Fluorescein Diacetate Hydrolysis Using the Whole Biofilm as a Sensitive Tool to Evaluate the Physiological State of Immobilized Bacterial Cells


Abstract
:1. Introduction
2. Results and Discussion
2.1. Fluorescein Adsorption by Polyurethane Foam (PUR)
2.2. Fluorescein Diacetate Application and Impact of Shaking
2.3. pH Optimization
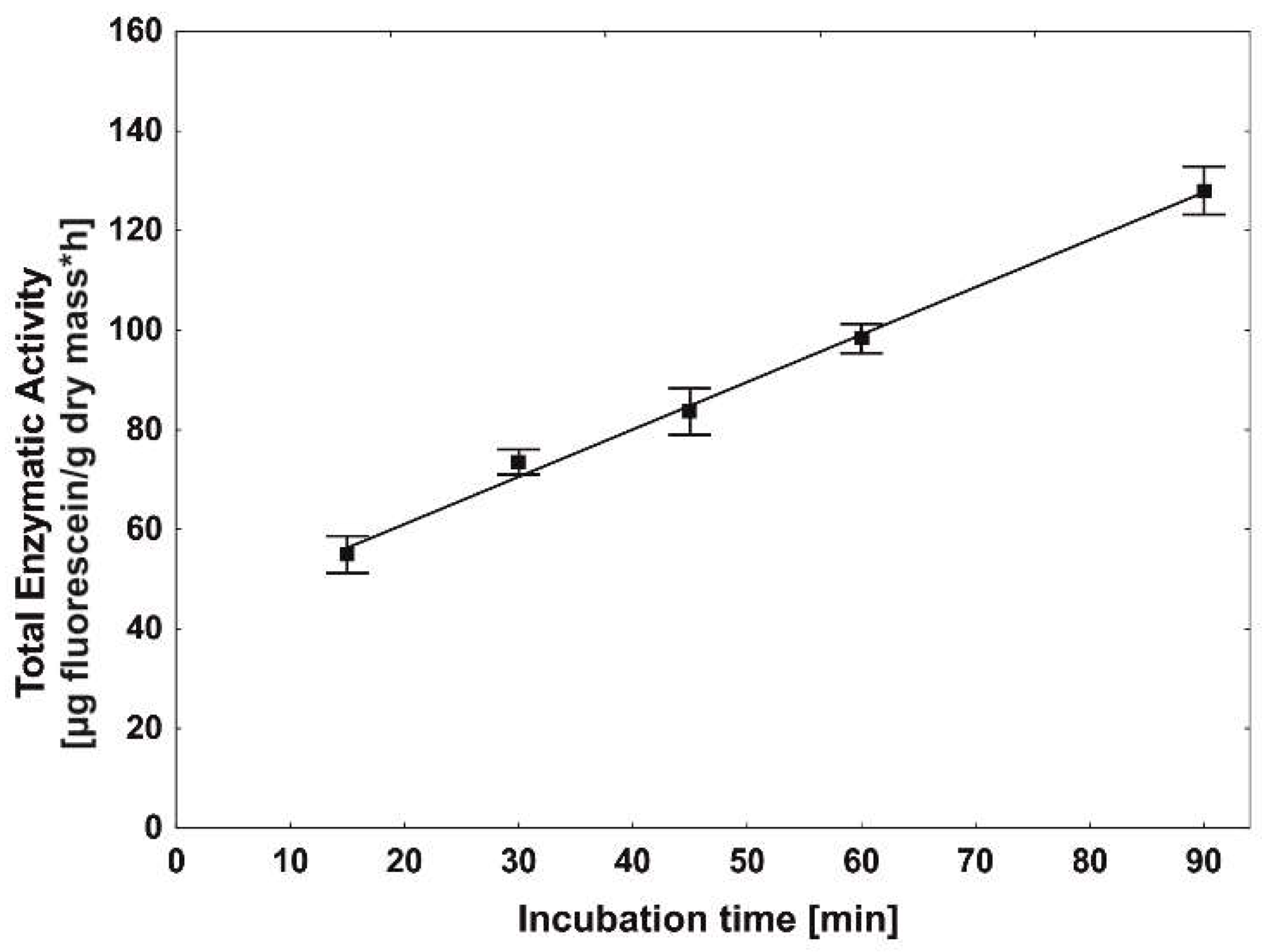
2.4. Incubation Time
2.5. Sensitivity Assay—Carbon Starvation
2.6. Comparision of the Modified FDA Method with Oxygen Consumption
3. Materials and Methods
3.1. Materials
3.2. Bacterial Strains and Growth Conditions
3.3. Immobilization Procedure
3.4. Standard Method of Non-Specific Esterase Activity with FDA Assay
3.5. Abiotic Controls for FDA Assay
3.6. Optimization Procedure
3.7. Modified Method of Non-Specific Esterase Activity with FDA Assay
3.8. Sensitivity Assay—Carbon Starvation
3.9. Scanning Electron Microscopy
3.10. Oxygen Consumption
3.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wojcieszyńska, D.; Domaradzka, D.; Hupert-Kocurek, K.; Guzik, U. Enzymes Involved in Naproxen Degradation by Planococcus sp. S5. Pol. J. Microbiol. 2016, 65, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Dzionek, A.; Wojcieszyńska, D.; Guzik, U. Natural carriers in bioremediation: A review. Electron. J. Biotechnol. 2016, 19, 28–36. [Google Scholar] [CrossRef]
- Bayat, Z.; Hassanshahian, M.; Cappello, S. Immobilization of microbes for bioremediation of crude oil polluted environments: A mini review. Open Microbiol. J. 2015, 9, 48–54. [Google Scholar] [PubMed]
- Dzionek, A.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Adamczyk-Habrajska, M.; Guzik, U. Immobilization of Planococcus sp. S5 strain on the loofah sponge and its application in naproxen removal. Catalysts 2018, 8, 176. [Google Scholar] [CrossRef]
- Partovinia, A.; Rasekh, B. Review of the immobilized microbial cell systems for bioremediation of petroleum hydrocarbons polluted environments. Crit. Rev. Environ. Sci. Technol. 2018, 48, 1–38. [Google Scholar] [CrossRef]
- Sarioglu, O.F.; Celebioglu, A.; Tekinay, T.; Uyar, T. Evaluation of fiber diameter and morphology differences for electrospun fibers on bacterial immobilization and bioremediation performance. Int. Biodeterior. Biodegradation 2017, 120, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Mrozik, A.; Piotrowska-Seget, Z. Bioaugmentation as a strategy for cleaning up of soils contaminated with aromatic compounds. Microbiol. Res. 2010, 165, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Alessandrello, M.J.; Parellada, E.A.; Juárez Tomás, M.S.; Neske, A.; Vullo, D.L.; Ferrero, M.A. Polycyclic aromatic hydrocarbons removal by immobilized bacterial cells using annonaceous acetogenins for biofilm formation stimulation on polyurethane foam. J. Environ. Chem. Eng. 2017, 5, 189–195. [Google Scholar] [CrossRef]
- Ohashi, A.; Harada, H. Adhesion strength of biofilm developed in an attached-growth reactor. Water Sci. Technol. 1994, 29, 281–288. [Google Scholar] [CrossRef]
- Stanley, P.M. Factors affecting the irreversible attachment of Pseudomonas aeruginosa to stainless steel. Can. J. Microbiol. 1983, 29, 1493–1499. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, I.W. The biofilm matrix—An immobilized but dynamic microbial environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Niknezhad, S.V.; Asadollahi, M.A.; Zamani, A.; Biria, D. Production of xanthan gum by free and immobilized cells of xanthomonas campestris and xanthomonas pelargonii. Int. J. Biological Macromol. 2016, 82, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Nie, H.; He, M.; Lin, Y.; Wang, L.; Jin, P.; Zhang, S. Immobilization of biofilms of pseudomonas aeruginosa ny3 and their application in the removal of hydrocarbons from highly concentrated oil-containing wastewater on the laboratory scale. J. Environ. Manag. 2016, 173, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.; Rosales, E.; Sanromán, M.A.; Pazos, M. Preliminary testing and design of permeable bioreactive barrier for phenanthrene degradation by pseudomonas stutzeri CECT 930 immobilized in hydrogel matrices. J. Chem. Technol. Biotechnol. 2014, 90, 500–506. [Google Scholar] [CrossRef]
- Swisher, R.; Carroll, G.C. Fluorescein diacetate hydrolysis as an estimator of microbial biomass on coniferous needle surfaces. Microb. Ecol. 1980, 6, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Fontvieille, D.A.; Outaguerouine, A.; Thevenot, D.R. Fluorescein diacetate hydrolysis as a measure of microbial activity in aquatic systems: Application to activated sludges. Environ. Technol. 1992, 13, 531–540. [Google Scholar] [CrossRef]
- Jiang, S.; Huang, J.; Lu, H.; Liu, J.; Yan, C. Optimisation for assay of fluorescein diacetate hydrolytic activity as a sensitive tool to evaluate impacts of pollutants and nutrients on microbial activity in coastal sediments. Mar. Pollut. Bull. 2016, 110, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, X.; Dai, D.; Li, G. Porous biocarrier-enhanced biodegradation of crude oil contaminated soil. Int. Biodeterior. Biodegradation 2009, 63, 80–87. [Google Scholar] [CrossRef]
- Picioreanu, C.; van Loosdrecht, M.C.M.; Heijnen, J.J. Two-dimensional model of biofilm detachment caused by internal stress from liquid flow. Biotechnol. Bioeng. 2001, 72, 205–218. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Hupert-Kocurek, K.; Guzik, U.; Wojcieszyńska, D. Characterization of catechol 2,3-dioxygenase from Planococcus sp. strain S5 induced by high phenol concentration. Acta Biochem. Pol. 2012, 59, 345–351. [Google Scholar]
- Green, V.S.; Stott, D.E.; Diack, M. Assay for fluorescein diacetate hydrolytic activity: optimization for soil samples. Soil Biol. Biochem. 2006, 38, 693–701. [Google Scholar] [CrossRef]
- Fisher, K.A.; Huddersman, K.D.; Taylor, M.J. Comparison of micro- and mesoporous inorganic materials in the uptake and release of the drug model fluorescein and its analogues. Chem. - Eur. J. 2003, 9, 5873–5878. [Google Scholar] [CrossRef] [PubMed]
- Kasnavia, T.; Vu, D.; Sabatini, D.A. Fluorescent dye and media properties affecting sorption and tracer selection. Groundwater 1999, 37, 376–381. [Google Scholar] [CrossRef]
- Sabatini, D.A.; Austin, T.A. Characteristics of rhodamine WT and fluorescein as adsorbing ground-water tracers. Groundwater 1991, 29, 341–349. [Google Scholar] [CrossRef]
- Marchlewicz, A.; Domaradzka, D.; Guzik, U.; Wojcieszyńska, D. Bacillus thuringiensis b1 (2015b) is a gram-positive bacteria able to degrade naproxen and ibuprofen. Water, Air, Soil Pollut. 2016, 227, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Manohar, S.; Kim, C.K.; Karegoudar, T.B. Enhanced degradation of naphthalene by immobilization of pseudomonas sp. strain NGK1 in polyurethane foam. Appl. Microbiol. Biotechnol. 2001, 55, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Pradeep, T. Potential of silver nanoparticle-coated polyurethane foam as an antibacterial water filter. Biotechnol. Bioeng. 2005, 90, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phoenix, V.R.; Holmes, W.M. Magnetic resonance imaging of structure, diffusivity, and copper immobilization in a phototrophic biofilm. Appl. Environ. Microbiol. 2008, 74, 4934–4943. [Google Scholar] [CrossRef] [PubMed]
- Dikshit, P.K.; Moholkar, V.S. Kinetic analysis of dihydroxyacetone production from crude glycerol by immobilized cells of gluconobacter oxydans MTCC 904. Bioresour. Technol. 2016, 216, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, E.; Biggs, C.A. Mechanisms of bacillus cereus biofilm formation: An investigation of the physicochemical characteristics of cell surfaces and extracellular proteins. Appl. Microbiol. Biotechnol. 2010, 89, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Ingesson, H.; Zacchi, G.; Yang, B.; Esteghlalian, A.R.; Saddler, J.N. The effect of shaking regime on the rate and extent of enzymatic hydrolysis of cellulose. J. Biotechnol. 2001, 88, 177–182. [Google Scholar] [CrossRef]
- Sirisha, E.; Rajasekar, N.; Narasu, M.L. Isolation and optimization of lipase producing bacteria from oil contaminated soils. Adv. Biol. Res. 2010, 4, 249–252. [Google Scholar]
- Bisswanger, H. Enzyme assays. Perspect. Sci. 2014, 1, 41–55. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N. Fluorometric determination of lipase, acylase, alpha-, and gamma-chymotrypsin and inhibitors of these enzymes. Anal. Chem. 1964, 36, 409–412. [Google Scholar] [CrossRef]
- Frølund, B.; Griebe, T.; Nielsen, P.H. Enzymatic activity in the activated-sludge floc matrix. Appl. Microbiol. Biotechnol. 1995, 43, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, P.E.; Eriksen, T.; Jensen, B.K. Estimation of viable biomass in wastewater and activated sludge by determination of ATP, oxygen utilization rate and FDA hydrolysis. Water Res. 1992, 26, 1495–1501. [Google Scholar] [CrossRef]
- Pietikäinen, J.; Pettersson, M.; Bååth, E. Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol. Ecol. 2005, 52, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, G.; Duncan, H. Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol. Biochem. 2001, 33, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Sebastián, M.; Auguet, J.-C.; Restrepo-Ortiz, C.X.; Sala, M.M.; Marrasé, C.; Gasol, J.M. Deep ocean prokaryotic communities are remarkably malleable when facing long-term starvation. Environ. Microbiol. 2018, 20, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Cox, H.H.J.; Deshusses, M.A. Effect of starvation on the performance and re-acclimation of biotrickling filters for air pollution control. Environ. Sci. Technol. 2002, 36, 3069–3073. [Google Scholar] [CrossRef] [PubMed]
- Roszak, D.B.; Colwell, R.R. Survival strategies of bacteria in the natural environment. Microbiol. Rev. 1987, 51, 365–379. [Google Scholar] [PubMed]
- Ensign, J.C. Long-term starvation survival of rod and spherical cells of Arthrobacter crystallopoietes. J. Bacteriol. 1970, 103, 569–577. [Google Scholar] [PubMed]
- Gengenbacher, M.; Rao, S.P.S.; Pethe, K.; Dick, T. Nutrient-starved, non-replicating mycobacterium tuberculosis requires respiration, ATP synthase and isocitrate lyase for maintenance of ATP homeostasis and viability. Microbiology 2009, 156, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Voelker, U.; Voelker, A.; Maul, B.; Hecker, M.; Dufour, A.; Haldenwang, W.G. Separate mechanisms activate sigma B of Bacillus subtilis in response to environmental and metabolic stresses. J. Bacteriol. 1995, 177, 3771–3780. [Google Scholar] [CrossRef] [PubMed]
- Mielich-Süss, B.; Lopez, D. Molecular mechanisms involved in Bacillus subtilis biofilm formation. Environ. Microbiol. 2015, 17, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Sonenshein, A.L. Control of sporulation initiation in Bacillus subtilis. Curr. Opin. Microbiol. 2000, 3, 561–566. [Google Scholar] [CrossRef]
- Majed, R.; Faille, C.; Kallassy, M.; Gohar, M. Bacillus cereus biofilms—same, only different. Front. Microbiol. 2016, 7, 1054. [Google Scholar] [CrossRef] [PubMed]
- Correction: SinR controls enterotoxin expression in bacillus thuringiensis biofilms. PLoS ONE 2014, 9, e96707.
- Garcia-Ochoa, F.; Gomez, E.; Santos, V.E.; Merchuk, J.C. Oxygen uptake rate in microbial processes: An overview. Biochem. Eng. J. 2010, 49, 289–307. [Google Scholar] [CrossRef]
- Norsker, N.H.; Nielsen, P.H.; Hvitved-Jacobsen, T. Influence of oxygen on biofilm growth and potential sulfate reduction in gravity sewer biofilm. Water Sci. Tech. 1995, 31, 159–167. [Google Scholar] [CrossRef]
- Zhou, X.-H.; Qiu, Y.-Q.; Shi, H.-C.; Yu, T.; He, M.; Cai, Q. A new approach to quantify spatial distribution of biofilm kinetic parameters by in situ determination of oxygen uptake rate (our). Environ. Sci. Technol. 2009, 43, 757–763. [Google Scholar] [CrossRef] [PubMed]
- De Beer, D.; Stoodley, P.; Roe, F.; Lewandowski, Z. Effects of biofilm structures on oxygen distribution and mass transport. Biotechnol. Bioeng. 1994, 43, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wu, W.; Wang, J. Immobilization of activated sludge using improved polyvinyl alcohol (PVA) gel. J. Environ. Sci. 2007, 19, 1293–1297. [Google Scholar] [CrossRef]
- Bandaiphet, C.; Prasertsan, P. Effect of aeration and agitation rates and scale-up on oxygen transfer coefficient, kla in exopolysaccharide production from enterobacter cloacae WD7. Carbohydr. Polym. 2006, 66, 216–228. [Google Scholar] [CrossRef]
- Ganesh, R.; Balaji, G.; Ramanujam, R.A. Biodegradation of tannery wastewater using sequencing batch reactor—respirometric assessment. Bioresour. Technol. 2006, 97, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, S.; Schrader, J.; Sell, D. Oxygen uptake rate measurements to monitor the activity of terpene transforming fungi. Process Biochem. 2004, 39, 2221–2228. [Google Scholar] [CrossRef]
- Greń, I.; Wojcieszyńska, D.; Guzik, U.; Perkosz, M.; Hupert-Kocurek, K. Enhanced biotransformation of mononitrophenols by stenotrophomonas maltophilia kb2 in the presence of aromatic compounds of plant origin. World J. Microbiol. Biotechnol. 2009, 26, 289–295. [Google Scholar] [CrossRef]
- Battin, T.J. Assessment of fluorescein diacetate hydrolysis as a measure of total esterase activity in natural stream sediment biofilms. Sci. Total. Environ. 1997, 198, 51–60. [Google Scholar] [CrossRef]
- Subramanian, S.B.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Extracellular polymeric substances (EPS) producing bacterial strains of municipal wastewater sludge: Isolation, molecular identification, EPS characterization and performance for sludge settling and dewatering. Water Res. 2010, 44, 2253–2266. [Google Scholar] [CrossRef] [PubMed]
- Amon, R.M.W.; Benner, R. Photochemical and microbial consumption of dissolved organic carbon and dissolved oxygen in the Amazon River system. Geochim. Cosmochim. Acta 1996, 60, 1783–1792. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location of Application | Biofilm Dry Mass (g) | Fluorescein Concentration (µg/mL) | TEA (µg/g dry mass per h) | Mean | SD | CV (%) |
|---|---|---|---|---|---|---|
| Solution | 0.0082–0.0089 | 1.35–2.30 | 157–267 | 210 | 48 | 23 |
| Carrier | 0.0084–0.0086 | 2.02–2.40 | 238–283 | 262 | 18 | 7 |
| Agitation | Dry Biofilm Mass (g) | Fluorescein Concentration (µg/mL) | TEA (µg/g dry mass per h) | Mean | SD | CV (%) |
|---|---|---|---|---|---|---|
| With | 0.0079–0.0086 | 1.36–3.89 | 164–469 | 275 | 126 | 46 |
| Without | 0.0081–0.0089 | 1.92–2.32 | 226–273 | 249 | 21 | 8 |
| Incubation time (h) | Biofilm dry mass (g) | Dry EPS mass(g) | Fluorescein concentration (µg/ml) | TEA (µg/g dry mass per h) | Mean | SD | CV (%) |
|---|---|---|---|---|---|---|---|
| 24 | 0.0034–0.0049 | 0.0005–0.0007 | 1.21–1.66 | 326–383 | 360 | 24 | 7 |
| 48 | 0.0064–0.0078 | 0.0008–0.0010 | 1.63–2.45 | 255–325 | 287 | 28 | 10 |
| 72 | 0.0080–0.0091 | 0.0010–0.0013 | 1.43–1.50 | 166–180 | 170 | 7 | 4 |
| Incubation time (h) | TEA (µg/g dry mass per h) | OUR (µg O2/g dry mass per h) |
|---|---|---|
| 24 | 360 ± 24 | 176 ± 13 |
| 48 | 287 ± 28 | 120 ± 9 |
| 72 | 170 ± 7 | 70 ± 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dzionek, A.; Dzik, J.; Wojcieszyńska, D.; Guzik, U. Fluorescein Diacetate Hydrolysis Using the Whole Biofilm as a Sensitive Tool to Evaluate the Physiological State of Immobilized Bacterial Cells. Catalysts 2018, 8, 434. https://doi.org/10.3390/catal8100434
Dzionek A, Dzik J, Wojcieszyńska D, Guzik U. Fluorescein Diacetate Hydrolysis Using the Whole Biofilm as a Sensitive Tool to Evaluate the Physiological State of Immobilized Bacterial Cells. Catalysts. 2018; 8(10):434. https://doi.org/10.3390/catal8100434
Chicago/Turabian StyleDzionek, Anna, Jolanta Dzik, Danuta Wojcieszyńska, and Urszula Guzik. 2018. "Fluorescein Diacetate Hydrolysis Using the Whole Biofilm as a Sensitive Tool to Evaluate the Physiological State of Immobilized Bacterial Cells" Catalysts 8, no. 10: 434. https://doi.org/10.3390/catal8100434