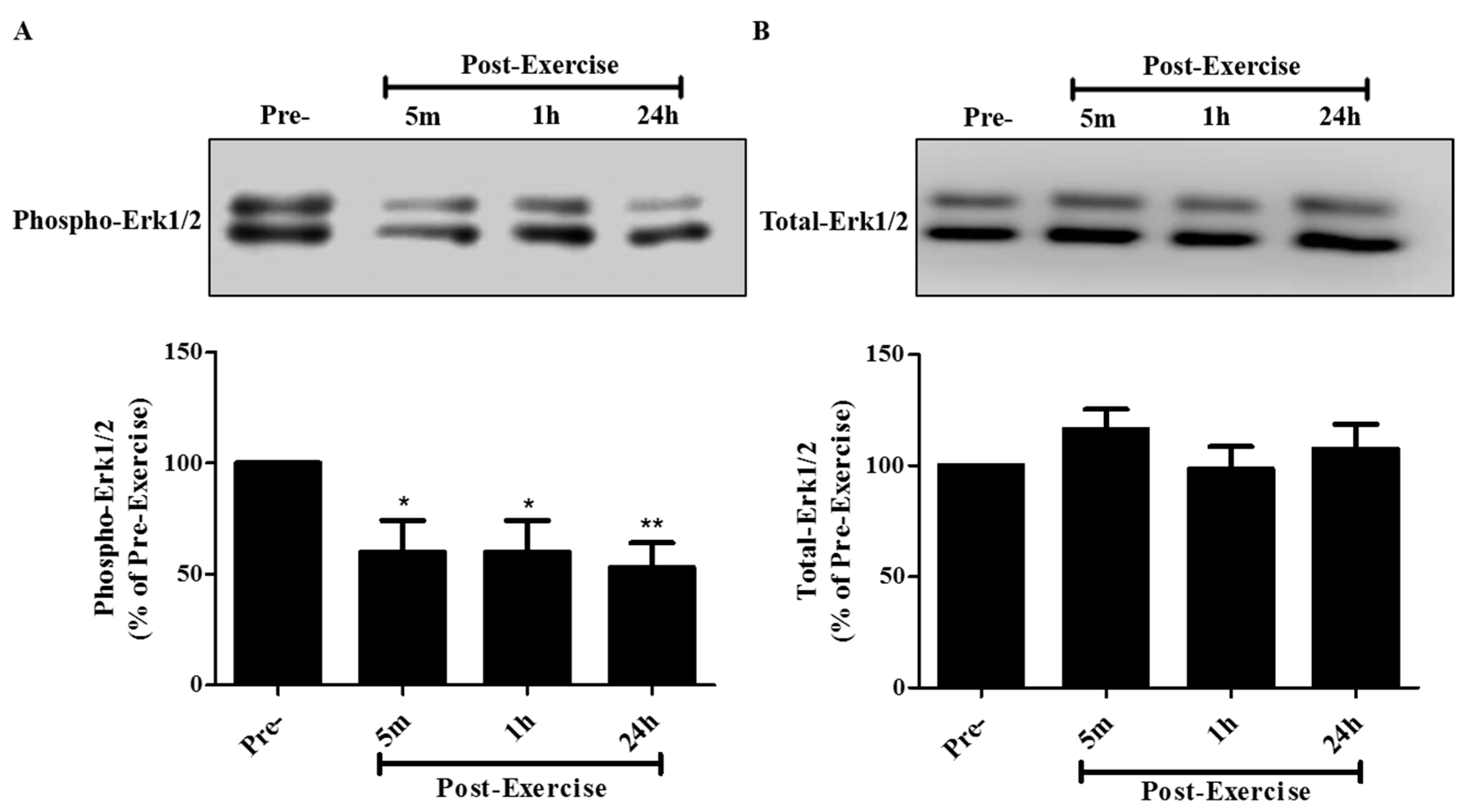
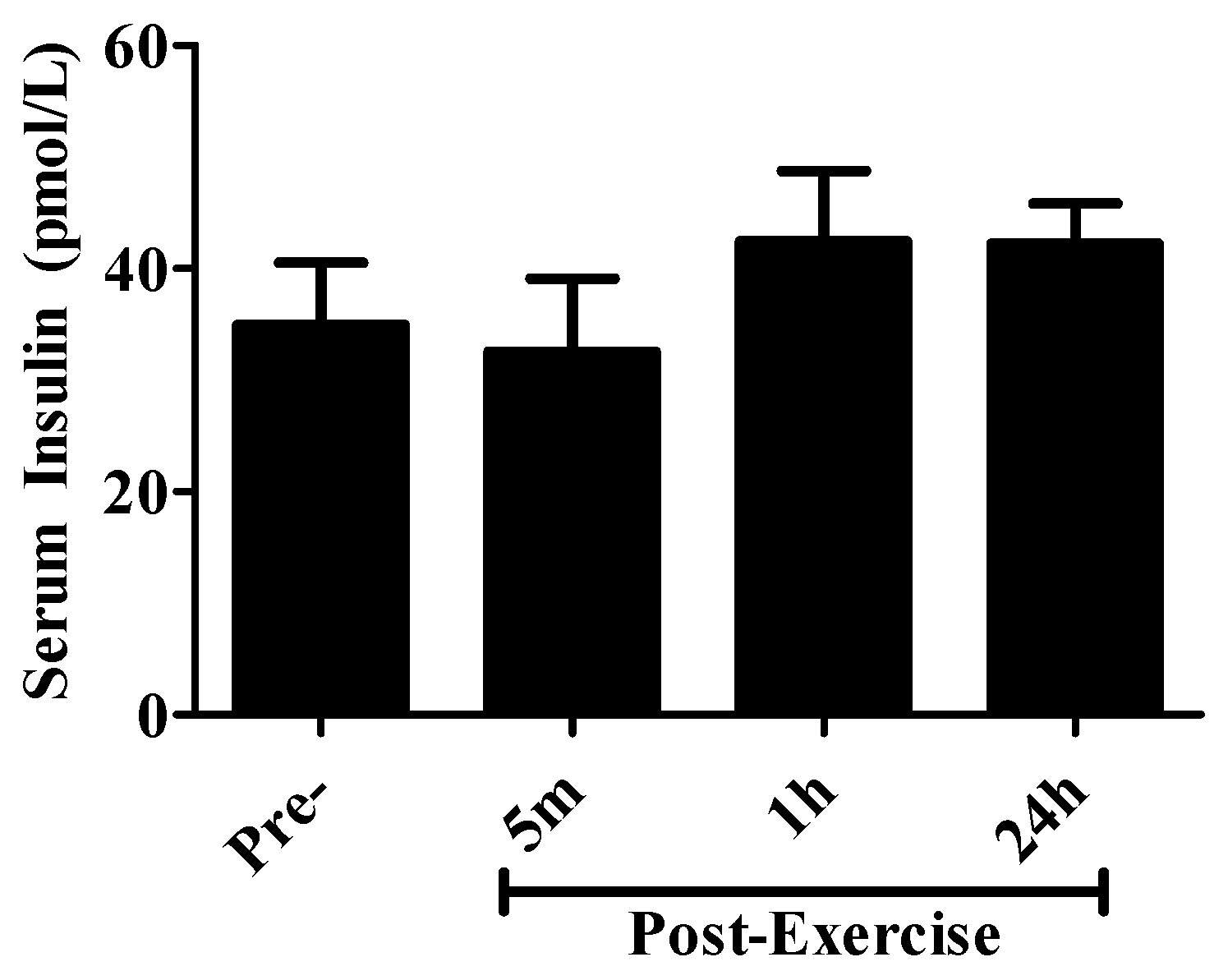
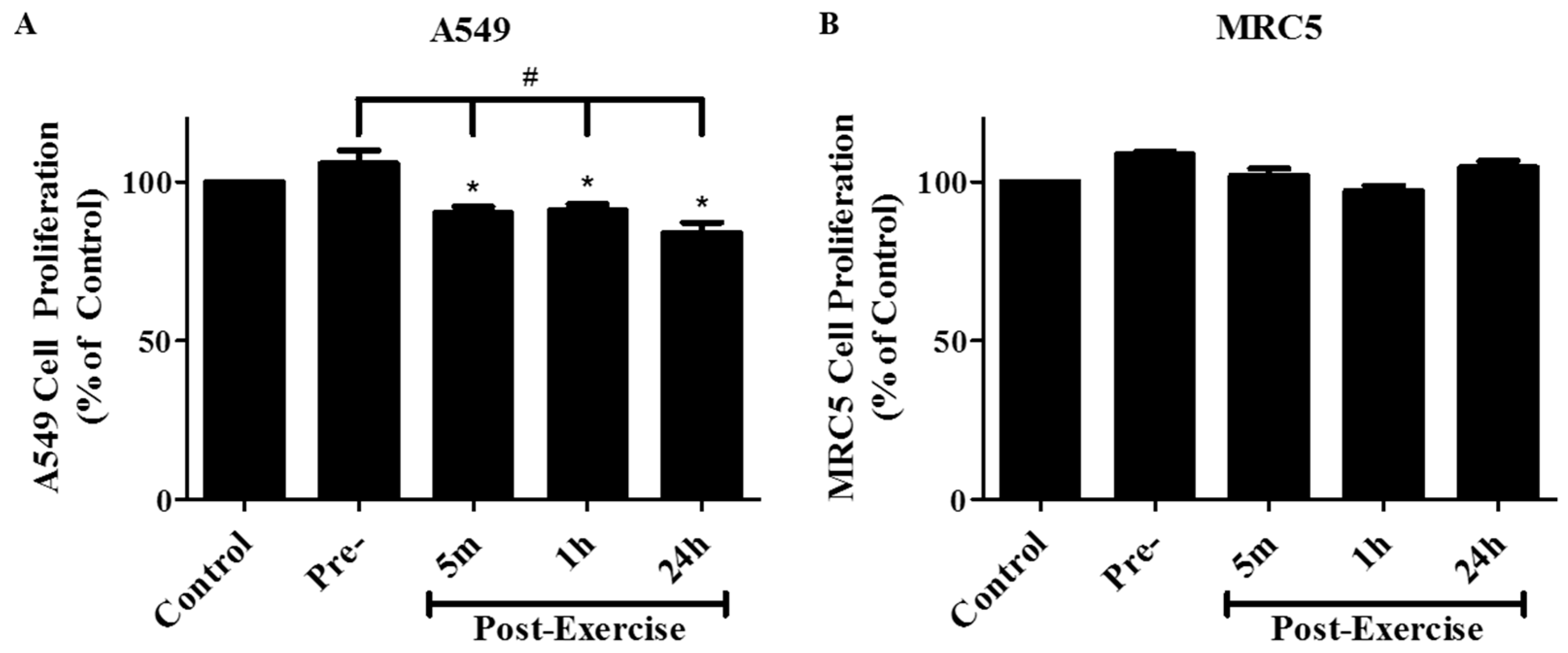
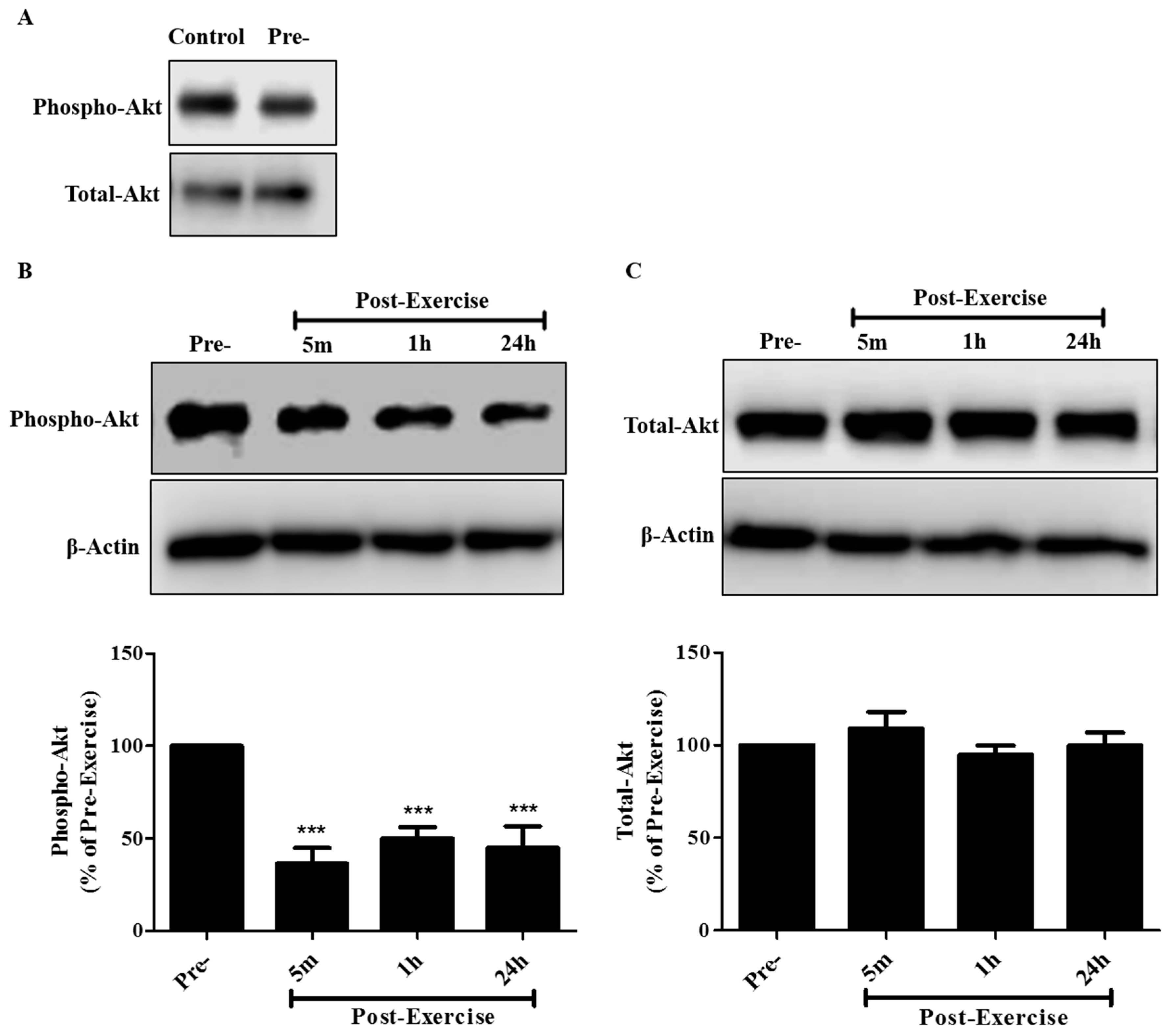
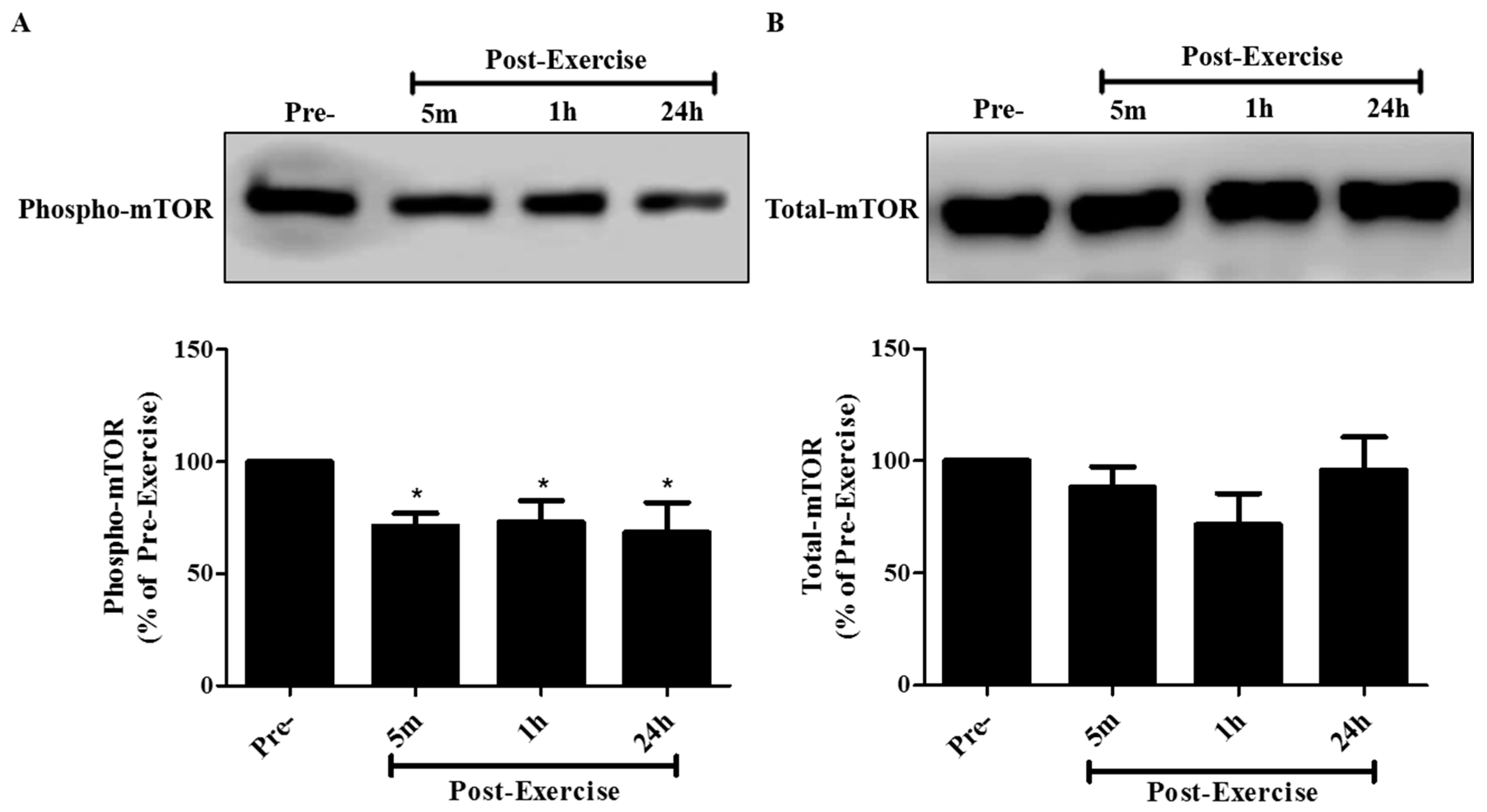
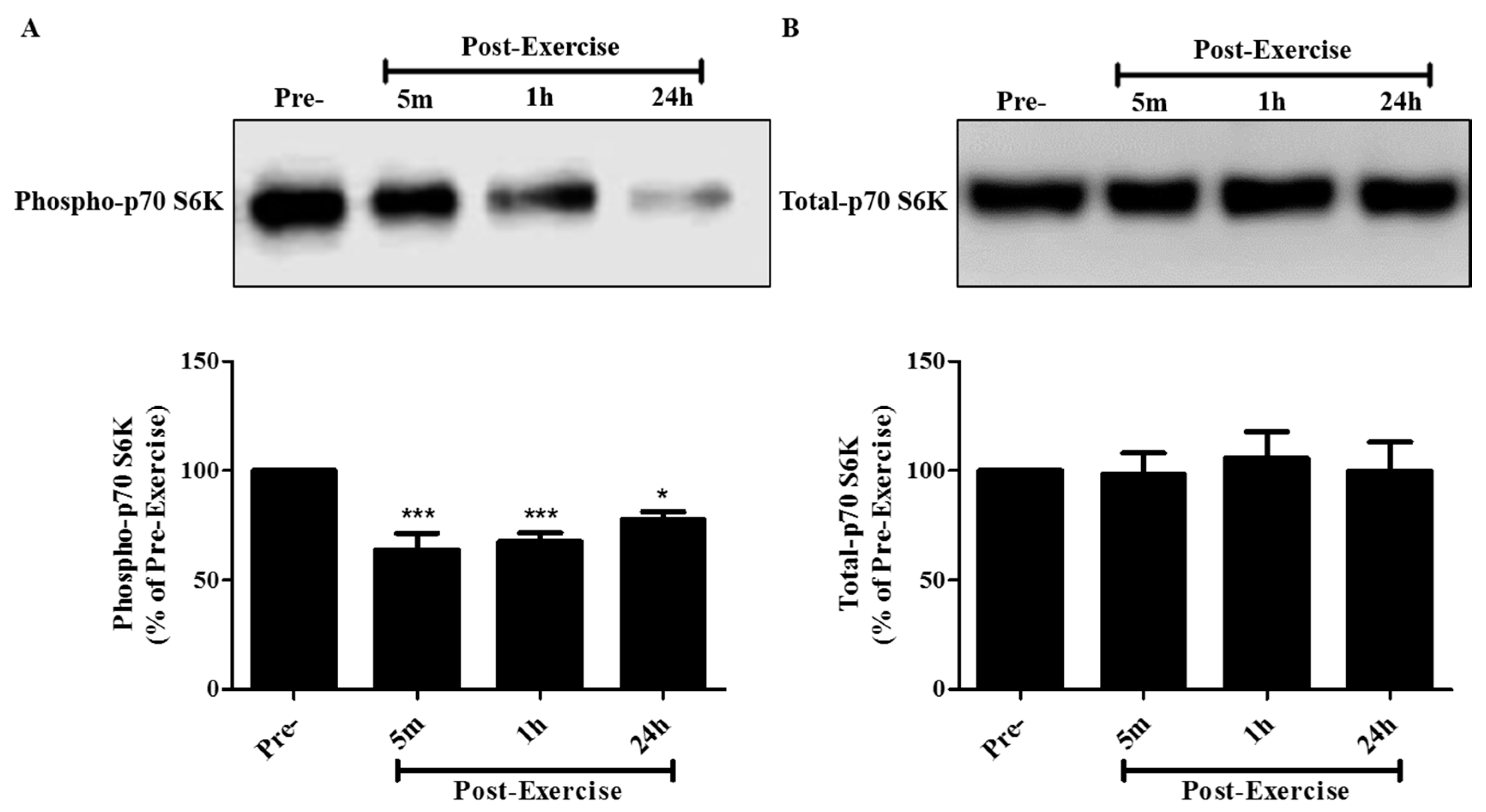
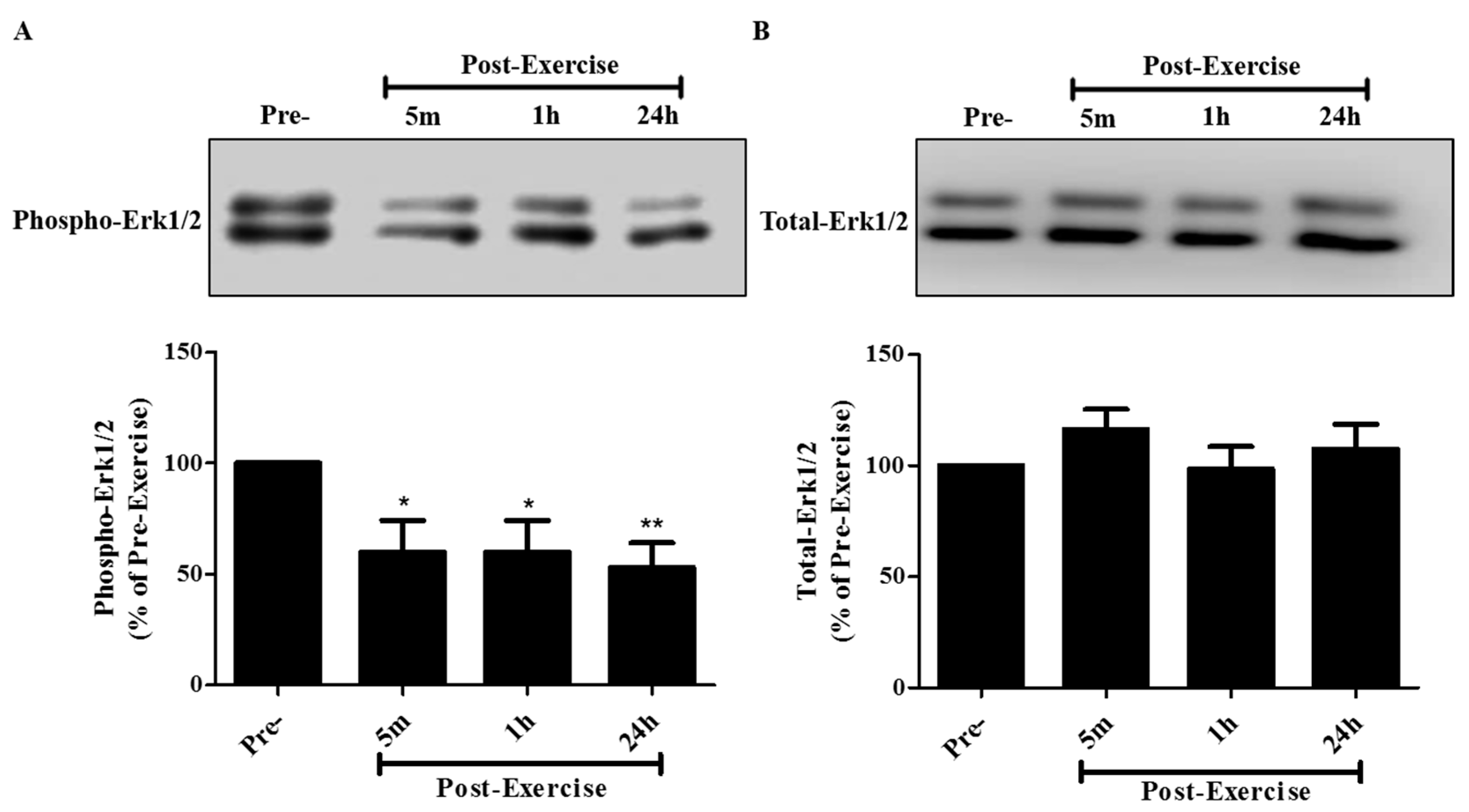
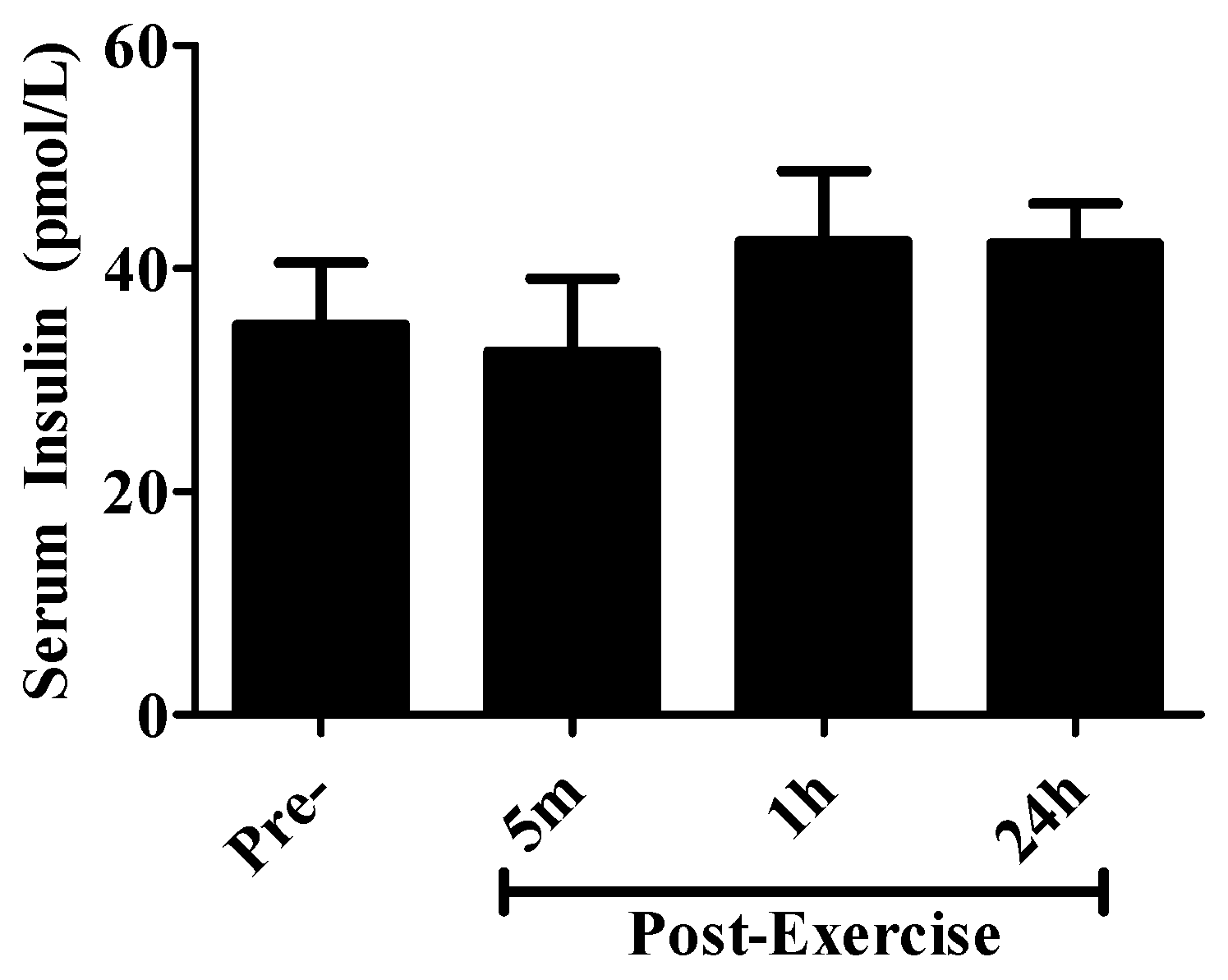
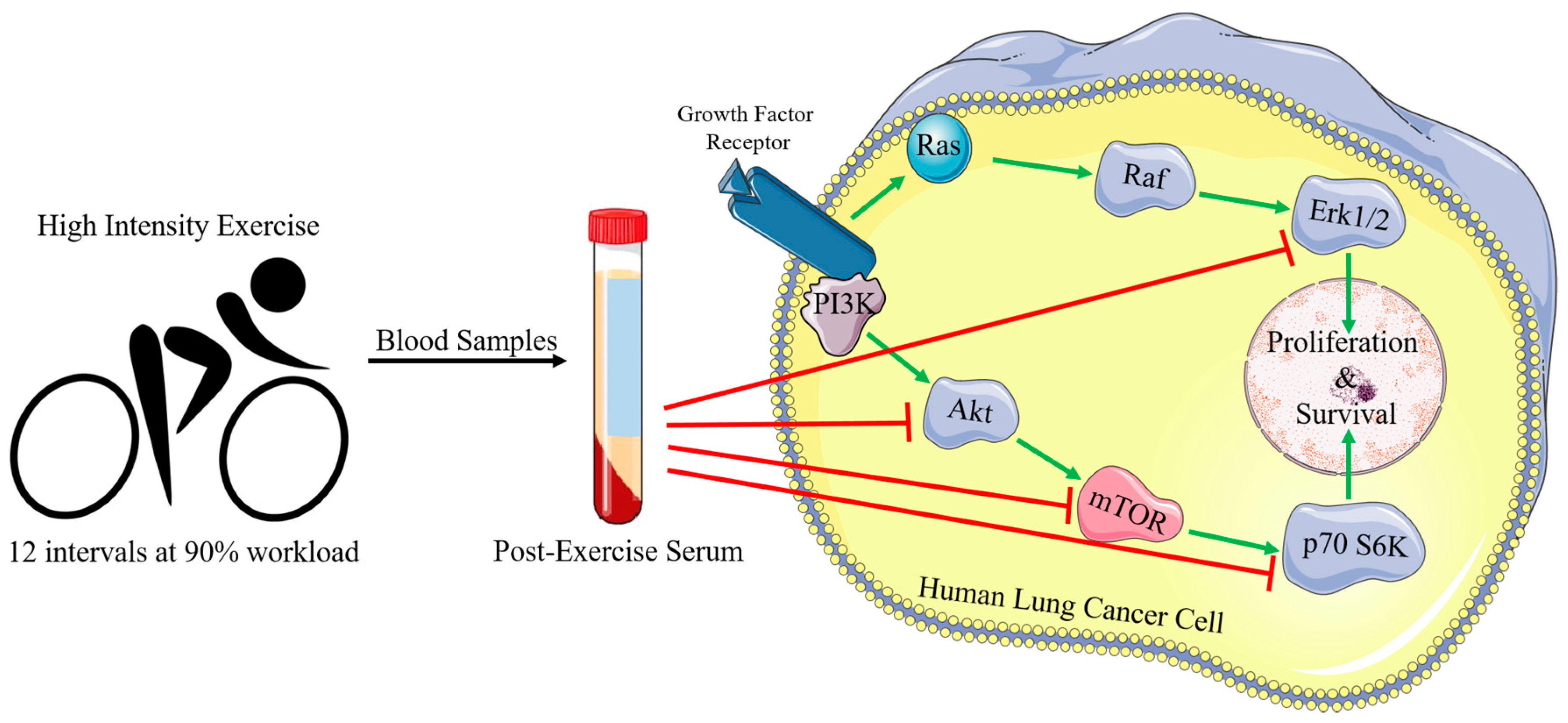
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Exercise Protocol and Blood Collection
4.3. Cell Culture and Treatment
4.4. Cell Proliferation Assay
4.5. Clonogenic Survival Assay
4.6. Immunoblotting
4.7. Serum Insulin Measurements
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non–small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. Mayo Clin. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Devesa, S.S.; Shaw, G.L.; Blot, W.J. Changing patterns of lung cancer incidence by histological type. Cancer Epidemiol. Biomark. Prev. 1991, 1, 29–34. [Google Scholar]
- Devesa, S.S.; Bray, F.; Vizcaino, A.P.; Parkin, D.M. International lung cancer trends by histologic type: Male:Female differences diminishing and adenocarcinoma rates rising. Int. J. Cancer 2005, 117, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Portt, L.; Norman, G.; Clapp, C.; Greenwood, M.; Greenwood, M.T. Anti-apoptosis and cell survival: A review. Biochim. Biophys. Acta 2011, 1813, 238–259. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, J.; Drilon, A. Kras-mutant lung cancers in the era of targeted therapy. Adv. Exp. Med. Biol. 2016, 893, 155–178. [Google Scholar] [PubMed]
- Cheng, H.; Shcherba, M.; Pendurti, G.; Liang, Y.; Piperdi, B.; Perez-Soler, R. Targeting the pi3k/akt/mtor pathway: Potential for lung cancer treatment. Lung Cancer Manag. 2014, 3, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Naizabekov, S.; Chen, Z.; Tokay, T. Power of pten/akt: Molecular switch between tumor suppressors and oncogenes. Oncol. Lett. 2016, 12, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Ilagan, E.; Manning, B.D. Emerging role of mtor in the response to cancer therapeutics. Trends Cancer 2016, 2, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Vicary, G.W.; Roman, J. Targeting the mammalian target of rapamycin in lung cancer. Am. J. Med. Sci. 2016, 352, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ouyang, G.; Bao, S. The activation of akt/pkb signaling pathway and cell survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Balsara, B.R.; Pei, J.; Mitsuuchi, Y.; Page, R.; Klein-Szanto, A.; Wang, H.; Unger, M.; Testa, J.R. Frequent activation of akt in non-small cell lung carcinomas and preneoplastic bronchial lesions. Carcinogenesis 2004, 25, 2053–2059. [Google Scholar] [CrossRef] [PubMed]
- Riely, G.J.; Marks, J.; Pao, W. Kras mutations in non–small cell lung cancer. Proc. Am. Thorac. Soc. 2009, 6, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Asati, V.; Mahapatra, D.K.; Bharti, S.K. K-ras and its inhibitors towards personalized cancer treatment: Pharmacological and structural perspectives. Eur. J. Med. Chem. 2017, 125, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Heigener, D.F.; Gandara, D.R.; Reck, M. Targeting of mek in lung cancer therapeutics. Lancet Respir. Med. 2015, 3, 319–327. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Steelman, L.S.; Kempf, C.R.; Chappell, W.H.; Abrams, S.L.; Stivala, F.; Malaponte, G.; Nicoletti, F.; Libra, M.; Basecke, J.; et al. Therapeutic resistance resulting from mutations in raf/mek/erk and pi3k/pten/akt/mtor signaling pathways. J. Cell. Physiol. 2011, 226, 2762–2781. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.C.; Lee, I.M.; Weiderpass, E.; Campbell, P.T.; Sampson, J.N.; Kitahara, C.M.; Keadle, S.K.; Arem, H.; Berrington de Gonzalez, A.; Hartge, P.; et al. Association of leisure-time physical activity with risk of 26 types of cancer in 1.44 million adults. JAMA Int. Med. 2016, 176, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hu, F.; Li, D.; Wang, F.; Zhu, L.; Chen, W.; Ge, J.; An, R.; Zhao, Y. Does physical activity reduce the risk of prostate cancer? A systematic review and meta-analysis. Eur. Urol. 2011, 60, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Fournier, A.; Dos Santos, G.; Guillas, G.; Bertsch, J.; Duclos, M.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; Mesrine, S. Recent recreational physical activity and breast cancer risk in postmenopausal women in the e3n cohort. Cancer Epidemiol. Biomark. Prev. 2014, 23, 1893–1902. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.D.; Chen, W.Y.; Feskanich, D.; Kroenke, C.H.; Colditz, G.A. Physical activity and survival after breast cancer diagnosis. JAMA 2005, 293, 2479–2486. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.T. Significantly greater reduction in breast cancer mortality from post-diagnosis running than walking. Int. J. Cancer 2014, 135, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Thune, I.; Lund, E. The influence of physical activity on lung-cancer risk: A prospective study of 81,516 men and women. Int. J. Cancer 1997, 70, 57–62. [Google Scholar] [CrossRef]
- Thune, I.; Furberg, A.S. Physical activity and cancer risk: Dose-response and cancer, all sites and site-specific. Med. Sci. Sports Exerc. 2001, 33, S530–S550; discussion S609–S610. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Czerniak, U. Physical activity and its relation to cancer risk: Updating the evidence. Asian Pac. J. Cancer Prev. 2013, 14, 3993–4003. [Google Scholar] [CrossRef] [PubMed]
- Rundqvist, H.; Augsten, M.; Stromberg, A.; Rullman, E.; Mijwel, S.; Kharaziha, P.; Panaretakis, T.; Gustafsson, T.; Ostman, A. Effect of acute exercise on prostate cancer cell growth. PLoS ONE 2013, 8, e67579. [Google Scholar] [CrossRef] [PubMed]
- Dethlefsen, C.; Lillelund, C.; Midtgaard, J.; Andersen, C.; Pedersen, B.K.; Christensen, J.F.; Hojman, P. Exercise regulates breast cancer cell viability: Systemic training adaptations versus acute exercise responses. Breast Cancer Res. Treat. 2016, 159, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Hojman, P. Exercise-induced muscle-derived cytokines inhibit mammary cancer cell growth. Am. J. Physiol. Endocrinol. MeTable 2011, 301, E504–E510. [Google Scholar] [CrossRef] [PubMed]
- Tymchuk, C.N.; Barnard, R.J.; Heber, D.; Aronson, W.J. Evidence of an inhibitory effect of diet and exercise on prostate cancer cell growth. J. Urol. 2001, 166, 1185–1189. [Google Scholar] [CrossRef]
- Ngo, T.H.; Barnard, R.J.; Tymchuk, C.N.; Cohen, P.; Aronson, W.J. Effect of diet and exercise on serum insulin, IGF-I, and IGFBP-1 levels and growth of lncap cells in vitro. Cancer Causes Control 2002, 13, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Tymchuk, C.N.; Barnard, R.J.; Ngo, T.H.; Aronson, W.J. Role of testosterone, estradiol, and insulin in diet- and exercise-induced reductions in serum-stimulated prostate cancer cell growth in vitro. Nutr. Cancer 2002, 42, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Ngo, T.H.; Leung, P.S.; Aronson, W.J.; Golding, L.A. A low-fat diet and/or strenuous exercise alters the igf axis in vivo and reduces prostate tumor cell growth in vitro. Prostate 2003, 56, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.H.; Barnard, R.J.; Leung, P.S.; Cohen, P.; Aronson, W.J. Insulin-like growth factor i (IGF-I) and IGF binding protein-1 modulate prostate cancer cell growth and apoptosis: Possible mediators for the effects of diet and exercise on cancer cell survival. Endocrinology 2003, 144, 2319–2324. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.; Aronson, W.J.; Barnard, R.J. Analyzing serum-stimulated prostate cancer cell lines after low-fat, high-fiber diet and exercise intervention. Evid. Based Complement. Altern. Med. 2011, 2011, 529053. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Leung, P.S.; Aronson, W.J.; Cohen, P.; Golding, L.A. A mechanism to explain how regular exercise might reduce the risk for clinical prostate cancer. Eur. J. Cancer Prev. 2007, 16, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.S.; Aronson, W.J.; Ngo, T.H.; Golding, L.A.; Barnard, R.J. Exercise alters the igf axis in vivo and increases p53 protein in prostate tumor cells in vitro. J. Appl. Physiol. 2004, 96, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Barnard, R.J.; Gonzalez, J.H.; Liva, M.E.; Ngo, T.H. Effects of a low-fat, high-fiber diet and exercise program on breast cancer risk factors in vivo and tumor cell growth and apoptosis in vitro. Nutr. Cancer 2006, 55, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Monda, M.; Messina, G.; Scognamiglio, I.; Lombardi, A.; Martin, G.A.; Sperlongano, P.; Porcelli, M.; Caraglia, M.; Stiuso, P. Short-term diet and moderate exercise in young overweight men modulate cardiocyte and hepatocarcinoma survival by oxidative stress. Oxid. Med. Cell. Longev. 2014, 2014, 131024. [Google Scholar] [CrossRef] [PubMed]
- Whitham, M.; Febbraio, M.A. The ever-expanding myokinome: Discovery challenges and therapeutic implications. Nat. Rev. Drug Discov. 2016, 15, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.M.; Wang, M.L.; Chiou, S.H.; Chen, H.Y.; Wu, C.W. Oncostatin m suppresses metastasis of lung adenocarcinoma by inhibiting slug expression through coordination of stats and piass signalings. Oncotarget 2016, 7, 60395–60406. [Google Scholar] [CrossRef] [PubMed]
- Lauber, S.; Wong, S.; Cutz, J.C.; Tanaka, M.; Barra, N.; Lhotak, S.; Ashkar, A.; Richards, C.D. Novel function of oncostatin m as a potent tumour-promoting agent in lung. Int. J. Cancer 2015, 136, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Naito, Y.; Takagi, T.; Tanimura, Y.; Takanami, Y.; Kawai, Y.; Sakuma, K.; Hang, L.P.; Mizushima, K.; Hirai, Y.; et al. A novel myokine, secreted protein acidic and rich in cysteine (sparc), suppresses colon tumorigenesis via regular exercise. Gut 2013, 62, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Gannon, N.P.; Vaughan, R.A.; Garcia-Smith, R.; Bisoffi, M.; Trujillo, K.A. Effects of the exercise-inducible myokine irisin on malignant and non-malignant breast epithelial cell behavior in vitro. Int. J. Cancer 2015, 136, E197–E202. [Google Scholar] [CrossRef] [PubMed]
- Mezil, Y.A.; Allison, D.; Kish, K.; Ditor, D.; Ward, W.E.; Tsiani, E.; Klentrou, P. Response of bone turnover markers and cytokines to high-intensity low-impact exercise. Med. Sci. Sports Exerc. 2015, 47, 1495–1502. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.Y.; Siopi, A.; Mougios, V.; Park, K.H.; Mantzoros, C.S. Irisin in response to exercise in humans with and without metabolic syndrome. J. Clin. Endocrinol. Metab. 2015, 100, E453–E457. [Google Scholar] [CrossRef] [PubMed]
- Theriau, C.F.; Shpilberg, Y.; Riddell, M.C.; Connor, M.K. Voluntary physical activity abolishes the proliferative tumor growth microenvironment created by adipose tissue in animals fed a high fat diet. J Appl. Physiol. 2016, 121, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Barron, C.C.; Moore, J.; Tsakiridis, T.; Pickering, G.; Tsiani, E. Inhibition of human lung cancer cell proliferation and survival by wine. Cancer Cell Int. 2014, 14, 6. [Google Scholar] [CrossRef] [PubMed]
- Brognard, J.; Clark, A.S.; Ni, Y.; Dennis, P.A. Akt/protein kinase b is constitutively active in non-small cell lung cancer cells and promotes cellular survival and resistance to chemotherapy and radiation. Cancer Res. 2001, 61, 3986–3997. [Google Scholar] [PubMed]
- Lara, P.N., Jr.; Longmate, J.; Mack, P.C.; Kelly, K.; Socinski, M.A.; Salgia, R.; Gitlitz, B.; Li, T.; Koczywas, M.; Reckamp, K.L.; et al. Phase ii study of the akt inhibitor mk-2206 plus erlotinib in patients with advanced non-small cell lung cancer who previously progressed on erlotinib. Clin. Cancer Res. 2015, 21, 4321–4326. [Google Scholar] [CrossRef] [PubMed]
- Gadgeel, S.M.; Wozniak, A. Preclinical rationale for pi3k/akt/mtor pathway inhibitors as therapy for epidermal growth factor receptor inhibitor-resistant non-small-cell lung cancer. Clin. Lung Cancer 2013, 14, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Elrod, H.A.; Lin, Y.D.; Yue, P.; Wang, X.; Lonial, S.; Khuri, F.R.; Sun, S.Y. The alkylphospholipid perifosine induces apoptosis of human lung cancer cells requiring inhibition of akt and activation of the extrinsic apoptotic pathway. Mol. Cancer Ther. 2007, 6, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Massacesi, C.; Di Tomaso, E.; Urban, P.; Germa, C.; Quadt, C.; Trandafir, L.; Aimone, P.; Fretault, N.; Dharan, B.; Tavorath, R.; et al. Pi3k inhibitors as new cancer therapeutics: Implications for clinical trial design. OncoTargets Ther. 2016, 9, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.; Pickering, G.; Gaudette, N.; Tsiani, E. Resveratrol-fortification of red wine does not provide greater inhibition of human lung cancer cell survival compared to non-fortified wine. J. Mol. Biochem. 2015, 4, 52–62. [Google Scholar]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurgan, N.; Tsakiridis, E.; Kouvelioti, R.; Moore, J.; Klentrou, P.; Tsiani, E. Inhibition of Human Lung Cancer Cell Proliferation and Survival by Post-Exercise Serum Is Associated with the Inhibition of Akt, mTOR, p70 S6K, and Erk1/2. Cancers 2017, 9, 46. https://doi.org/10.3390/cancers9050046
Kurgan N, Tsakiridis E, Kouvelioti R, Moore J, Klentrou P, Tsiani E. Inhibition of Human Lung Cancer Cell Proliferation and Survival by Post-Exercise Serum Is Associated with the Inhibition of Akt, mTOR, p70 S6K, and Erk1/2. Cancers. 2017; 9(5):46. https://doi.org/10.3390/cancers9050046
Chicago/Turabian StyleKurgan, Nigel, Evelyn Tsakiridis, Rozalia Kouvelioti, Jessy Moore, Panagiota Klentrou, and Evangelia Tsiani. 2017. "Inhibition of Human Lung Cancer Cell Proliferation and Survival by Post-Exercise Serum Is Associated with the Inhibition of Akt, mTOR, p70 S6K, and Erk1/2" Cancers 9, no. 5: 46. https://doi.org/10.3390/cancers9050046