Highly Regioselective and Stereoselective Biohydroxylations of Oxandrolone


Department of Chemistry, Wrocław University of Environmental and Life Sciences, 50-375 Wrocław, Poland
*
Authors to whom correspondence should be addressed.
Catalysts 2021, 11(1), 16; https://doi.org/10.3390/catal11010016
Submission received: 5 December 2020
/
Revised: 20 December 2020
/
Accepted: 23 December 2020
/
Published: 25 December 2020
(This article belongs to the Special Issue Biocatalysis-Biotransformations in Organic Synthesis)
Abstract
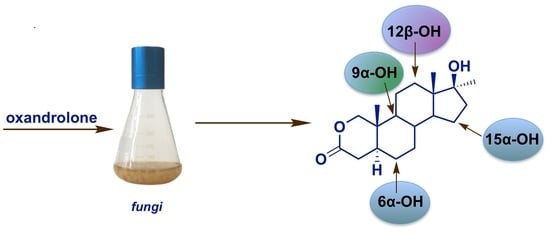
:Microbially catalyzed reactions are a powerful and valuable tool for organic synthesis of many compounds with potential biological activity. Herein, we report efficient hydroxylations of the steroidal anabolic-androgenic lactone, oxandrolone, in the cultures of three strains of fungi, Fusarium culmorum, Mortierella isabellina, and Laetiporus sulphureus. These reactions resulted in the production of four metabolites identified as 12β-hydroxyoxandrolone (2), 9α-hydroxyoxandrolone (3), 6α-hydroxyoxandrolone (4), and 15α-hydroxyoxandrolone (5), the latter being a new compound. The high substrate conversion rates and the product yields achieved indicate that these strains offer a new way to generate steroidal hydroxylactones with potential pharmaceutical interest. The structures of the isolated derivatives were characterized on the basis of spectroscopic data. The effect of modification of the A-ring structure of the steroid by the lactone group on the selectivity of hydroxylation in cultures of the tested fungi is also discussed.
1. Introduction
The ability of microorganisms to transform organic compounds makes them a suitable instrument for obtaining valuable molecules that cannot be formed easily by traditional chemical methods. As they occur at neutral pH and ambient temperature, and do not require expensive and harmful chemicals, biocatalytic reactions are easy to carry out, cost-effective and safe for the environment. Most fungal strains contain cytochrome P450 monooxygenases, similar to those of mammals, so they can carry out regioselective and stereoselective hydroxylation of molecules, such as steroids, which may increase or modify their biological activity [1]. The presence of a hydroxyl group in organic compounds affects their polarity, toxicity, extraction from the cell, and translocation via the cell membranes, while creating possibilities for various subsequent structural modifications in the search for new substances with healing properties [1,2].
Microbial hydroxylations are used commercially for the production of steroidal drugs and to produce steroidal key intermediates for their synthesis. The 11α-, 11β-, 9α-, and 16α-hydroxylations are utilized in the global manufacture of corticosteroids [2,3]. The introduction of a hydroxyl group at the 11α-position of progesterone by Rhizopus nigricans decreased the number of chemical steps from 36 to 11 in cortisone synthesis from deoxycholic acid and caused a remarkable reduction in the price of the drug [4]. As another example, 11β-hydroxylation of cortexolone by Culvularia lunata is used to obtain hydrocortisone [2,4,5], and 9α-hydroxy steroids are intermediates for the production of more active 9α-halogenated corticoids. The presence of a 16α-hydroxyl group in halogen corticosteroids enables a reduction of the undesirable mineralocorticoid activity of these derivatives [2,3,6]. Steroid hydroxylations occurring in other sites, including the 7α-, 7β-, 14α- and 15α-positions, may have the potential for industrial operation. The 7α-hydroxylation of cholesterol is essential for the production of bile acids, which have a variety of applications in the biomedical and pharmaceutical fields due to their peculiar physicochemical and biological properties [1,7]. Furthermore, 7-hydroxy derivatives of steroidal 5-en olefins, e.g., the widely available dietary supplement dehydroepiandrosterone (DHEA), exhibit several-fold higher immunoprotective and immunoregulatory properties than their precursors [8,9,10]. Other studies report that 14α-hydroxylation is used in the synthesis of intermediates for the production of cardioactive steroids or inhibitors for breast cancer cells [1,2], and 15α-hydroxylation is employed in the synthesis of contraceptives [2]. Although during systematic investigations it has been proved that, depending on the specificity of the given microorganism, every site in the steroid molecule is accessible for hydroxylation, current trends focus on the detection and isolation of strains with novel activity or a more efficient transformation capacity. Due to the nature of fungal hydroxylases and difficulties related to their isolation, purification and maintenance of catalytic activity, whole-cell transformation processes remain the method of choice for these studies.
In this study, as a continuation of the research on the transformation of androgen-anabolic steroids, reactions of a steroidal lactone-oxandrolone (1) with three fungal species (Fusarium culmorum AM282, Mortierella isabellina AM212 (current name Umbelopsis isabellina [11]), and Laetiporus sulphureus AM498) were carried out. Fusarium culmorum and M. isabellina were previously known for their ability to metabolize steroidal compounds with different structures [9,12,13,14,15].
Oxandrolone (17β-hydroxy-17α-methyl-2-oxa-5α-androstan-3-one) is a widely available synthetic 2-oxa derivative of dihydrotestosterone with much lower androgenic activity than the parent compound. First synthesized in 1962, two years later it was introduced to the US market under the name Anavar® and was used to treat alcohol hepatitis, Turner syndrome and osteoporosis [16,17,18]. Nowadays, oxandrolone is also used in the treatment of genetic growth disorders, people with significant physical trauma, particularly burns, and patients with AIDS who have suffered a large weight loss [19]. The use of oxandrolone significantly decreases wound healing time and increases weight re-gain, which directly translates into reducing the length of hospital stay [20]. As this compound does not significantly affect testosterone production and does not aromatize, it is considered the safest of all steroids [21]. Regarding oxandrolone’s accessibility, anabolic activity and relatively weak side-effects, it is also one of the most often abused drugs in sport [22,23]. It should be noted that, according to the regulations of the World Anti-Doping Agency, the administration of oxandrolone is prohibited [24]. Among the known metabolites of oxandrolone identified in humans are its epimeric forms—epioxandrolone, 18-nor-oxandrolone, and 16α-, and 16β-hydroxyoxandrolone [25]. Additionally, 17α- and 17β-hydroxymethyl-18-nor-oxandrolone have been detected as long-term metabolites [22].
Unlike other widely tested anabolic 17α-alkylated androstanes, i.e., 17α-methyltestosterone [26,27], so far only a few studies concerning oxandrolone’s bioconversion have been carried out. That substrate underwent 11α-, 6α- and 9α-hydroxlation with Rhizopus stolonifer [21], and 12β-hydroxylation with Cunninghamella blakesleeana [28]; however, the final yield of synthesized products was quite low (0.8–24%). Only 11α-hydroxyoxandrolone was synthesized with a 24% yield [21]. The mixture of four other oxygenated metabolites, namely, 11β-hydroxyoxandrolone (2.8%); 5α,11β-dihydroxyoxandrolone (0.9%), 11β,20-dihydroxyoxandrolone (2.3%), and 17β-hydroxy-17α-methyl-2-oxa-5α-androstan-3,11-dione (1.4%), was obtained after incubation of that steroid in Macrophomina phaseolina culture [28].
Here, we present the results of an effective transformation of oxandrolone (1) to the known derivatives 12β- (2), 9α- (3), and 6α-hydroxyoxandrolone (4), and to the new compound–15α-hydroxyoxandrolone (5). The effect of the modification of the A-ring structure of the steroid by the lactone group on the selectivity of hydroxylation in cultures of the tested fungi is also discussed.
2. Results and Discussion
Our results indicated that F. culmorum AM282, M. isabellina AM212 and L. sulphureus AM498 were able to convert oxandrolone (1) into its polar metabolites. The time-dependent progress of the transformations is compiled in Table 1. The structures of the obtained derivatives were established by spectroscopic methods by comparison of the characteristic shift values of selected diagnostic signals of products and the starting compound. The stereochemistry of the hydroxyl group was deduced on the basis of NOESY experiments.
The bioconversion of oxandrolone (1) with F. culmorum yielded only one main metabolite (Figure 1), which was obtained with a yield of 62% mol. (69% determined by GC analysis, see Table 1).
Mass spectrometry (MS) data of metabolite 2 revealed an [M]+ at m/z 323.2 (C19H30O4), reflecting the additional OH group in the molecule. The 1H-NMR spectrum showed a new downfield methine proton signal at δH 3.73–3.76 ppm, geminal to the hydroxyl group. In comparison to the spectrum of the substrate, the C-18 methyl signal of 2 demonstrated a downfield shift (Δ 0.07 ppm), suggesting that hydroxylation had occurred at the 12β position. The appearance of a new methine carbon signal at δC 73.1 ppm in the 13C NMR spectrum, in combination with the downfield shifts of C-11 (Δ 10.9 ppm) and C-13 (Δ 4.1 ppm), and upfield shift of C-18 (Δ 5.5 ppm) when compared to the spectrum of 1, was an important confirmation of 12β-hydroxylation. Additionally, the NOESY spectrum showed correlations between 12α-H and 9α- (δH 0.90 ppm), 14α- (δH 1.18 ppm) proton, and C-17α (δH 1.35 ppm) methyl group signals. Thus, product 2 was identified as 12β-hydroxy-oxandrolone. Its NMR data were comparable with those reported in the literature [28]. The effective introduction of 12β-OH group into the structure of 17α-alkylated androstanes by F. culmorum was not surprising. Although this strain converts C19 3-oxo-4-ene and 3β-hydroxy-5α-saturated steroids to a mixture of their 6β- and 15α-hydroxy derivatives [13,14,15,29], in the presence of an alkyl substituent at the 17α-position, which is a drawback for 1–3 cis hydroxylation at the 15α-position, the reaction occurs at position 12β, which is its equivalent in enzyme-substrate complexes [2]. Thus, after transformation of 17α-methyltestosterone in F. culmorum AM282 culture, the mixture of 6β-, 15α-, and 12β-hydroxy derivatives at a 5:2:2 ratio was obtained [14]. We also found that the modification of the ring A of steroids and replacement of 3-oxo-4-ene group with an unusual δ lactone moiety makes such orientation of the substrate in the complex with the enzyme, that 12β-hydroxylation occurs. [2]. 12β-hydroxy-oxandrolone (2) was previously synthesized after 18 days of the transformation of 1 using Cunninghamella blakesleeana strain but it was isolated with only a 0.8% yield [28].
After seven days of incubation of oxandrolone (1) with M. isabellina AM212, three potential products were detected in the post-transformation extracts, but only one metabolite, 3 (80% based on GC analysis, see Table 1), was present in sufficient quantity for characterization (Figure 2).
The MS data of metabolite 3 exhibited an [M]+ at m/z 323.2, which implied the incorporation of an oxygen atom to the substrate. In the 1H NMR spectrum of 3, the CHOH signal was absent. The signal of OH-bearing quaternary carbon at δC 74.2 ppm, the downfield shift of signals C-10 (Δ 4.4 ppm), C-11 (Δ 2.1 ppm), and C-8 (Δ 2.7 ppm), and the γ-gauche upfield shifts of C-1 (Δ 4.4 ppm), C-5 (Δ 8.1 ppm), C-7 (Δ 3.5 ppm), C-12 (Δ 4.1 ppm), and C-14 (Δ 6.8 ppm) signals in comparison with the spectrum of metabolite 1 indicated that 9α-hydroxylation had occurred. The proposed structure, 9α-hydroxy-oxandrolone (3), was supported by the downfield shift of C-19 methyl signal by 0.10 ppm with respect to the substrate. The spectroscopic data of this metabolite were consistent with those described in the literature [21] and 9α-hydroxyoxandrolone (3) was previously reported as one of three products of a transformation using Rhizopus stolonifer with an isolated yield of 8% after 10 days of cultivation [21]. In the current study with M. isabellina AM212, the yield was improved to 71%. In comparison to the transformation of epiandrosterone (3β-hydroxy-5α-androstan-17-one) by M. isabellina, which resulted in a mixture of 7α-, 9α-, and 11α-hydroxy derivatives [9], there is another example that the introduction of a lactone moiety into the A-ring of 5α-saturated substrate enhances the regioselectivity of the hydroxylation.
Two others metabolites (4 and 5 at a 1:4.2 ratio) were detected after ten days of incubation of 1 with Laetiporus sulphureus AM498 (Table 1, Figure 3).
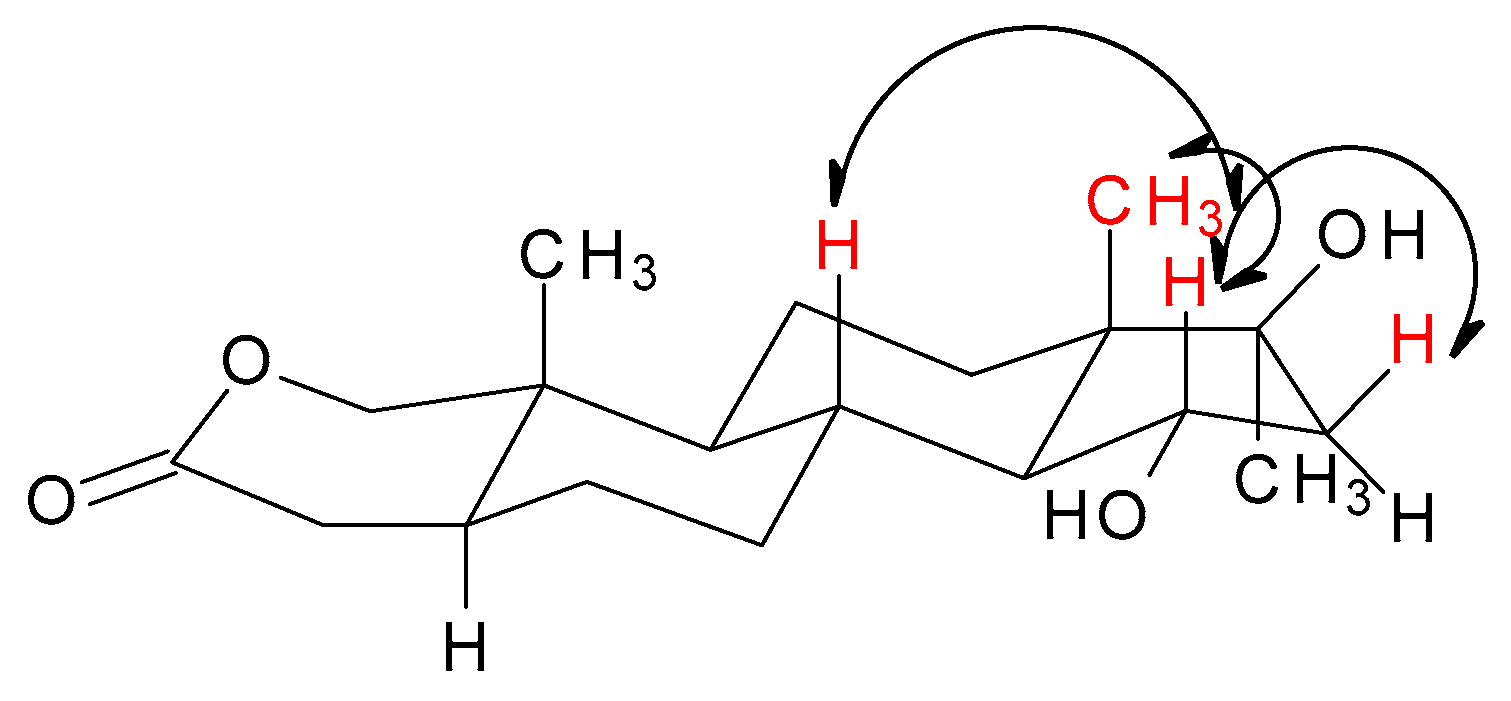
The MS spectra of both metabolites showed an [M]+ at m/z 323.2, corresponding to the formula C19H30O4 and indicating the incorporation of an oxygen atom into the structure of the substrate. The 1H NMR spectrum of the less polar product revealed a new broad signal at δH 3.45–3.49 ppm suggesting hydroxylation at an equatorial proton. The significant shift of the 4-H signals (by 0.46 ppm and 0.12 ppm) and lack of a shift of both the methyl signals pointed towards substitution at 6α-H. The appearance of a new methine carbon signal at δC 69.8 ppm, in combination with the downfield shift of the C-5 (Δ 6.7 ppm) and C-7 (Δ 10.0 ppm), and γ-carbon upfield signal shift of C-4 (Δ 3.1 ppm) and C-8 (Δ 0.9 ppm) confirmed 6α-hydroxylation. Additionally, the NOESY spectrum showed a correlation between 6β-H and 4β-(δH 2.34 ppm), 8β (δH 1.61 ppm) proton, and C-19 (δH 1.02 ppm) methyl signals. These observations fully supported the structure of 4 as 6α-hydroxyoxandrolone and were consistent with the spectroscopic data available in the literature [21]. It was previously reported that 6α-hydroxyoxandrolone (4) was one of three metabolites obtained after 10 days transformation of 1 by Rhizopus stolonifer with a yield of 5% [21]. The 1H NMR spectrum of the main reaction product, metabolite 5, showed a new broad hydroxyl-bearing methine proton signal at δH 4.07 ppm. The characteristic shape and multiplicity of this signal (td, J = 3.4 Hz; J = 9.3 Hz) suggested 15α-hydroxylation. This was supported by an oxygen-bearing methine carbon signal at δC 72.3 ppm, downfield shifts of the β-carbons C-14 (Δ 7.7 ppm) and C-16 (Δ 11.6 ppm), and γ-carbon upfield signal shifts of C-8 (Δ 0.5 ppm) and C-17 (Δ 2.5 ppm). Further supporting evidence of the stereochemistry of the new hydroxyl group was provided by NOESY spectrum analysis, in which the correlation signals between 15β-H and H-8 (δH 1.68 ppm), and H-16 (δH 2.42 ppm) and C-18 methyl (δH 0.87 ppm) signals were visible (Table 2 and Figure 4). These results confirmed the identification of 5 as 15α-hydroxyoxandrolone. This compound is a new derivative of oxandrolone.
To the best of our knowledge, this is the first report confirming 15α-hydroxylation of oxandrolone by fungi. The use of L. sulphureus AM498 allowed us to obtain this hydroxylactone with a high yield (70% mol.). Moreover, the use of mycelium of this edible polypore in the transformation of steroids compounds is described here for the first time.
3. Materials and Methods
3.1. Substrate and Microorganisms
The substrate oxandrolone (1) was purchased from Biosynth Carbosynth® (Berkshire, United Kingdom). The fungal strains Fusarium culmorum AM282, Mortierella isabellina AM212 and Laetiporus sulphureus AM498 were obtained from the collection of the Department of Pharmaceutical Biology and Botany of the Wrocław Medical University, Wrocław, Poland. Fungi were maintained on Sabouraud 4% dextrose agar slopes and freshly sub-cultured before use in the transformation experiments.
3.2. Conditions of Cultivation and Transformation
All strains were incubated in 300 mL Erlenmeyer flasks with 100 mL of a medium composed of 3% of glucose and 1% of proteose peptone (Biocorp, Warsaw Poland). Fungi were grown at 26 °C on a rotary shaker (130 rpm). After a specified time of cultivation that depended on the growth rate of the strain (2 days for Laetiporus sulphureus AM498 and Fusarium culmorum AM282, 3 days for Mortierella isabellina AM212), 0.06 g of substrate 1 in the form of an acetone suspension was equally distributed among three flasks with fungal cultures (final concentration 0.2 gl−1). Transformations were carried out under the same conditions until the substrate was metabolized. To study the time-dependent progress of the bioconversion, 5 mL samples of the transformation medium were taken out at regular intervals (48 h) from the reaction flasks and processed as described in Section 3.3.
3.3. Isolation and Identification of Products
After transformations were performed, the mixture was extracted three times with chloroform (3 × 100 mL). The organic extract was dried over anhydrous magnesium sulphate, concentrated in vacuo and analyzed by TLC and GC. Merck Silica gel 60 F254 plates and eluent chloroform/methanol (70:6 v/v) were used for TLC analysis. To detect the steroids, the TLC plates were sprayed with a methanol/concentrated sulfuric acid (1:1 v/v) mixture, heated at 120 °C until the colors developed, and observed under UV light (254 nm). GC analysis was performed on a Hewlett Packard 5890 Series II GC instrument (FID, carrier gas H2) with the DB-5MS capillary column (30 m × 0.32 mm, film 0.25 µm; Agilent Technologies, Inc., Warsaw, Poland). The temperature program for the GC analysis was as follows: 220 °C for 1 min increasing by increments of 4 °C/min to 270 °C, then at a gradient of 30 °C/min to 300 °C, held at 300 °C for 3 min; the injector and detector temperature was taken at 300 °C. MS analysis was performed on Varian CP-3800/Saturn 2000 apparatus (Varian, Walnut Creek, CA, USA) with a Zebron ZB-5 MSI (30 m × 0.25 mm × 0.25 μm) column (Phenomenex, Torrance, CA, USA). Temperature conditions were as follows: 220 °C for 1 min, increasing by 5 °C/min to 300 °C, held at 300 °C for 7 min. The injector temperature was 270 °C and the detector temperature was 220 °C. Biotransformation products were separated by column chromatography using Merck Silica gel 60 (0.040–0.063 mm) with chloroform/methanol (70:6 v/v) as eluent. After evaporation of the eluent, the separated metabolites were weighed. The products’ structures were defined in accordance with 1H NMR and 13C NMR analysis. The spectra were recorded on a Bruker AvanceTM 600 MHz spectrometer in CDCl3. Jasco P-2000 digital polarimeter (the version with an iRM controller, Mary’s Court, Easton, MD, USA) was used to measure optical rotation. Melting points (uncorrected) were determined on the Boetius apparatus.
3.4. Transformations and Spectroscopic Data of the Isolated Metabolites
3.4.1. Transformation with Fusarium culmorum
The seven day transformation process of oxandrolone (1) with F. culmorum AM282 resulted in a mycelial and broth extract, which was chromatographed on silica gel. Elution with methanol in chloroform yielded untransformed oxandrolone (1) (11 mg, 18%mol.), and 12β-hydroxyoxandrolone (2) (39 mg, 62% mol.). Compound 2 was as follows: white plates, mp 190–193 °C, = −49 (c 0.02, CHCl3) (lit.: mp 195–196 °C), = −55 (c 0.01, MeOH) [28]. EI-MS m/z 323.2 [M]+ (6), 307.3 (5), 286.2 (60), 246.5 (100), 43.1 (57). 1H NMR (CDCl3, 600 MHz) δ (ppm): 0.90 (m, 1H, 9-H), 0.93 (s, 3H, 18-H), 1.02 (s, 3H, 19-H), 1.18 (m, 1H, 14-H), 1.21 (m,1H, 6α-H), 1.32 (m, 1H, 11α-H), 1.35 (s, 3H, 20-H), 1.41 (m, 1H, 15β-H), 1.43 (m, 1H, 8-H), 1.44 (m, 1H, 11β-H), 1.48 (m, 1H, 7β-H), 1.49 (m, 1H, 6βH), 1.63 (m, 1H, 15α-H), 1.68 (m, 1H, 16β-H), 1.72 (m, 1H, 7α-H), 1.76 (m, 1H, 5-H), 1.85 (m, 1H, 16α-H), 2.23 (dd, J = 13.1 Hz, J = 18.7 Hz, 1H, 4β-H), 2.53 (dd, J = 5.8 Hz, J = 18.6 Hz, 1H, 4α-H), 3.73–3.76 (m, 1H, 12α-H), 3.92 (d, J = 10.7 Hz, 1H, 1α-H), 4.21 (d, J = 10.7 Hz, 1H, 1β-H); 13C NMR (CDCl3, 151 MHz) δ (ppm): 8.4 (C-18), 10.1 (C-19), 23.0 (C-15), 25.6 (C-20), 27.0 (C-6), 29.8 (C-11), 30.2 (C-7), 33.6 (C-4), 34.6 (C-10), 34.7 (C-8), 38.1 (C-16), 40.3 (C-5), 48.2 (C-9), 48.9 (C-14), 49.4 (C-13), 73.1 (C-12), 80.7 (C-1), 82.0 (C-17), 170.3 (C-3). See Figures S1–S4 in the Supplementary Materials for the MS and NMR spectra of compound 2.
3.4.2. Transformation with Mortierella isabellina (Umbelopsis isabellina)
The seven day transformation of oxandrolone (1) with M. isabellina AM212 resulted in a mycelial and broth extract, which was chromatographed on silica gel. Elution with methanol in chloroform yielded 9α-hydroxyoxandrolone (3) (45 mg, 71% mol.). Compound 3: amorphous solid, = −52 (c 0.02, CHCl3) (lit.: = −55 (c 0.0008, CHCl3)) [21]. EI-MS m/z 323.2 [M]+ (36), 307.2 (13), 286.2 (48), 246.5 (80), 124.3 (61). 1H NMR (CDCl3, 600 MHz) δ (ppm): 0.89 (s, 3H, 18-H), 1.09 (s, 3H, 19-H), 1.21 (m, 1H, 6β-H), 1.25 (s, 3H, 20-H), 1.30 (m, 1H, 11β-H), 1.34 (m, 1H, 15β-H), 1.41 (m,1H,7β-H), 1.48 (m, 1H, 15α-H), 1.53 (m, 1H, 6α-H), 1.55 (m, 1H, 11α-H), 1.58 (m, 1H, 14-H), 1.71 (m, 2H, 12β-H, 16β-H, 1.75 (m, 2H, 8-H, 12α-H), 1.80 (m, 1H, 16α-H), 1.88 (m, 1H, 7α-H), 2.18 (dd, J = 12.8 Hz, J = 18.6 Hz, 1H, 4β-H), 2.45 (m, 1H, 5-H), 2.56 (dd, J = 6.2 Hz, J = 18.6 Hz, 1H, 4α-H), 4.14 (d, J = 10.6 Hz, 1H, 1α-H), 4.53 (d, J = 10.6 Hz, 1H, 1β-H); 13C NMR (CDCl3,151 MHz) δ (ppm): 11.9 (C-19), 13.0 (C-18), 23.0 (C-11), 24.2 (C-15), 26.0 (C-20), 27.0 (C-6), 27.1(C-12), 27.3 (C-7), 32.4 (C-5), 34.0 (C-4), 38.3 (C-8), 38.7 (C-16), 39.1 (C-10), 43.4 (C-14), 45.4 (C-13), 74.2 (C-9), 76.6 (C-1), 81.5 (C-17), 170.4 (C-3). See Figures S5–S7 for the MS and NMR spectra of compound 3.
3.4.3. Transformation with Laetiporus sulphureus
The ten day transformation process of oxandrolone (1) with L. sulphureus AM498 resulted in a mycelial and broth extract, which was chromatographed on silica gel. Elution with methanol in chloroform yielded two metabolites: 6α-hydroxyoxandrolone (4) (6 mg, 9% mol.) and 15α-hydroxyoxandrolone (5) (44 mg, 70% mol.). Compound 4 was as follows: resisted crystallization, = −57 (c 0.02, CHCl3) (lit.: = −62.5 (c 0.0008, CHCl3)) [21]; EI-MS m/z 323.2 [M]+ (16), 307.2 (41), 286.3 (42), 246.5 (75), 79.3 (40). 1H NMR (CDCl3, 600 MHz) δ (ppm): 0.86 (m, 1H, 9-H), 0.87 (s, 3H, 18-H), 0.97 (1H, m, 7β-H),1.02 (s, 3H, 19-H), 1.22 (s, 3H, 20-H), 1.28 (m, 1H, 14-H), 1.31 (m, 1H, 15β-H), 1.33 (m, 1H, 12β-H), 1.39 (m, 1H, 11α-H), 1.44 (m, 1H, 11α-H), 1.55 (m, 1H, 12α-H), 1.61 (m, 3H, 5-H, 8-H, 15α-H), 1.77 (m, 1H, 16α-H), 1.83 (m, 1H, 16β-H), 2.05 (dt, J = 4.1 Hz, J = 12.3 Hz, 1H, 7β-H); 2.34 (dd, J = 12.7 Hz, J = 19.0 Hz, 1H, 4β-H), 2.98 (dd, J = 6.0 Hz, J = 19.0 Hz, 1H, 4α-H), 3.45–3.49 (m, 6β-H), 3.93 (d, J = 10.7 Hz, 1H, 1α-H), 4.21 (d, J = 10.7 Hz, 1H, 1β-H); 13C NMR (CDCl3, 151 MHz) δ (ppm): 11.5 (C-19), 13.9 (C-18), 20.8 (C-11), 23.3 (C-15), 25.7 (C-20), 30.4 (C-4), 31.1 (C-12), 34.8 (C-8), 35.5 (C-10), 38.6 (C-16), 40.8 (C-7), 45.3 (C-13), 47.2 (C-5), 49.2 (C-9), 50.0 (C-14), 69.8 (C-6), 80.6 (C-1), 81.5 (C-17), 170.3 (C-3). See Figures S8–S11 for MS and NMR spectra of compound 4. (5): amorphous solid, = +39 (c 0.02, CHCl3); EI-MS m/z 323.0 [M]+ (40), 307.2 (17), 287.1 (31), 246.3 (60), 176.3 (95). 1H NMR and 13C NMR data are shown in Table 2 and Figures S12–S17.
4. Conclusions
Here we reported the relative resistance of oxandrolone (1) to extensive transformations by the tested strains of fungi. The comparison of the transformation results between compound 1 and other steroidal substrates [9,12] indicated that the structure of the A ring of the steroid molecule significantly influences the position of the introduced hydroxyl group, making these processes highly regioselective. The obtained results extend our knowledge of the steroid-transforming activities of microbial fungi. The high substrate conversion rates and achieved product yields indicate that these strains may be promising biocatalysts in the synthesis of new bioactive steroids which might be of interest in academia and clinical medicine in the future.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4344/11/1/16/s1, Figure S1: MS spectrum of 12β-hydroxyoxandrolone (2), Figure S2: 1H NMR spectrum of 12β-hydroxyoxandrolone (2), Figure S3: 13C NMR spectrum of 12β-hydroxyoxandrolone (2), Figure S4: NOESY spectrum of 12β-hydroxyoxandrolone (2), Figure S5: MS spectrum of 9α-hydroxyoxandrolone (3), Figure S6: 1H NMR spectrum of 9α-hydroxyoxandrolone (3), Figure S7: 13C NMR spectrum of 9α-hydroxyoxandrolone (3), Figure S8: MS spectrum of 6α-hydroxyoxandrolone (4), Figure S9: 1H NMR spectrum of 6α-hydroxyoxandrolone (4), Figure S10: 13C NMR spectrum of 6α-hydroxyoxandrolone (4), Figure S11: NOESY spectrum of 6α-hydroxyoxandrolone (4), Figure S12: MS spectrum of 15α-hydroxyoxandrolone (5), Figure S13: 1H NMR spectrum of 15α-hydroxyoxandrolone (5), Figure S14: 13C NMR spectrum of 15α-hydroxyoxandrolone (5), Figure S15: DEPT spectrum of 15α-hydroxyoxandrolone (5), Figure S16: HSQC spectrum of 15α-hydroxyoxandrolone (5), Figure S17: NOESY spectrum of 15α-hydroxyoxandrolone (5).
Author Contributions
Conceptualization, A.Ś. and P.Ł.; methodology, P.Ł.; formal analysis, A.Ś.; P.Ł. and A.P.; investigation, P.Ł.; data curation, P.Ł.; writing—original draft preparation A.Ś., P.Ł. and A.P.; writing—review and editing, A.Ś. and A.P.; supervision, A.Ś.; project administration, P.Ł.; funding acquisition, P.Ł. and A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Wrocław University of Environmental and Life Sciences (Poland) as the Ph.D. research program “Innowacyjny Doktorat”, no. N070/0011/20. Article Processing Charge (APC) was financed under the Leading Research Groups support project from the subsidy increased for the period 2020–2025 by the amount of 2% of the subsidy referred to Art. 387 (3) of the Law of 20 July 2018 on Higher Education and Science, obtained in 2019.
Data Availability Statement
Data available in a publicly accessible repository.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kristan, K.; Rižner, T.L. Steroid-transforming enzymes in fungi. J. Steroid Biochem. Mol. Biol. 2012, 129, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Świzdor, A.; Kołek, T.; Panek, A.; Milecka, N. Selective modifications of steroids performed by oxidative enzymes. Curr. Org. Chem. 2012, 16, 2551–2582. [Google Scholar] [CrossRef]
- Sedlaczek, L.; Smith, L.L. Biotransformations of steroids. Crit. Rev. Biotechnol. 1988, 7, 187–236. [Google Scholar] [CrossRef] [PubMed]
- Hogg, J.A. Steroids, the steroid community, and Upjohn in perspective: A profile of innovation. Steroids 1992, 57, 593–616. [Google Scholar] [CrossRef]
- Shull, G.M.; Kita, D.A. Microbiological conversion of steroids. I. Introduction of the 11β-hydroxyl group into C21 steroids. J. Am. Chem. Soc. 1955, 77, 763–764. [Google Scholar] [CrossRef]
- Świzdor, A.; Panek, A.; Milecka-Tronina, N. Hydroxylative activity of Aspergillus niger towards androst-4-ene and androst-5-ene steroids. Steroids 2017, 126, 101–106. [Google Scholar] [CrossRef]
- Mishra, R.; Mishra, S. Updates in bile acid-bioactive molecule conjugates and their applications. Steroids 2020, 159, 108639. [Google Scholar] [CrossRef]
- El Kihel, L. Oxidative metabolism of dehydroepiandrosterone (DHEA) and biologically active oxygenated metabolites of DHEA and epiandrosterone (EpiA)—Recent reports. Steroids 2012, 77, 10–26. [Google Scholar] [CrossRef]
- Kołek, T.; Milecka, N.; Świzdor, A.; Panek, A.; Białońska, A. Hydroxylation of DHEA, androstenediol and epiandrosterone by Mortierella isabellina AM212. Evidence indicating that both constitutive and inducible hydroxylases catalyze 7α- as well as 7β-hydroxylations of 5-ene substrates. Org. Biomol. Chem. 2011, 9, 5414–5422. [Google Scholar] [CrossRef]
- Milecka-Tronina, N.; Kołek, T.; Świzdor, A.; Panek, A. Hydroxylation of DHEA and its analogues by Absidia coerulea AM93. Can an inducible microbial hydroxylase catalyze 7α- and 7β-hydroxylation of 5-ene and 5α-dihydro C19-steroids? Bioorg. Med. Chem. 2014, 22, 883–891. [Google Scholar] [CrossRef]
- Meyer, W.; Gams, W. Delimination of Umbelopsis (Mucorales, Umbelopsidaceae fam. nov.) based on ITS sequence and RFLP data. Mycol. Res. 2003, 107, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kołek, T.; Świzdor, A. Biotransformation XLV. Transformations of 4-ene-3-oxo steroids in Fusarium culmorum culture. J. Steroid Biochem. Mol. Biol. 1998, 67, 63–69. [Google Scholar] [CrossRef]
- Kołek, T. Biotransformation XLVII: Transformations of 5-ene steroids in Fusarium culmorum culture. J. Steroid Biochem. Mol. Biol. 1999, 71, 83–90. [Google Scholar] [CrossRef]
- Świzdor, A.; Kołek, T. Transformations of 4- and 17α-substituted testosterone analogues by Fusarium Culmorum. Steroids 2005, 70, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Świzdor, A.; Kołek, T.; Szpineter, A. Transformations of steroid esters by Fusarium culmorum. Z. Nat. C 2006, 61, 809–814. [Google Scholar] [CrossRef] [Green Version]
- Pappo, R.; Jung, C.J. 2-Oxasteroids: A new class of biologically active compounds. Tetrahedron Lett. 1962, 3, 365–371. [Google Scholar] [CrossRef]
- Orr, R.; Singh, M.F. The anabolic androgenic steroid oxandrolone in the treatment of wasting and catabolic disorders: Review of efficacy and safety. Drugs 2004, 64, 725–750. [Google Scholar] [CrossRef]
- Kratena, N.; Stöger, B.; Weil, M.; Enev, V.S.; Gärtner, P. Synthesis of two epimeric long-term metabolites of oxandrolone. Tetrahedron Lett. 2017, 58, 1316–1318. [Google Scholar] [CrossRef]
- Foster, M.A.; Taylor, A.E.; Hill, N.E.; Bentley, C.; Bishop, J.; Gilligan, L.C.; Shaheen, F.; Bion, J.F.; Fallowfield, J.L.; Woods, D.R.; et al. Mapping the steroid response to major trauma from injury to recovery: A prospective cohort study. J. Clin. Endocrinol. Metab. 2020, 105, 925–937. [Google Scholar] [CrossRef] [Green Version]
- Ring, J.; Heinelt, M.; Sharma, S.; Letourneau, S.; Jeschke, M.G. Oxandrolone in the treatment of burn injuries: A systematic review and meta-analysis. J. Burn. Care Res. 2020, 41, 190–199. [Google Scholar] [CrossRef]
- Choudhary, M.I.; Mohammad, M.Y.; Musharraf, S.G.; Parvez, M.; Al-Aboudi, A.; Atta-ur-Rahman. New oxandrolone derivatives by biotransformation using Rhizopus stolonifer. Steroids 2009, 74, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Guddat, S.; Fußhöller, G.; Beuck, S.; Thomas, A.; Geyer, H.; Rydevik, A.; Bondesson, U.; Hedeland, M.; Lagojda, A.; Schänzer, W.; et al. Synthesis, characterization, and detection of new oxandrolone metabolites as long-term markers in sports drug testing. Anal. Bioanal. Chem. 2013, 405, 8285–8294. [Google Scholar] [CrossRef] [PubMed]
- Rzeppa, S.; Viet, L. Analysis of sulfate metabolites of the doping agents oxandrolone and danazol using high performance liquid chromatography coupled to tandem mass spectrometry. J. Chromatogr. B 2016, 1029, 1–9. [Google Scholar] [CrossRef] [PubMed]
- World Anti-Doping Agency Prohibited List. 2020. Available online: https://www.wada-ama.org/sites/default/files/wada_2020_english_prohibited_list_0.pdf (accessed on 11 November 2020).
- Massé, R.; Bi, H.; Ayotte, C.; Dugal, R. Studies on anabolic steroids II—gas chromatographic/mass spectrometric characterization of oxandrolone urinary metabolites in man. Biomed. Environ. Mass Spectrom. 1989, 18, 429–438. [Google Scholar] [CrossRef]
- Desoky, E.-S.I.; Reyad, M.; Afsah, E.M.; Dawidar, A.-A.M. Synthesis and chemical reactions of the steroidal hormone 17α-methyltestosterone. Steroids 2016, 105, 68–95. [Google Scholar] [CrossRef] [PubMed]
- Janeczko, T.; Popłoński, J.; Kozłowska, E.; Dymarska, M.; Huszcza, E.; Kostrzewa-Susłow, E. Application of α- and β-naphthoflavones as monooxygenase inhibitors of Absidia coerulea KCh 93, Syncephalastrum racemosum KCh 105 and Chaetomium sp. KCh 6651 in transformation of 17α-methyltestosterone. Bioorg. Chem. 2018, 78, 178–184. [Google Scholar] [CrossRef]
- Smith, C.; Atia-Tul-Wahab; Khan, M.S.A.; Ahmad, M.S.; Farran, D.; Choudhary, M.I.; Baydoun, E. Microbial transformation of oxandrolone with Macrophomina phaseolina and Cunninghamella blakesleeana. Steroids 2015, 102, 39–45. [Google Scholar] [CrossRef]
- Świzdor, A.; Milecka, N.; Kołek, T.; Panek, A. Process for the Preparation of 3β,15α-dihydroxy-5α-androst-17-one. Polish Patent PL 215785 B1, 31 January 2014. [Google Scholar]
Figure 1.
Transformation of oxandrolone (1) in the culture of Fusarium culmorum AM282.
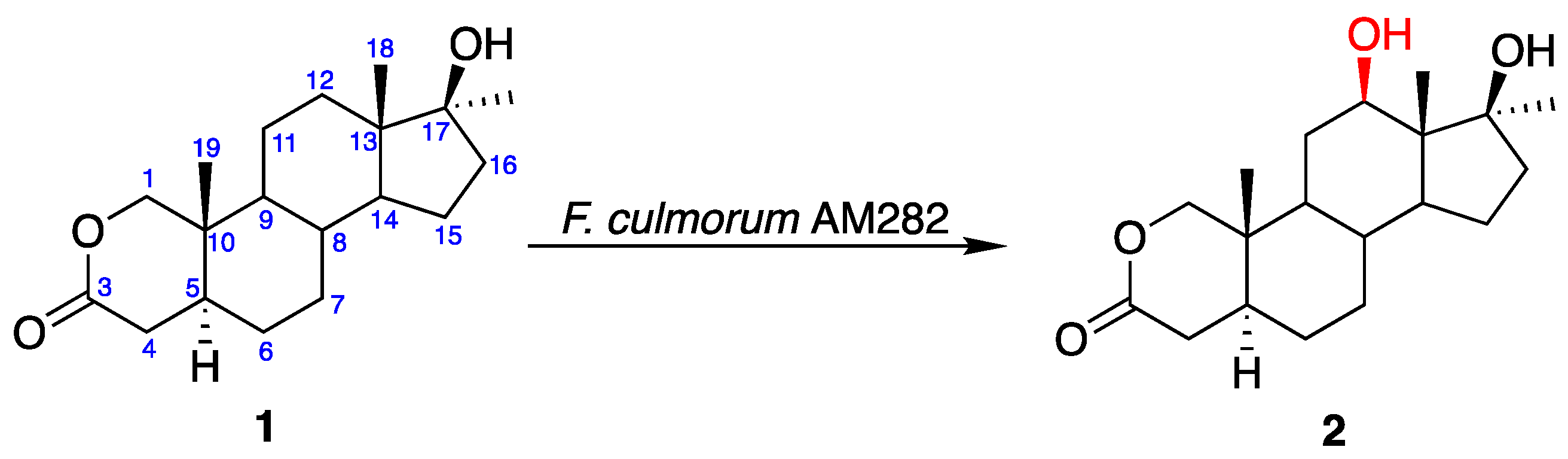
Figure 2.
Transformation of oxandrolone (1) in the culture of Mortierella isabellina AM212.

Figure 3.
Transformation of oxandrolone (1) in the culture of Laetiporus sulphureus AM498.

Figure 4.
Key NOESY correlations for 15α-hydroxyoxandrolone (5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Compositions of crude extracts obtained in transformations of oxandrolone (1).
| Strain | Compounds Present in the Mixture | Rt (min.) | Time of Transformation (Days) | ||||
|---|---|---|---|---|---|---|---|
| 1 | 3 | 5 | 7 | 10 | |||
| (%) a | |||||||
| Fusarium culmorum AM282 | Oxandrolone (1) | 7.14 | 84 | 55 | 26 | 22 | - |
| 12β-Hydroxyoxandrolone (2) | 10.06 | 16 | 38 | 66 | 69 | - | |
| Mortierella isabellina AM212 | Oxandrolone (1) | 7.14 | 90 | 40 | 19 | - | - |
| 9α-Hydroxyoxandrolone (3) | 9.15 | 10 | 48 | 69 | 80 | 78 | |
| Laetiporus sulphureus AM498 | Oxandrolone (1) | 7.14 | 93 | 82 | 48 | 23 | - |
| 6α-Hydroxyoxandrolone (4) | 10.24 | - | - | 8 | 12 | 18 | |
| 15α-Hydroxyoxandrolone (5) | 9.83 | 7 | 15 | 39 | 61 | 76 | |
a Determined by GC analysis.
Table 2.
Spectroscopic data of 15α-hydroxyoxandrolone (5) *.
| Position | δC, | Type | δH, Multiplicity (J in Hz) | NOESY (H → H) a |
|---|---|---|---|---|
| 1α | 80.9, | CH2 | 3.93, d (10.7) | 5, 9 |
| 1β | 4.22, d (10.7) | 11, 19 | ||
| 3 | 170.4, | C | - | - |
| 4α | 33.7, | CH2 | 2.53, dd (5.9; 18.8) | 6 |
| 4β | 2.22, dd (13.0; 18.8) | 6, 8, 19 | ||
| 5 | 40.2, | CH | 1.72, m | 1, 7, 9 |
| 6α | 27.0, | CH2 | 1.51, m | 4 |
| 6β | 1.21 b, m | 4, 8, 19, | ||
| 7α | 31.3, | CH2 | 1.14, m | 5, 9, 14 |
| 7β | 2.02, dm | 6, 8, 15 | ||
| 8 | 35.1, | CH | 1.68, m | 6, 11, 18, 19 |
| 9 | 49.5, | CH | 0.85, m | 1, 5, 7, 12, 14 |
| 10 | 34.7, | C | - | - |
| 11α | 20.8, | CH2 | 1.46 c, m | 9 |
| 11β | 1.46 c, dm | 8, 18, 19 | ||
| 12α | 31.2, | CH2 | 1.51, dm | 9, 14, 20 |
| 12β | 1.37, m | 11, 18 | ||
| 13 | 46.5, | C | - | - |
| 14 | 57.9, | CH | 1.24 b, m | 9, 12, 20 |
| 15 | 72.3, | CH | 4.07 (td, 3.4; 9.3) | 7, 8, 11, 16, 18 |
| 16α | 50.3, | CH2 | 1,64, dd (3.4; 14.6) | 14, 20 |
| 16β | 2.42, dd (9.7; 14.6) | 8, 15, 18 | ||
| 17 | 79.1, | C | - | - |
| 18 | 15.4, | CH3 | 0.87, s | 8, 11, 19 |
| 19 | 10.2, | CH3 | 1.02, s | 1, 4, 6, 8, 11 |
| 20 | 26.1, | CH3 | 1.35, s | 12, 14, 16 |
* 600 MHz for 1H NMR and 151 MHz for 13C NMR. (a) NOESY correlations are from the stated proton to the indicated proton(s). (b,c) Signals are interchangeable.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Łyczko, P.; Panek, A.; Świzdor, A. Highly Regioselective and Stereoselective Biohydroxylations of Oxandrolone. Catalysts 2021, 11, 16. https://doi.org/10.3390/catal11010016
AMA Style
Łyczko P, Panek A, Świzdor A. Highly Regioselective and Stereoselective Biohydroxylations of Oxandrolone. Catalysts. 2021; 11(1):16. https://doi.org/10.3390/catal11010016
Chicago/Turabian StyleŁyczko, Paulina, Anna Panek, and Alina Świzdor. 2021. "Highly Regioselective and Stereoselective Biohydroxylations of Oxandrolone" Catalysts 11, no. 1: 16. https://doi.org/10.3390/catal11010016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.