
Identification of Cytochrome P450 Enzymes Responsible for Oxidative Metabolism of Synthetic Cannabinoid (1-Hexyl-1H-Indol-3-yl)-1-naphthalenyl-methanone (JWH-019)

 ,
,
Abstract
:1. Introduction
2. Results
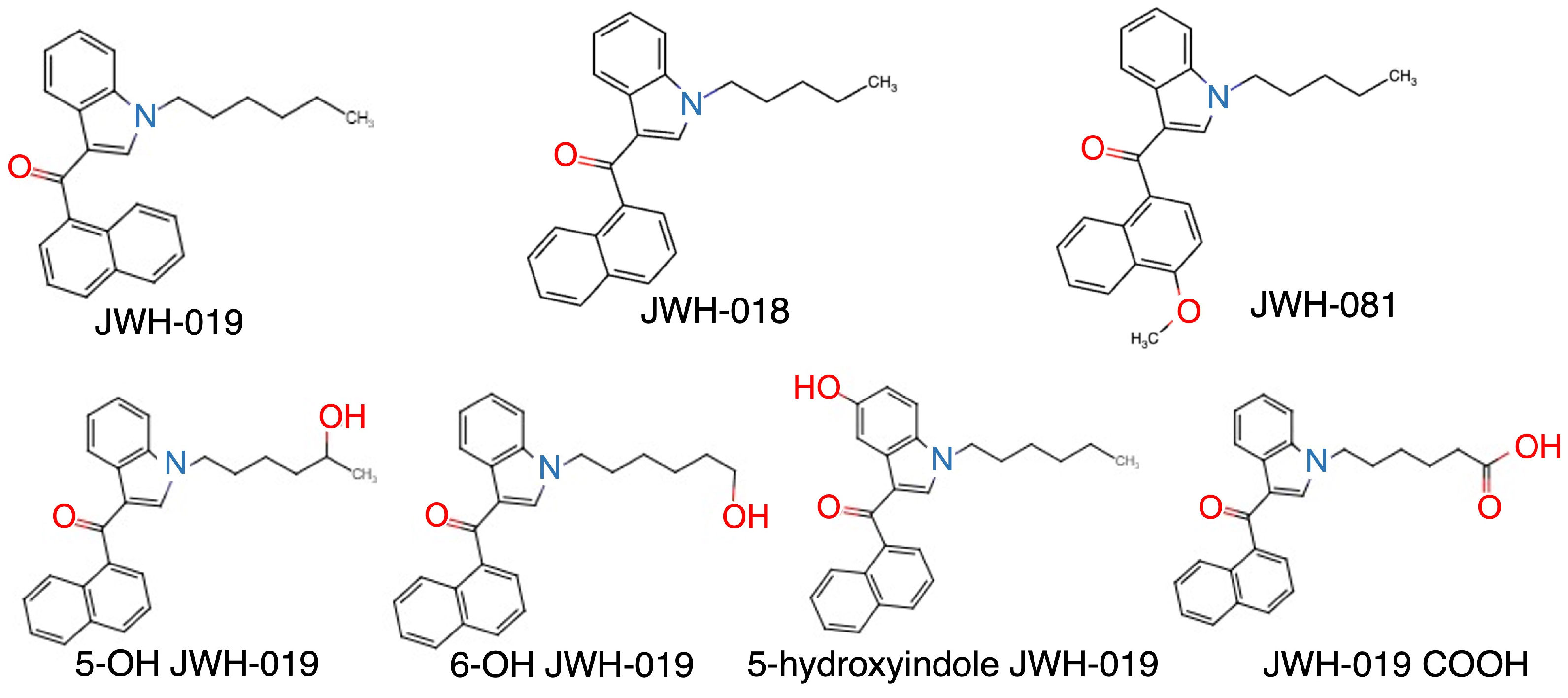
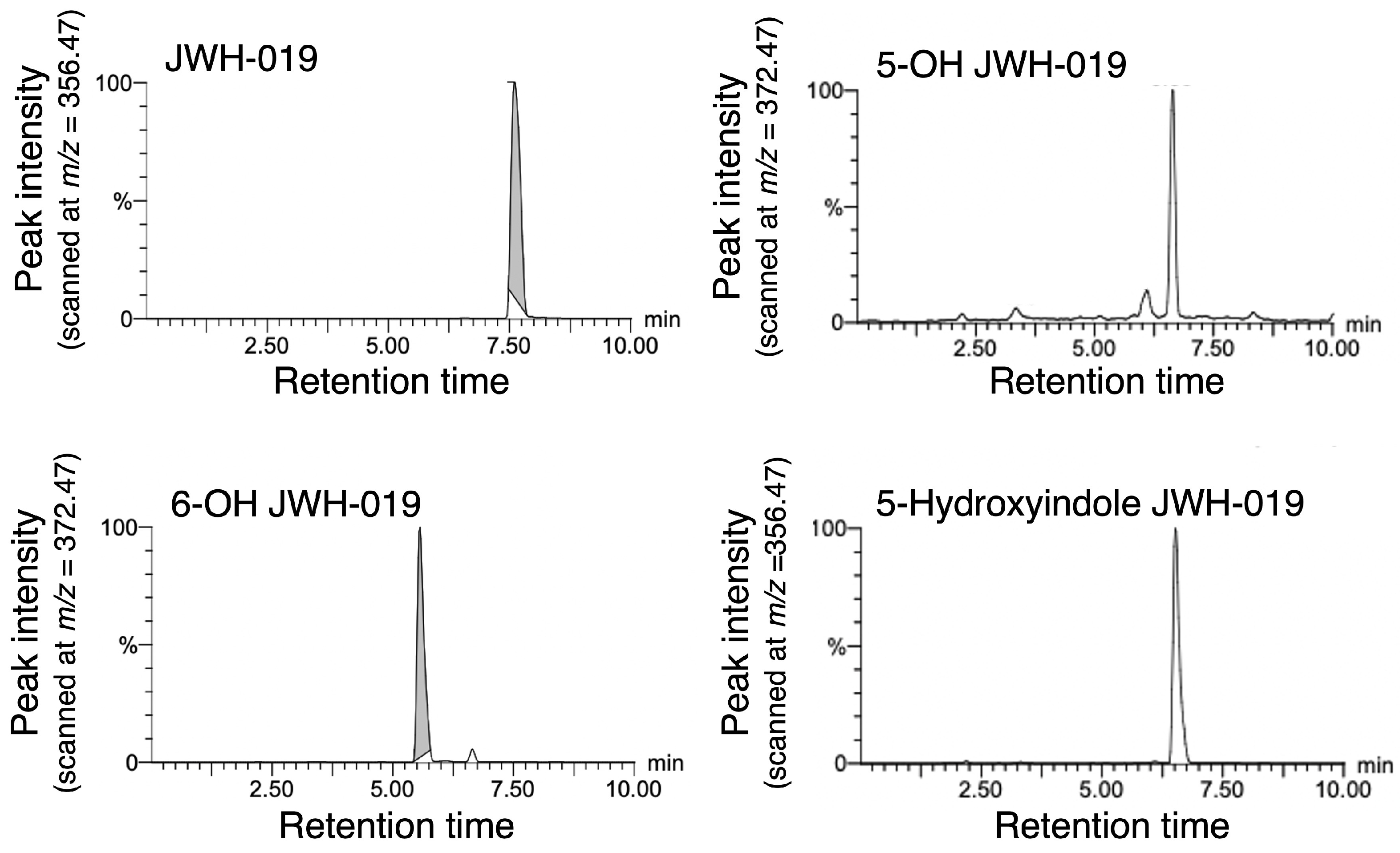
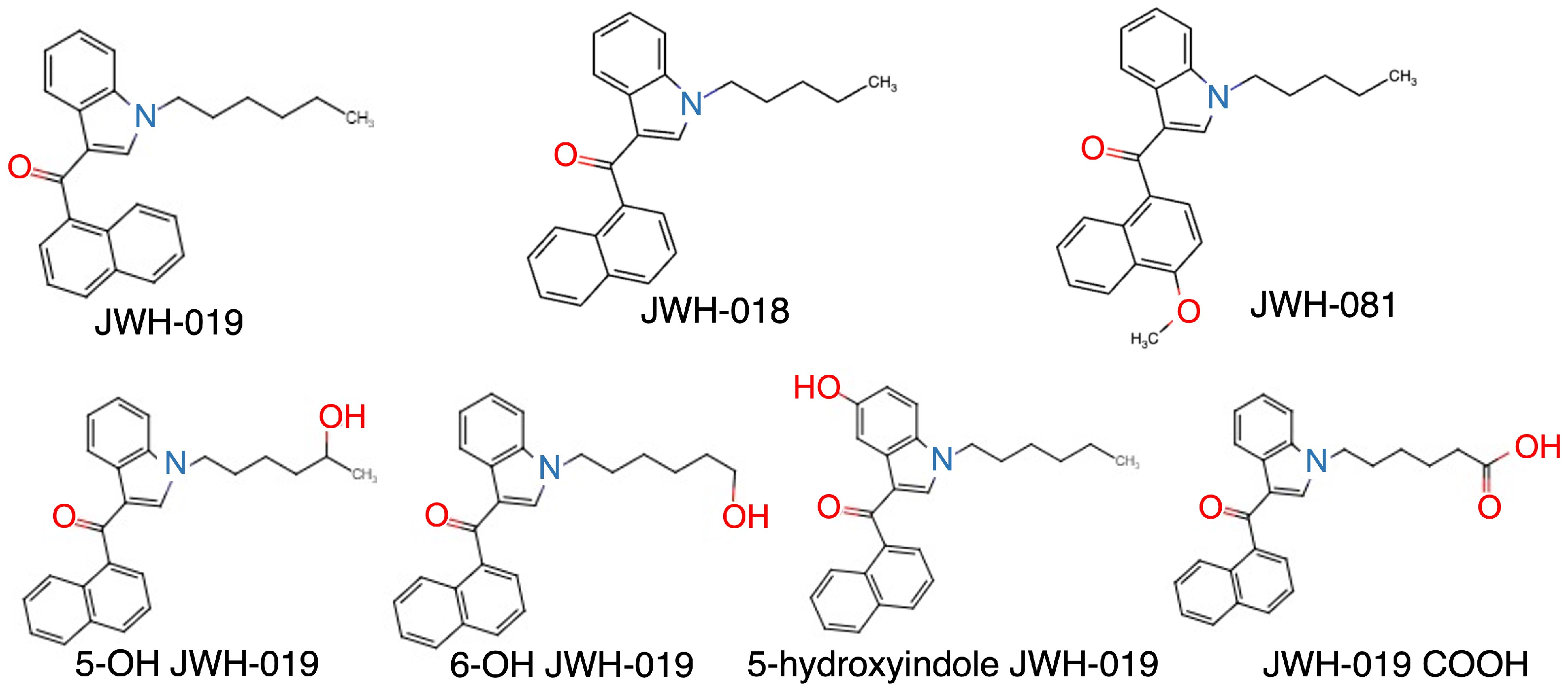
2.1. Development of a UPLC Method for JWH-019 and Its Metabolites
2.2. Kinetic Analysis of 6-OH JWH-019 Formation in HLMs
2.3. Identification of JWH-019-Oxidating Enzymes
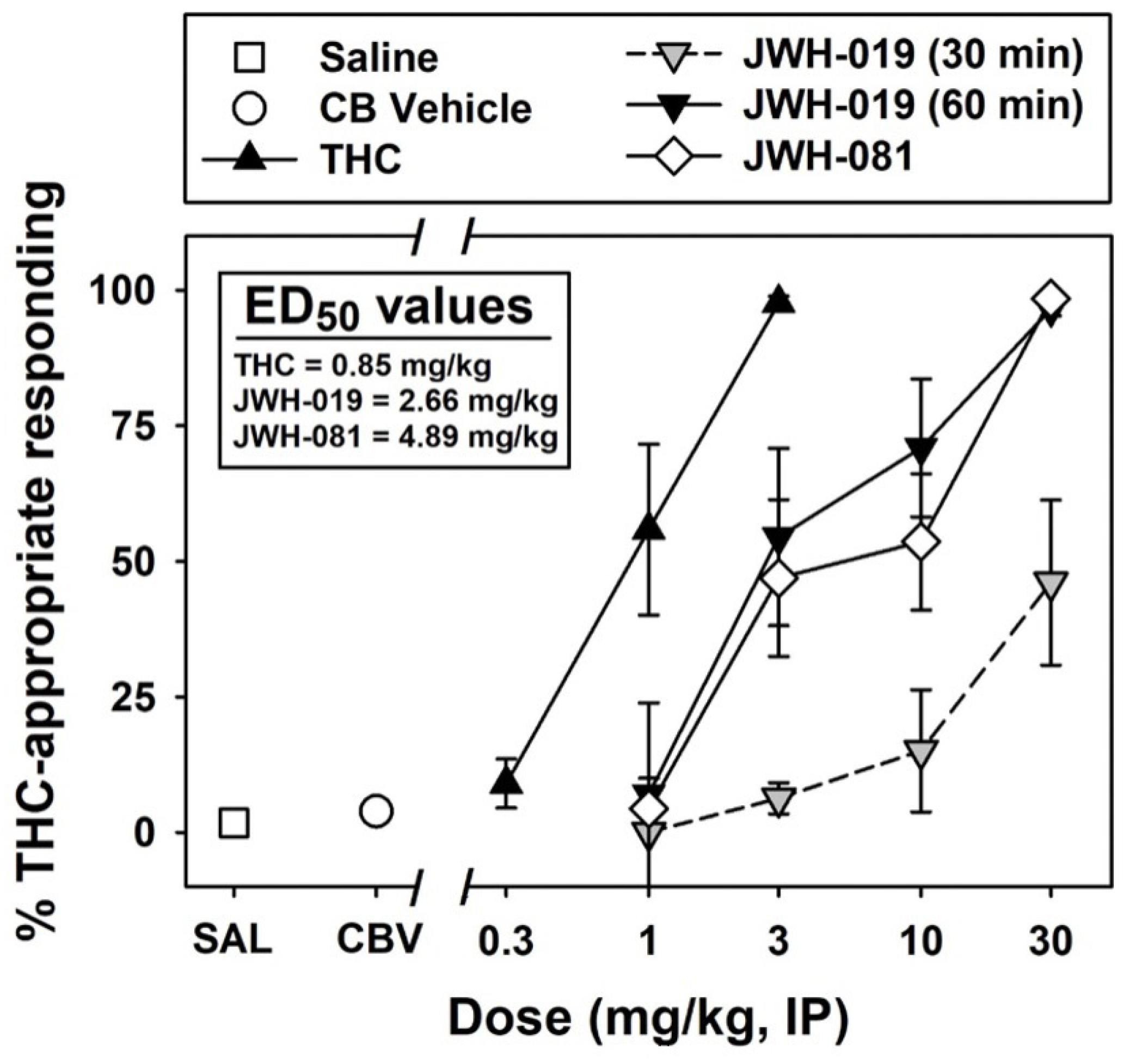
2.4. Pharmacological Activity of JWH-019
3. Materials and Methods
3.1. Materials
3.2. Separation and Detection of JWH-019 and Its Metabolites
3.3. Metabolism of JWH-019 in HLMs and Recombinant P450s
3.4. Kinetic Assay and Data Analysis
3.5. Assessment of the Pharmacological Activity
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasin, D.S.; Saha, T.D.; Kerridge, B.T.; Goldstein, R.B.; Chou, S.P.; Zhang, H.; Jung, J.; Pickering, R.P.; Ruan, W.J.; Smith, S.M.; et al. Prevalence of marijuana use disorders in the United States between 2001–2002 and 2012–2013. JAMA Psychiatry 2015, 72, 1235–1242. [Google Scholar] [CrossRef]
- Howlett, A.C.; Abood, M.E. CB1 and CB2 Receptor Pharmacology. Adv. Pharmacol. 2017, 80, 169–206. [Google Scholar]
- Schneir, A.B.; Cullen, J.; Ly, B.T. “Spice” girls: Synthetic cannabinoid intoxication. J. Emerg. Med. 2011, 40, 296–299. [Google Scholar] [CrossRef]
- Palamar, J.J.; Acosta, P. Synthetic cannabinoid use in a nationally representative sample of US high school seniors. Drug. Alcohol. Depend. 2015, 149, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winstock, A.R.; Barratt, M.J. Synthetic cannabis: A comparison of patterns of use and effect profile with natural cannabis in a large global sample. Drug. Alcohol. Depend. 2013, 131, 106–111. [Google Scholar] [CrossRef]
- Lapoint, J.; James, L.P.; Moran, C.L.; Nelson, L.S.; Hoffman, R.S.; Moran, J.H. Severe toxicity following synthetic cannabinoid ingestion. Clin. Toxicol. 2011, 49, 760–764. [Google Scholar] [CrossRef]
- Law, R.; Schier, J.; Martin, C.; Chang, A.; Wolkin, A. Increase in reported adverse health effects related to synthetic cannabinoid use—United States, January–May 2015. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 618. [Google Scholar]
- Kasper, A.M.; Ridpath, A.D.; Gerona, R.R.; Cox, R.; Galli, R.; Kyle, P.B.; Parker, C.; Arnold, J.K.; Chatham-Stephens, K.; Morrison, M.A.; et al. Severe illness associated with reported use of synthetic cannabinoids: A public health investigation (Mississippi, 2015). Clin. Toxicol. 2019, 57, 10–18. [Google Scholar] [CrossRef]
- American Association of Poison Control Centers. Synthetic Cannabinoid Data. 23 September 2015. Available online: http://www.aapcc.org/ (accessed on 1 March 2022).
- Connors, J.M. Hemorrhagic highs from synthetic cannabinoids–A new epidemic. N. Engl. J. Med. 2018, 379, 1275–1277. [Google Scholar] [CrossRef] [PubMed]
- Shanks, K.G.; Dahn, T.; Behonick, G.; Terrell, A. Analysis of First and Second Generation Legal Highs for Synthetic Cannabinoids and Synthetic Stimulants by Ultra-Performance Liquid Chromatography and Time of Flight Mass Spectrometry. J. Anal. Toxicol. 2012, 36, 360–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, B.K.; Reinhold, L.E.; Xu, A.; Diamond, F.X. Identification of synthetic cannabinoids in herbal incense blends in the United States. J. Forensic Sci. 2012, 57, 1168–1180. [Google Scholar] [CrossRef]
- Müller, H.H.; Kornhuber, J.; Sperling, W. The behavioral profile of spice and synthetic cannabinoids in humans. Brain Res. Bull. 2016, 126, 3–7. [Google Scholar] [CrossRef]
- Hermanns-Clausen, M.; Kneisel, S.; Szabo, B.; Auwärter, V. Acute toxicity due to the confirmed consumption of synthetic cannabinoids: Clinical and laboratory findings. Addiction 2013, 108, 534–544. [Google Scholar] [CrossRef]
- Musshoff, F.; Madea, B.; Kernbach-Wighton, G.; Bicker, W.; Kneisel, S.; Hutter, M.; Auwärter, V. Driving under the influence of synthetic cannabinoids (“Spice”): A case series. Int. J. Legal Med. 2014, 128, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Anzillotti, L.; Marezza, F.; Calò, L.; Banchini, A.; Cecchi, R. A case report positive for synthetic cannabinoids: Are cardiovascular effects related to their protracted use? Leg. Med. 2019, 41, 101637. [Google Scholar] [CrossRef] [PubMed]
- Brents, L.K.; Prather, P.L. The K2/Spice phenomenon: Emergence, identification, legislation and metabolic characterization of synthetic cannabinoids in herbal incense products. Drug. Metab. Rev. 2014, 46, 72–85. [Google Scholar] [CrossRef] [Green Version]
- Rendic, S. Summary of information on human CYP enzymes: Human P450 metabolism data. Drug. Metab. Rev. 2002, 34, 83–448. [Google Scholar] [CrossRef]
- Fisher, M.B.; Paine, M.F.; Strelevitz, T.J.; Wrighton, S.A. The role of hepatic and extrahepatic UDP-glucuronosyltransferases in human drug metabolism. Drug. Metab. Rev. 2001, 33, 273–297. [Google Scholar] [CrossRef]
- Hirth, J.; Watkins, P.B.; Strawderman, M.; Schott, A.; Bruno, R.; Baker, L.H. The effect of an individual’s cytochrome CYP3A4 activity on docetaxel clearance. Clin. Cancer Res. 2000, 6, 1255–1258. [Google Scholar] [PubMed]
- Ding, X.; Kaminsky, L.S. Human extrahepatic cytochromes P450: Function in xenobiotic metabolism and tissue-selective chemical toxicity in the respiratory and gastrointestinal tracts. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 149–173. [Google Scholar] [CrossRef]
- Vikingsson, S.; Josefsson, M.; Gréen, H. Identification of AKB-48 and 5F-AKB-48 Metabolites in Authentic Human Urine Samples Using Human Liver Microsomes and Time of Flight Mass Spectrometry. J. Anal. Toxicol. 2015, 39, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Scheidweiler, K.B.; Wohlfarth, A.; Pang, S.; Kronstrand, R.; Huestis, M.A. In Vitro and In Vivo Human Metabolism of Synthetic Cannabinoids FDU-PB-22 and FUB-PB-22. AAPS J. 2016, 18, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Patton, A.L.; Seely, K.A.; Yarbrough, A.L.; Fantegrossi, W.; James, L.P.; McCain, K.R.; Fujiwara, R.; Prather, P.L.; Moran, J.H.; Radominska-Pandya, A. Altered metabolism of synthetic cannabinoid JWH-018 by human cytochrome P450 2C9 and variants. Biochem. Biophys. Res. Commun. 2018, 498, 597–602. [Google Scholar] [CrossRef]
- Aung, M.M.; Griffin, G.; Huffman, J.W.; Wu, M.; Keel, C.; Yang, B.; Showalter, V.M.; Abood, M.E.; Martin, B.R. Influence of the N-1 alkyl chain length of cannabimimetic indoles upon CB(1) and CB(2) receptor binding. Drug. Alcohol. Depend. 2000, 60, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Hutter, M.; Broecker, S.; Kneisel, S.; Franz, F.; Brandt, S.D.; Auwärter, V. Metabolism of synthetic cannabinoid receptor agonists encountered in clinical casework: Major in vivo phase I metabolites of JWH-007, JWH-019, JWH-203, JWH-307, UR-144, XLR-11, AM-2201, MAM-2201 and AM-694 in human urine using LC-MS/MS. Curr. Pharm. Biotechnol. 2018, 19, 144–162. [Google Scholar] [CrossRef] [PubMed]
- Jonsson-Schmunk, K.; Schafer, S.C.; Croyle, M.A. Impact of nanomedicine on hepatic cytochrome P450 3A4 activity: Things to consider during pre-clinical and clinical studies. J. Pharm. Investig. 2018, 48, 113–134. [Google Scholar] [CrossRef]
- Wu, Z.; Lee, D.; Joo, J.; Shin, J.H.; Kang, W.; Oh, S.; Lee, D.Y.; Lee, S.J.; Yea, S.S.; Lee, H.S.; et al. CYP2J2 and CYP2C19 are the major enzymes responsible for metabolism of albendazole and fenbendazole in human liver microsomes and recombinant P450 assay systems. Antimicrob. Agents Chemother. 2013, 57, 5448–5456. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.; Yarbrough, A.L.; Fantegrossi, W.E.; Prather, P.L.; Bush, J.M.; Radominska-Pandya, A.; Fujiwara, R. Identifying cytochrome P450s involved in oxidative metabolism of synthetic cannabinoid N-(adamantan-1-yl)-1-(5-fluoropentyl)-1H-indole-3-carboxamide (STS-135). Pharmacol. Res. Perspect. 2020, 8, e00561. [Google Scholar] [CrossRef] [Green Version]
- Gannon, B.M.; Williamson, A.; Suzuki, M.; Rice, K.C.; Fantegrossi, W.E. Stereoselective Effects of Abused “Bath Salt” Constituent 3,4-Methylenedioxypyrovalerone in Mice: Drug Discrimination, Locomotor Activity, and Thermoregulation. J. Pharmacol. Exp. Ther. 2016, 356, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Hiranita, T.; Soto, P.L.; Tanda, G.; Katz, J.L. Lack of cocaine-like discriminative-stimulus effects of σ-receptor agonists in rats. Behav. Pharmacol. 2011, 22, 525–530. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 6th ed.; Iowa State University Press: Ames, IA, USA, 1967. [Google Scholar]
- Keyes, K.M.; Rutherford, C.; Hamilton, A.; Palamar, J.J. Age, period, and cohort effects in synthetic cannabinoid use among US adolescents, 2011–2015. Drug. Alcohol. Depend. 2016, 166, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, D.M.; Woodwell, D.A. Update: Drug-related emergency department visits involving synthetic cannabinoids. In The CBHSQ Report; Substance Abuse and Mental Health Services Administration (US): Rockville, MD, USA, 2014. [Google Scholar]
- Wintermeyer, A.; Moller, I.; Thevis, M.; Jubner, M.; Beike, J.; Rothschild, M.A.; Bender, K. In vitro phase I metabolism of the synthetic cannabimimetic JWH-018. Anal. Bioanal. Chem. 2010, 398, 2141–2153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Y.; Dunbar, D.; Ostrowska, A.; Zeisloft, S.; Yang, J.; Kaminsky, L.S. Characterization of human small intestinal cytochromes P-450. Drug Metab. Dispos. 1999, 27, 804–809. [Google Scholar] [PubMed]
- Di, L. The Impact of Carboxylesterases in Drug Metabolism and Pharmacokinetics. Curr. Drug. Metab. 2019, 20, 91–102. [Google Scholar] [CrossRef]
- Rogers, S.; de Souza, A.R.; Zago, M.; Iu, M.; Guerrina, N.; Gomez, A.; Matthews, J.; Baglole, C.J. Aryl hydrocarbon receptor (AhR)-dependent regulation of pulmonary miRNA by chronic cigarette smoke exposure. Sci. Rep. 2017, 7, 40539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laika, B.; Leucht, S.; Heres, S.; Schneider, H.; Steimer, W. Pharmacogenetics and olanzapine treatment: CYP1A2*1F and serotonergic polymorphisms influence therapeutic outcome. Pharmacogenomics J. 2010, 10, 20–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delozier, T.C.; Kissling, G.E.; Coulter, S.J.; Dai, D.; Foley, J.F.; Bradbury, J.A.; Murphy, E.; Steenbergen, C.; Zeldin, D.C.; Goldstein, J.A. Detection of human CYP2C8, CYP2C9, and CYP2J2 in cardiovascular tissues. Drug. Metab. Dispos. 2007, 35, 682–688. [Google Scholar] [CrossRef] [Green Version]
- Evangelista, E.A.; Kaspera, R.; Mokadam, N.A.; Jones, J.P., 3rd; Totah, R.A. Activity, inhibition, and induction of cytochrome P450 2J2 in adult human primary cardiomyocytes. Drug. Metab. Dispos. 2013, 41, 2087–2094. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Activity (pmol/min/nmol P450) | Abundance (%) | Contribution (%) | |
|---|---|---|---|
| CYP1A2 | 2359 ± 68 | 12.1 | 33.0 ± 0.9 |
| CYP2A6 | 25 ± 7 | 8.3 | 0.25 ± 0.07 |
| CYP2B6 | 395 ± 48 | 2.5 | 1.14 ± 0.14 |
| CYP2C8 | 1238 ± 17 | 5.5 | 7.86 ± 0.11 |
| CYP2C9 | 559 ± 17 | 16.8 | 10.8 ± 0.3 |
| CYP2C19 | 4195 ± 43 | 3.2 | 15.5 ± 0.2 |
| CYP2D6 | 1637 ± 65 | 1.8 | 3.40 ± 0.14 |
| CYP2E1 | 2.8 ± 0.6 | 14.1 | 0.05 ± 0.01 |
| CYP2J2 | 8000 ± 334 | 1.16 | 10.7 ± 0.4 |
| CYP3A4 | 366 ± 8 | 30.5 | 12.9 ± 0.3 |
| CYP3A5 | 2014 ± 104 | 1.9 | 4.41 ± 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, N.; Fantegrossi, W.E.; McCain, K.R.; Wang, X.; Fujiwara, R. Identification of Cytochrome P450 Enzymes Responsible for Oxidative Metabolism of Synthetic Cannabinoid (1-Hexyl-1H-Indol-3-yl)-1-naphthalenyl-methanone (JWH-019). Catalysts 2023, 13, 1008. https://doi.org/10.3390/catal13061008
Tran N, Fantegrossi WE, McCain KR, Wang X, Fujiwara R. Identification of Cytochrome P450 Enzymes Responsible for Oxidative Metabolism of Synthetic Cannabinoid (1-Hexyl-1H-Indol-3-yl)-1-naphthalenyl-methanone (JWH-019). Catalysts. 2023; 13(6):1008. https://doi.org/10.3390/catal13061008
Chicago/Turabian StyleTran, Ngoc, William E. Fantegrossi, Keith R. McCain, Xinwen Wang, and Ryoichi Fujiwara. 2023. "Identification of Cytochrome P450 Enzymes Responsible for Oxidative Metabolism of Synthetic Cannabinoid (1-Hexyl-1H-Indol-3-yl)-1-naphthalenyl-methanone (JWH-019)" Catalysts 13, no. 6: 1008. https://doi.org/10.3390/catal13061008