The Synthetic Potential of Fungal Feruloyl Esterases: A Correlation with Current Classification Systems and Predicted Structural Properties

 ,
,  , ,
, ,  , , , and
, , , and 
Abstract
:1. Introduction
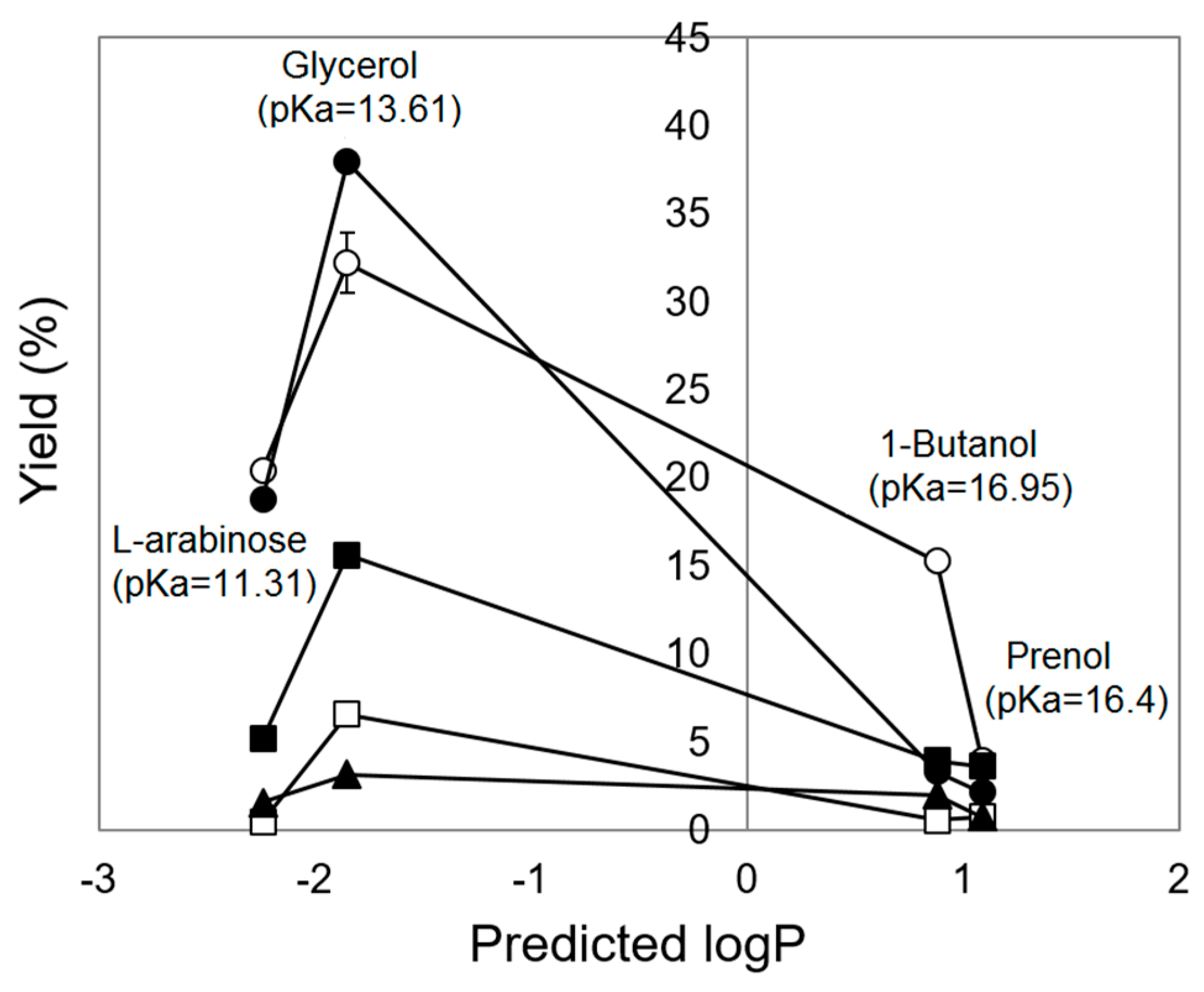
2. Results and Discussion
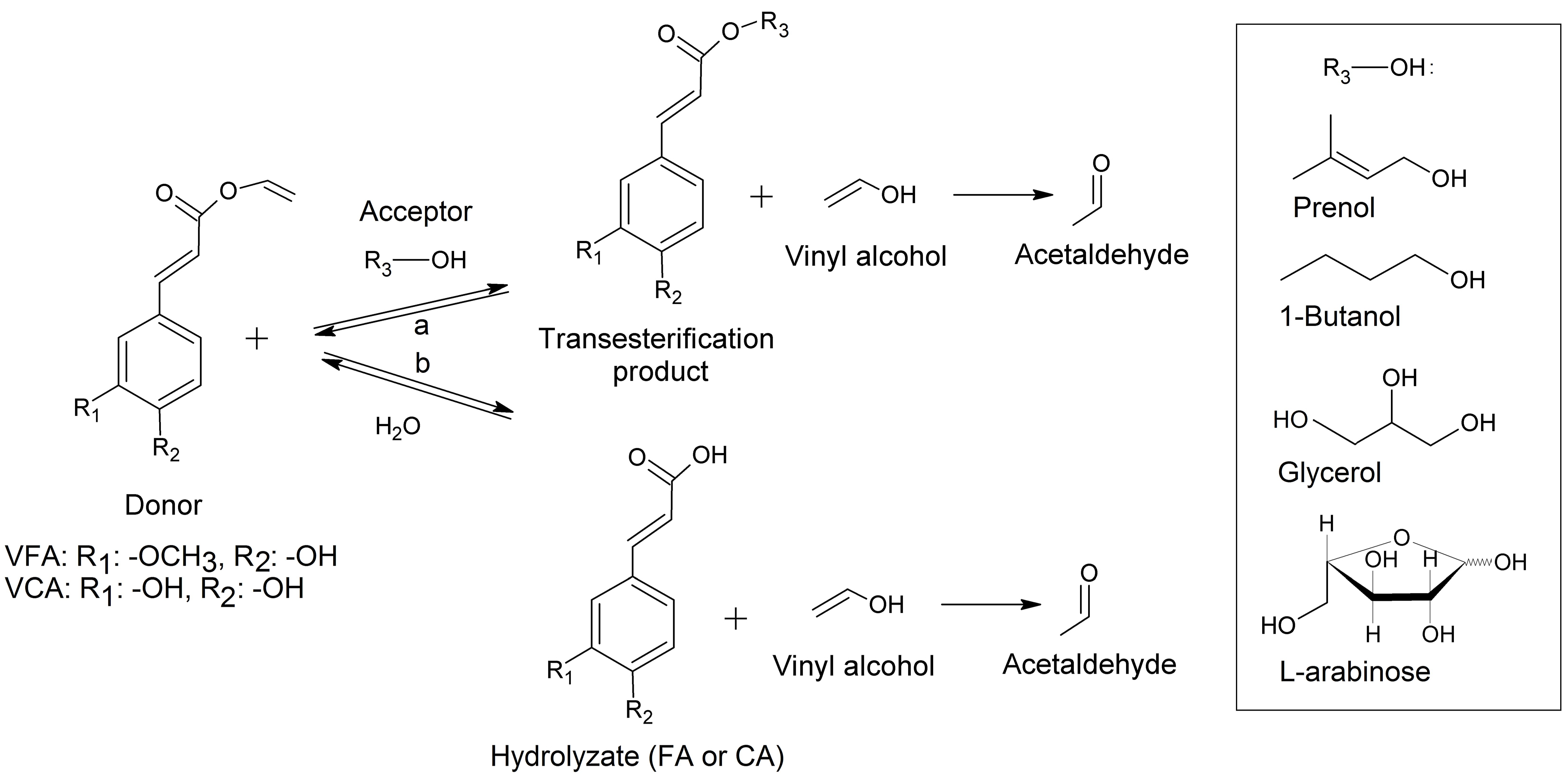
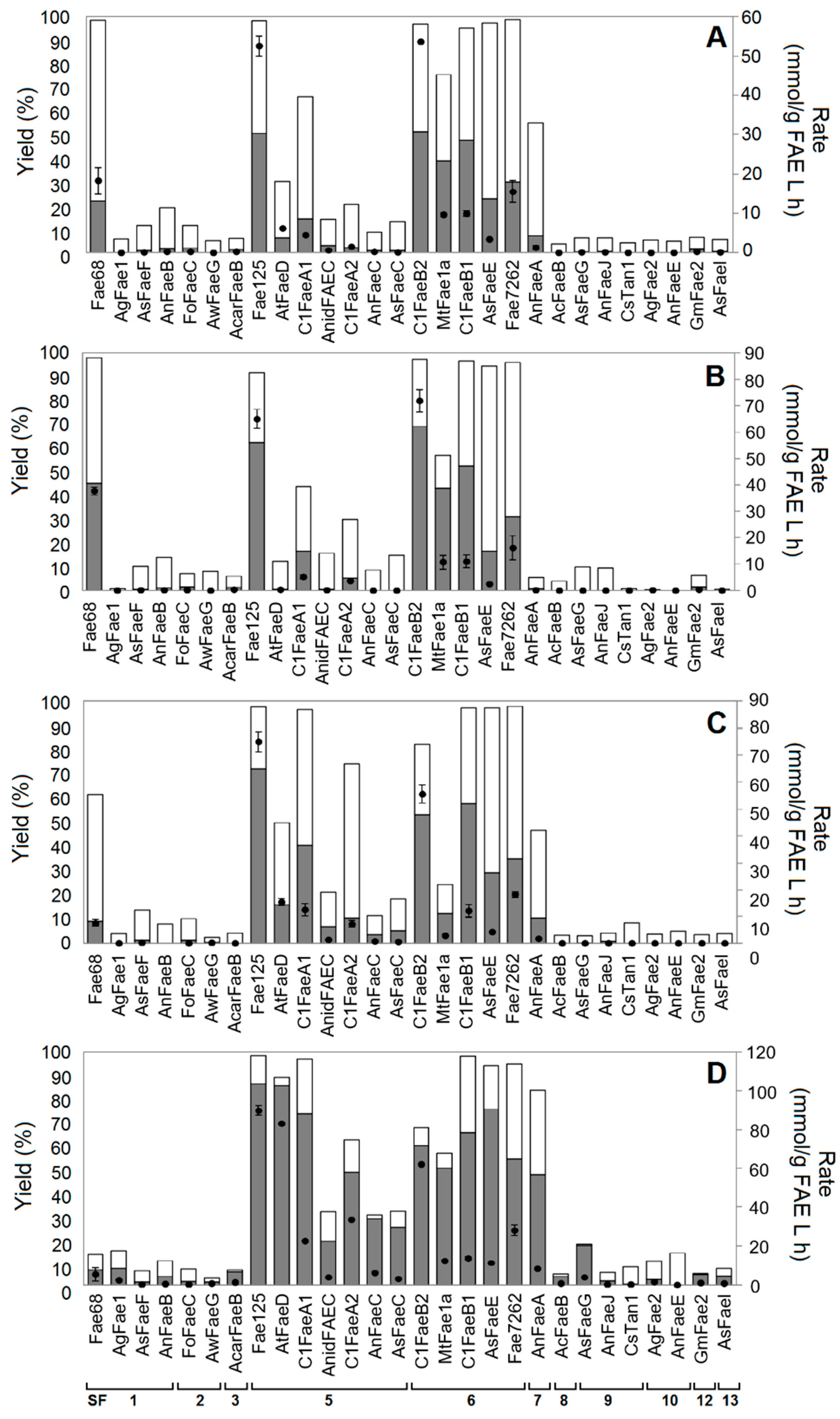
2.1. Synthesis of Aliphatic Esters
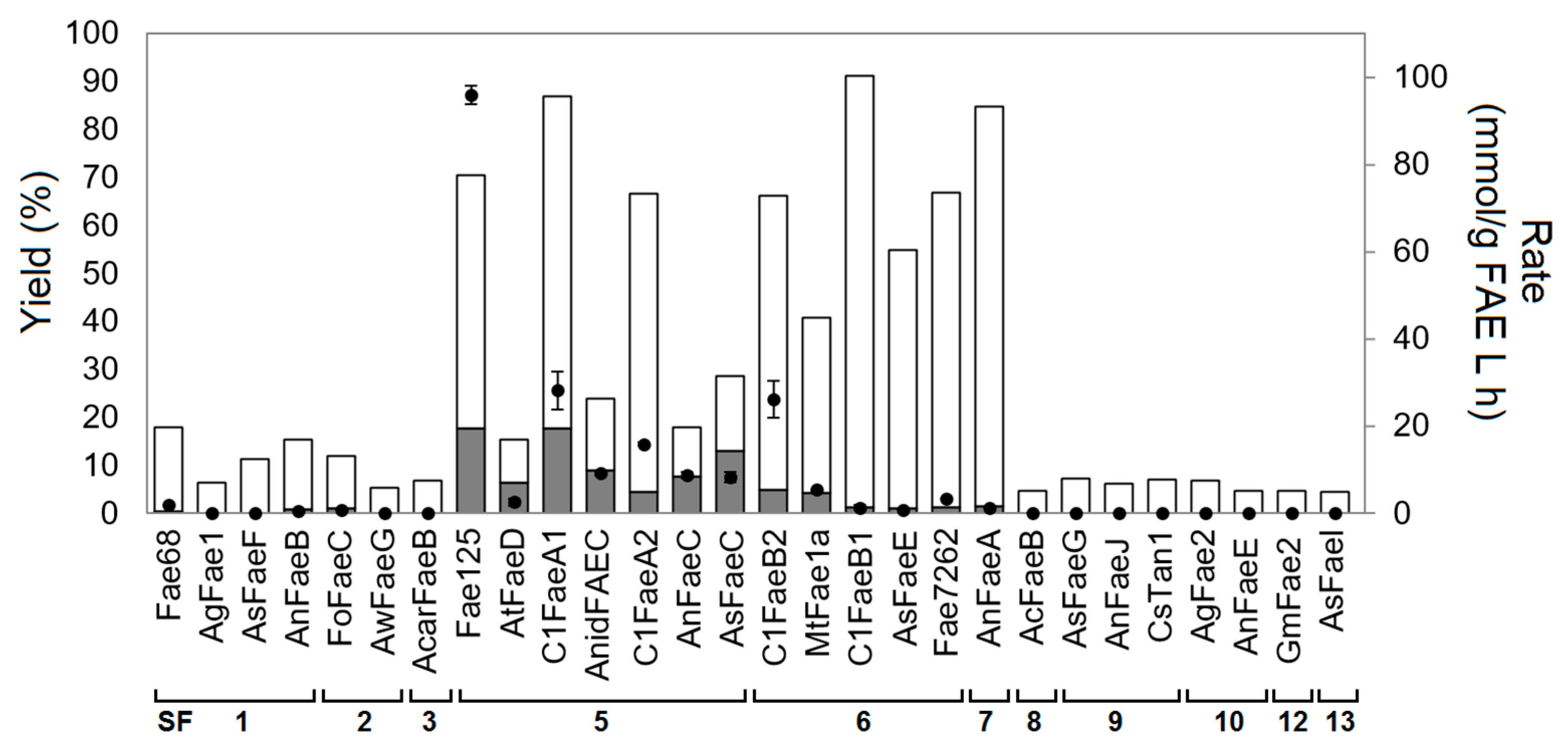
2.2. Synthesis of l-Arabinose Ferulate
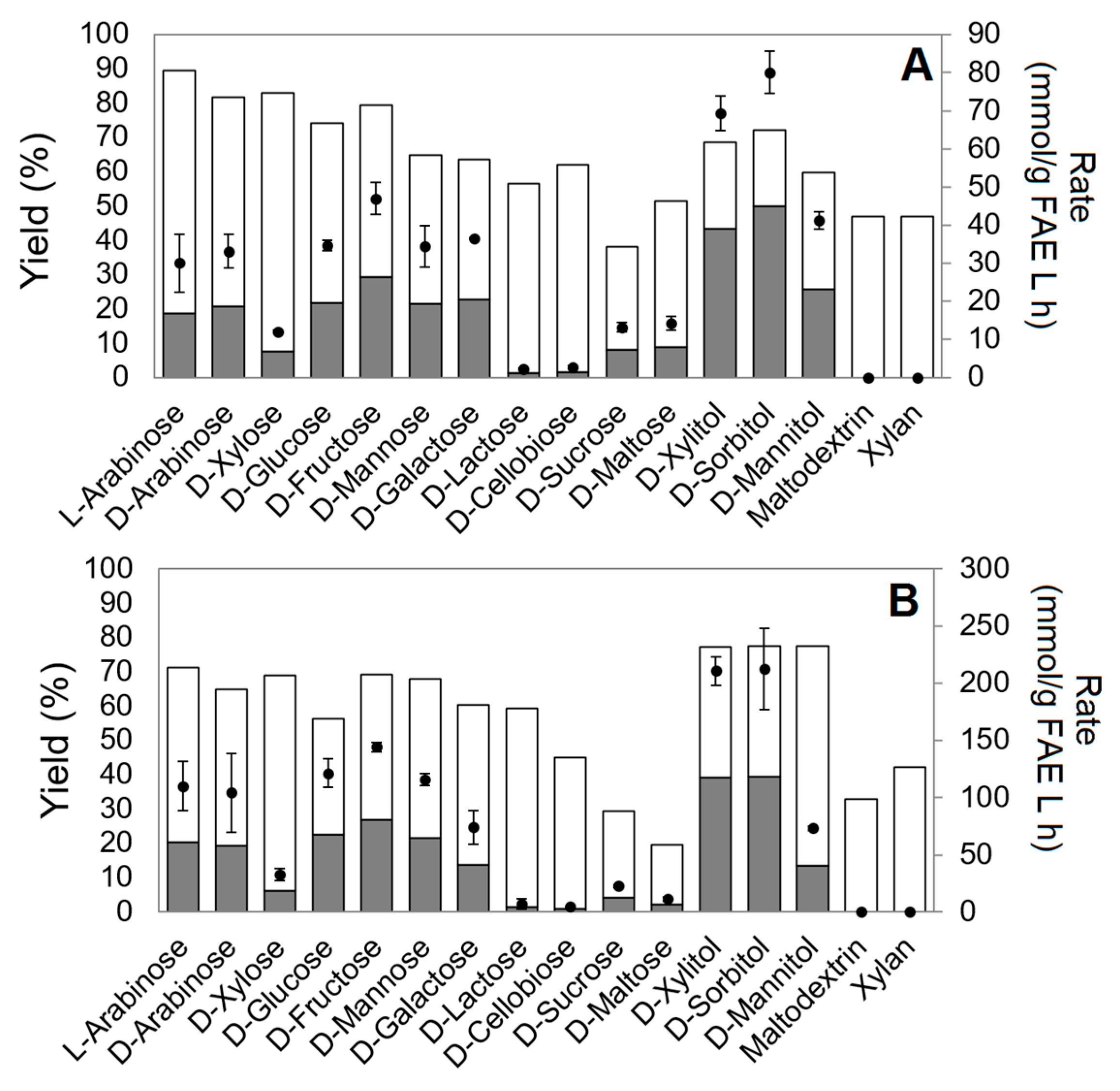
2.3. Sugar Ester Synthesis by Two SF5 FAEs
2.4. Correlation with Phylogenetic Classification
2.5. Correlation with ABCD Classification
2.6. Factors Affecting Transesterification
3. Materials and Methods
3.1. Production of Enzymes in Myceliophthora thermophila C1
3.2. Production of Enzymes in Pichia pastoris
3.3. Enzymatic Assays
3.4. Transesterification Reactions
3.5. Quantitative Analysis of Hydroxycinnamates
3.6. Bioinformatic Tools
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Giuliani, S.; Piana, C.; Setti, L.; Hochkoeppler, A.; Pifferi, P.G.; Williamson, G.; Faulds, C.B. Synthesis of pentylferulate by feruloyl esterase from Aspergillus niger using water-in-oil microemulsions. Biotechnol. Lett. 2001, 23, 325–330. [Google Scholar] [CrossRef]
- Topakas, E.; Stamatis, H.; Biely, P.; Kekos, D.; Macris, B.J.; Christakopoulos, P. Purification and characterization of a feruloyl esterase from Fusarium oxysporum catalyzing esterification of phenolic acids in ternary water-organic solvent mixtures. J. Biotechnol. 2003, 102, 33–44. [Google Scholar] [CrossRef]
- Vafiadi, C.; Topakas, E.; Wong, K.K.Y.; Suckling, I.D.; Christakopoulos, P. Mapping the hydrolytic and synthetic selectivity of a type C feruloyl esterase (StFaeC) from Sporotrichum thermophile using alkyl ferulates. Tetrahedron Asymmetry 2005, 16, 373–379. [Google Scholar] [CrossRef]
- Tsuchiyama, M.; Sakamoto, T.; Fujita, T.; Murata, S.; Kawasaki, H. Esterification of ferulic acid with polyols using a ferulic acid esterase from Aspergillus niger. Biochim. Biophys. Acta 2006, 7, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiyama, M.; Sakamoto, T.; Tanimori, S.; Murata, S.; Kawasaki, H. Enzymatic synthesis of hydroxycinnamic acid glycerol esters using type A feruloyl esterase from Aspergillus niger. Biosci. Biotechnol. Biochem. 2007, 71, 2606–2609. [Google Scholar] [CrossRef] [PubMed]
- Vafiadi, C.; Topakas, E.; Christakopoulos, P. Preparation of multipurpose cross-linked enzyme aggregates and their application to production of alkyl ferulates. J. Mol. Catal. B Enzym. 2008, 54, 35–41. [Google Scholar] [CrossRef]
- Vafiadi, C.; Topakas, E.; Alissandratos, A.; Faulds, C.B.; Christakopoulos, P. Enzymatic synthesis of butyl hydroxycinnamates and their inhibitory effect on LDL-oxidation. J. Biotechnol. 2008, 133, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Couto, J.; Karboune, S.; Mathew, R. Regioselective synthesis of feruloylated glycosides using the feruloyl esterase expressed in selected commercial multi-enzymatic preparations as biocatalysts. Biocatal. Biotransform. 2010, 28, 235–244. [Google Scholar] [CrossRef]
- Couto, J.; St-Louis, R.; Karboune, S. Optimization of feruloyl esterase-catalyzed synthesis of feruloylated oligosaccharides by response face methodology. J. Mol. Catal. B Enzym. 2011, 73, 53–62. [Google Scholar] [CrossRef]
- Kikugawa, M.; Tsuchiyama, M.; Kai, K.; Sakamoto, T. Synthesis of highly water-soluble feruloyl diglycerols by esterification of an Aspergillus niger feruloyl esterase. Appl. Microbiol. Biotechnol. 2012, 95, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, I.; Leonov, L.; Jütten, P.; Cerullo, G.; Faraco, V.; Papadopoulou, A.; Kletsas, D.; Ralli, M.; Rova, U.; Christakopoulos, P. Optimized synthesis of novel prenyl ferulate performed by feruloyl esterases from Myceliophthora thermophila in microemulsions. Appl. Microbiol. Biotechnol. 2017, 101, 3213–3226. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, I.; Papadopoulou, A.; Iancu, L.; Cerullo, G.; Ralli, M.; Jütten, P.; Piechot, A.; Faraco, V.; Kletsas, D.; Rova, U.; et al. Optimization of enzymatic synthesis of l-arabinose ferulate catalyzed by feruloyl esterases from Myceliophthora thermophila in detergentless microemulsions and assessment of its antioxidant and cytotoxicity activities. Process Biochem. 2018, 101, 3213–3226. [Google Scholar] [CrossRef]
- Faulds, C.B.; Perez-Boada, M.; Martinez, A.T. Influence of organic co-solvents on the activity of substrate specificity of feruloyl esterase. Bioresour. Technol. 2011, 102, 4962–4967. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, I.; Varriale, S.; Topakas, E.; Rova, U.; Christakopoulos, P.; Faraco, V. Enzymatic synthesis of bioactive compounds with high potential for cosmeceutical application. Appl. Microbiol. Biotechnol. 2016, 100, 6519–6543. [Google Scholar] [CrossRef] [PubMed]
- Khmelnitsky, Y.L.; Hilhorst, R.; Verger, C. Detergentless microemulsions as media for enzymatic reactions. FEBS J. 1988, 176, 265–271. [Google Scholar] [CrossRef]
- Topakas, E.; Vafiadi, C.; Christakopoulos, P. Microbial production, characterization and applications of feruloyl esterases. Process Biochem. 2007, 42, 497–509. [Google Scholar] [CrossRef]
- Katsimpouras, C.; Antonopoulou, I.; Christakopoulos, P.; Topakas, E. Role and applications of feruloyl esterases in biomass bioconversion. In Microbial Enzymes in Bioconversion of Biomass, 1st ed.; Gupta, V., Ed.; Springer: Cham, Switzerland, 2016; pp. 79–123. [Google Scholar]
- Dilokpimol, A.; Mäkelä, M.R.; Aguilar-Pontes, M.V.; Benoit-Gelber, I.; Hildén, K.S.; de Vries, R.P. Diversity of fungal feruloyl esterases: Updated phylogenetic classification, properties and industrial applications. Biotechnol. Biofuels 2016, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Crepin, V.F.; Faulds, C.B.; Connerton, I.F. Functional classification of the microbial feruloyl esterases. Appl. Microbiol. Biotechnol. 2004, 63, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Benoit, I.; Danchin, E.G.J.; Blechrodt, R.-J.; de Vries, R.P. Biotechnological applications and potential of fungal feruloyl esterases based on prevalence, classification and biochemical diversity. Biotechnol. Lett. 2008, 30, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Dilokpimol, A.; Mäkelä, M.R.; Varriale, S.; Zhou, M.; Cerullo, G.; Gidijala, L.; Brás, J.L.A.; Jütten, P.; Piechot, A.; Verhaert, R.; et al. Fungal feruloyl esterases: Functional validation of genome mining based enzyme discovery including uncharacterized subfamilies. New Biotechnol. 2018, 41, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Vafiadi, C.; Topakas, E.; Nahmias, V.R.; Faulds, C.B.; Christakopoulos, P. Feruloyl esterase-catalysed synthesis of glycerol sinapate using ionic liquids mixtures. J. Biotechnol. 2009, 139, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Vafiadi, C.; Topakas, E.; Bakx, E.J.; Schols, H.A.; Christakopoulos, P. Structural characterization of ESI-MS of feruloylated arabino-oligosaccharides synthesized by chemoenzymatic esterification. Molecules 2007, 12, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Vafiadi, C.; Topakas, E.; Christakopoulos, P. Regioselective esterase-catalyzed feruloylation of l-arabinobiose. Carbohydr. Res. 2006, 241, 1992–1997. [Google Scholar] [CrossRef] [PubMed]
- Kelle, S.; Nieter, A.; Krings, U.; Zelena, K.; Linke, D.; Berger, R.G. Heterologous production of a feruloyl esterase from Pleurotus sapidus synthesizing feruloyl-saccharide esters. Biotechnol. Appl. Biochem. 2016, 63, 852–862. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.P.; van Kuyk, P.A.; Kester, H.C.M.; Visser, J. The Aspergillus niger faeB gene induced in the presence of aromatic compounds. Biochem. J. 2002, 363, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Moukouli, M.; Topakas, E.; Christakopoulos, P. Cloning, characterization and functional expression of an alkalitolerant type C feruloyl esterase from Fusarium oxysporum. Appl. Microbiol. Biotechnol. 2008, 79, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, M.R.; Dilokpimol, A.; Koskela, S.; Kuuskeri, J.; de Vries, R.P.; Hilden, K. Characterization of a feruloyl esterase from Aspergillus terreus facilitates the division of fungal enzymes from Carbohydrate Esterase family 1 of the CAZy database. Microb. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kühnel, S.; Pouvreau, L.; Appeldoorn, M.M.; Hinz, S.W.A.; Schols, H.A.; Gruppen, H. The ferulic acid esterases from Chrysosporium lucknowense C1: Purification, characterization and their potential application in biorefinery. Enzyme Microb. Technol. 2012, 50, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Dilokpimol, A.; Mäkelä, M.R.; Mansouri, S.; Belova, O.; Waterstraat, M.; Bunzel, M.; de Vries, R.P.; Hilden, K. Expanding the feruloyl esterase gene family of Aspergillus niger by characterization of feruloyl esterase, FaeC. New Biotechnol. 2017, 37, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Topakas, E.; Moukouli, M.; Dimarogona, M.; Christakopoulos, P. Expression, characterization and structural modelling of a feruloyl esterase from the thermophilic fungus Myceliophthora thermophila. Appl. Microbiol. Biotechnol. 2012, 94, 399–411. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.P.; Michelsen, B.; Roulsen, C.H.; Kroon, P.A.; van den Heuvel, R.H.; Faulds, C.B.; Williamson, G.; van den Hombergh, J.P.; Visser, J. The faeA genes from Aspergillus niger and Aspergillus tubingensis encode ferulic acid esterases involved in degradation of complex cell wall polysaccharides. Appl. Environ. Microbiol. 1997, 63, 4638–4644. [Google Scholar] [PubMed]
- Suzuki, K.; Hori, A.; Kawamoto, K.; Thangudu, R.R.; Ishida, T.; Igarashi, K.; Samejima, M.; Yamada, C.; Arakawa, T.; Wakagi, T.; et al. Crystal structure of a feruloyl esterase belonging to the tannase family: A disulfide bond near a catalytic triad. Proteins 2014, 82, 2857–2867. [Google Scholar] [CrossRef] [PubMed]
- Komiya, D.; Hori, A.; Ishida, T.; Igarashi, K.; Samejima, M.; Koseki, T.; Fushinobu, S. Crystal structure and substrate specificity modification of acetyl xylan esterase from Aspergillus luchuensis. Appl. Environ. Microbiol. 2017, 83, e01251-17. [Google Scholar] [CrossRef] [PubMed]
- Visser, H.; Joosten, V.; Punt, P.J.; Gusakov, A.V.; Olson, P.T.; Joosten, R.; Bartels, J.; Visser, J.; Sinitsyn, A.P.; Emalfarb, M.A.; et al. Development of a mature fungal technology and production platform for industrial enzymes based on a Myceliophthora thermophila isolate, previously known as Chrysosporium lucknowense C1. Ind. Biotechnol. 2011, 7, 214–223. [Google Scholar] [CrossRef]
- Verdoes, J.C.; Punt, P.J.; Burlingame, R.P.; Pynnonen, C.M.; Olson, P.T.; Wery, J. New Fungal Production System. Patent No. WO2010107303 A2, 23 September 2010. [Google Scholar]
- Petersen, T.N.; Brunak, S.; von Hejine, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy Server. In The Proteomics Protocols Handbook, 1st ed.; Walker, J.M., Ed.; Humana Press: New York, NY, USA, 2005; pp. 571–607. [Google Scholar]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.-B.G.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins 2009, 77, 114–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Hermoso, J.A.; Aparicia, J.S.; Molina, R.; Juge, N.; Gonzalez, R.; Faulds, C.B. The crystal structure of feruloyl esterase A from Aspergillus niger suggests evolutive functional convergence in feruloyl esterase family. J. Mol. Biol. 2004, 338, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Vriend, G. YASARA View—Molecular graphic for all devices- from smartphones to workstations. Bioinformatics 2014, 9, 114–122. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Sample | Origin | Expression | Calculated/Observed MW (kDa) | Calculated pI | N-Glycosylation | O-Glycosylation | FAE Content (% g FAE g−1 Protein) | Nucleophilic Elbow | SF a | Accession Number |
|---|---|---|---|---|---|---|---|---|---|---|
| Fungal FAEs | ||||||||||
| Fae68 | Talaromyces wortmannii | Myceliophthora thermophila C1 | 58.7/58.8 | 4.96 | 10 | 11 | 10–15 | GCSTG | 1 | MF362596 |
| AgFae1 | Aspergillus glaucus | Pichia pastoris | 55.8/75 | 5.49 | 7 | 1 | 31.3 | GCSTG | 1 | OJJ86166 |
| AsFaeF | Aspergillus sydowii | Pichia pastoris | 55.4/75 | 4.89 | 6 | 1 | 67.5 | GCSTG | 1 | jgi|Aspsy1|293049 |
| AnFaeB | Aspergillus niger | Pichia pastoris | 55.6/74 | 4.99 | 12 | 3 | 60.3 | GCSTG | 1 | Q8WZI8 |
| FoFaeC | Fusarium oxysporum | Pichia pastoris | 62.0/62 | 6.39 | 6 | 3 | 86.7 | GCSTG | 2 | jgi|Fusox1|5438 |
| AwFaeG | Aspergillus wentii | Pichia pastoris | 58.2/58 | 5.03 | 7 | 11 | 17.2 | GCSTG | 2 | jgi|Aspwe1|156253 |
| AcarFaeB | Aspergillus carbonarius | Pichia pastoris | 56.0/58 | 5.02 | 5 | 2 | n/q | GCSFG | 3 | jgi|Aspca3|176503 |
| Fae125 b | Talaromyces wortmannii | Myceliophthora thermophila C1 | 33.9/40 | 5.64 | 0 | 22 | 10 | GWSYG | 5 | MF362595 |
| AtFaeD | Aspergillus terreus | Pichia pastoris | 26.5/43 | 4.22 | 0 | 1 | 10.7 | GWSWG | 5 | XP_001215822 |
| C1FaeA1 | Myceliophthora thermophila C1 | Myceliophthora thermophila C1 | 27.2/29 | 6.15 | 0 | 2 | 33.7 | GWSYG | 5 | JF826027 |
| AnidFAEC | Aspergillus nidulans | Pichia pastoris | 25.8/30 | 4.47 | 0 | 2 | 52.9 | GFSWG | 5 | EAA62427 |
| C1FaeA2 | Myceliophthora thermophila C1 | Myceliophthora thermophila C1 | 29.1/36 | 5.05 | 1 | 8 | 15.0 | GFSYG | 5 | JF826028 |
| AnFaeC | Aspergillus niger | Pichia pastoris | 28.2/30 | 4.44 | 1 | 1 | 47.3 | GFSWG | 5 | An12g02550 |
| AsFaeC | Aspergillus sydowii | Pichia pastoris | 25.9/30 | 4.44 | 0 | 0 | 83.2 | GFSWG | 5 | jgi|Aspsy1|154482 |
| C1FaeB2 c | Myceliophthora thermophila C1 | Myceliophthora thermophila C1 | 28.4/33 | 4.57 | 2 | 1 | 10.0 | GFSSG | 6 | JF826029 |
| MtFae1a c | Myceliophthora thermophila ATCC 42464 | Pichia pastoris | 28.4/39 | 4.57 | 2 | 1 | 42.2 | GFSSG | 6 | AEO62008 |
| C1FaeB1 | Myceliophthora thermophila C1 | Myceliophthora thermophila C1 | 28.3/29 | 5.74 | 1 | 2 | 50.0 | GGSSG | 6 | API68922 |
| AsFaeE | Aspergillus sydowii | Pichia pastoris | 29.5/32 | 4.24 | 2 | 1 | 70.0 | GSSSG | 6 | jgi|Aspsy1|1158585 |
| Fae7262 b | Talaromyces wortmannii | Myceliophthora thermophila C1 | 35.8/43 | 5.23 | 2 | 23 | 15–25 | GSSSG | 6 | MF362597 |
| AnFaeA | Aspergillus niger | Pichia pastoris | 28.4/36 | 4.21 | 1 | 0 | 89.6 | GHSLG | 7 | CAA70510 |
| AcFaeB | Aspergillus clavatus | Pichia pastoris | 39.1/40 | 6.23 | 4 | 1 | n/q | GHSFG | 8 | jgi|Aspcl1|3045 |
| AsFaeG | Aspergillus sydowii | Pichia pastoris | 57.6/60 | 4.99 | 7 | 1 | 45.0 | GCSTG | 9 | jgi|Aspsy1|41271 |
| AnFaeJ | Aspergillus niger | Pichia pastoris | 58.3/100 | 4.88 | 9 | 2 | 61.2 | GCSTG | 9 | An15g05280 |
| CsTan1 | Ceriporiopsis subvermispora | Pichia pastoris | 57.6/90 | 4.67 | 7 | 4 | 48.5 | GCSTG | 9 | jgi|Cersu1|89153 |
| AgFae2 | Aspergillus glaucus | Pichia pastoris | 53.2/75 | 5.55 | 7 | 5 | 16.6 | GCSTG | 10 | OJJ88972 |
| AnFaeE | Aspergillus niger | Pichia pastoris | 55.0/88 | 4.11 | 8 | 14 | 71.2 | GCSTG | 10 | An11g01220 |
| GmFae2 | Galerina marginata | Pichia pastoris | 57.0/59 | 6.49 | 5 | 4 | n/q | GESAG | 12 | jgi|Galma1|254175 |
| AsFaeI | Aspergillus sydowii | Pichia pastoris | 59.4/55 | 4.90 | 5 | 5 | n/q | GESAG | 13 | jgi|Aspsy1|160668 |
| Enzyme Sample | Type | Relative Hydrolytic Activity (%) | SF a | Reference | ||||
|---|---|---|---|---|---|---|---|---|
| MFA | MCA | MSA | MpCA | pNP-Fe | ||||
| Fungal FAEs | ||||||||
| Fae68 | B | 4.10 | 3.39 | 0 | 5.84 | n/d | 1 | This work |
| AgFae1 | C | 0.031 | 0.047 | 0.005 | 0.020 | 0.025 | 1 | This work |
| AsFaeF | B | 60.12 | 65.88 | 0 | 100 | 1.552 | 1 | [21] |
| AnFaeB | B | 1.131 | 1.486 | 0 | 1.930 | 0.00074 | 1 | [21,26] |
| FoFaeC | C | 0.247 | 0.389 | 0.013 | 0.457 | n/d | 2 | [21,27] This work |
| AwFaeG | n/a | 0.00012 | 0 | 0 | 0 | 0.0016 | 2 | [21] |
| AcarFaeB | n/a | 0 | 0.0024 | 0 | 0 | 0.0006 | 3 | [21] |
| Fae125 | A | 0.059 | 0.024 | 0.047 | 0.023 | n/d | 5 | This work |
| AtFaeD | C | 0.00022 | 0.00007 | 0.00022 | 0.00011 | 0.0172 | 5 | [28] |
| C1FaeA1 | A | 0.127 | 0.057 | 0.114 | 0.059 | n/d | 5 | [29] |
| AnidFAEC | C or D | 2.53 | 1.07 | 1.18 | 2.48 | 0.555 | 5 | [21] |
| C1FaeA2 | A/B | 0.076 | 0.045 | 0.031 | 0.064 | n/d | 5 | [29] |
| AnFaeC | C | 0.998 | 0.466 | 0.555 | 0.887 | n/d | 5 | [30] |
| AsFaeC | C or D | 3.84 | 1.71 | 1.51 | 3.50 | 0.555 | 5 | [21] |
| C1FaeB2 c | B | 0.616 | 0.458 | 0.096 | 0.794 | n/d | 6 | [29] |
| MtFae1a c | B | 0.0010 | 0.0056 | 0 | 0 | 0.060 | 6 | [21,31] |
| C1FaeB1 | B | 0.596 | 1.50 | 0 | 0.981 | n/d | 6 | [28] |
| AsFaeE | C or D | 7.03 | 3.75 | 4.19 | 6.70 | 1.44 | 6 | [21] |
| Fae7262 | B | 0.377 | 0.290 | 0 | 0.576 | n/d | 6 | This work |
| AnFaeA | A | 1.708 | 0 | 1.375 | 0 | 0.0015 | 7 | [21,32] |
| AcFaeB b | n/a | ~0 | ~0 | ~0 | ~0 | ~0 | 8 | [21] |
| AsFaeG d | n/a | 0 | 0 | 0 | 0 | 0 | 9 | [21] |
| AnFaeJ d | n/a | 0 | 0 | 1.52 | 0.149 | 0.0023 | 9 | [21] |
| CsTan1 d | n/a | 0 | 0 | 0 | 0 | 0 | 9 | [21] |
| AgFae2 | C | 0.0019 | 0.0007 | 0.0019 | 0.0015 | 0.0077 | 10 | This work |
| AnFaeE | C | 0.011 | 0.040 | 0.009 | 0.001 | n/d | 10 | [21] |
| GmFae2 | n/a | 0 | 0 | 0 | 0 | 0.0007 | 12 | [21] |
| AsFaeI | B | 58.85 | 64.22 | 0 | 98.56 | 1.353 | 13 | [21] |
| SF a | Query | Subject | Sequence Alignment Results (pBLAST) | Correlation between Primary Sequence Identity and Transesterification Rate b | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Query Cover (%) | Identities (%) | Positives (%) | Gaps (%) | E-Value | PFA | BFA | GFA | AFA | |||
| 1 | Fae68 | AgFae1 | 96 | 57 | 74 | 3 | 0.0 | 0.9956 | 0.9933 | 0.9955 | 0.9833 |
| AsFaeF | 96 | 54 | 71 | 3 | 0.0 | ||||||
| AnFaeB | 97 | 52 | 71 | 3 | 0.0 | ||||||
| 5 | Fae125 | ATEG_Fae | 77 | 57 | 72 | 0 | 6 × 10−109 | 0.9774 | 0.9866 | 0.8199 | 0.9170 |
| C1FaeA1 | 82 | 51 | 67 | 1 | 2 × 10−90 | ||||||
| AnidFAEC | 77 | 46 | 63 | 6 | 7 × 10−79 | ||||||
| C1FaeA2 | 78 | 44 | 61 | 7 | 5 × 10−75 | ||||||
| AnFaeC | 77 | 44 | 61 | 6 | 6 × 10−75 | ||||||
| AsFaeC | 77 | 43 | 58 | 4 | 3 × 10−73 | ||||||
| 6 | C1FaeB2 | C1FaeB1 | 100 | 68 | 81 | 0 | 6 × 10−144 | 0.9044 | 0.7886 | 0.8741 | 0.8699 |
| AsFaeE | 100 | 48 | 68 | 0 | 4 × 10−99 | ||||||
| Fae7262 | 99 | 47 | 64 | 0 | 1 × 10−93 | ||||||
| 9 | AsFaeG | AnFaeJ | 99 | 46 | 64 | 2 | 7 × 10−173 | 0.6162 | 0.2161 | 0.9925 | - |
| CsTan1 | 93 | 37 | 56 | 4 | 1 × 10−100 | ||||||
| Enzyme | Fae68 | AgFae1 | AsFaeF | AnFaeB | FoFaeC | AwFaeG | C1FaeB2 | C1FaeB1 | Fae7262 | AsFaeE | AnFaeA a |
|---|---|---|---|---|---|---|---|---|---|---|---|
| SF | 1 | 1 | 1 | 1 | 2 | 2 | 6 | 6 | 6 | 6 | 7 |
| Protein size (kDa) | 57.7 | 55.5 | 55.4 | 55.5 | 56.1 | 55.9 | 30.5 | 30.2 | 31.2 | 30.2 | 28.4 |
| Protein volume (Å3) | 62,063 | 60,284 | 59,722 | 60,410 | 60,107 | 59,908 | 32,825 | 32,506 | 33,286 | 32,281 | 301,120 |
| Catalytic triad (Ser-His-Asp) | 214-469-429 | 187-440-400 | 187-439-399 | 185-437-397 | 201-452-412 | 190-442-402 | 136-275-219 | 137-277-221 | 136-276-220 | 137-277-221 | 133-247-194 |
| Molecular Surface (Å2) | |||||||||||
| Protein | |||||||||||
| Lipophilic (%) | 33.5 | 30.6 | 32.9 | 31.2 | 30.6 | 33.6 | 35.6 | 38.8 | 32.3 | 35.3 | 26.8 |
| Non-polar (%) | 40.0 | 36.9 | 39.8 | 38.0 | 38.3 | 40.6 | 44.2 | 47.0 | 47.8 | 43.6 | 32.3 |
| Neutral (%) | 42.1 | 41.9 | 36.4 | 42.9 | 35.4 | 39.1 | 44.2 | 37.9 | 54.2 | 38.3 | 48.5 |
| Acidic (%) | 11.4 | 11.6 | 14.6 | 13.1 | 12.7 | 12.6 | 11.5 | 8.6 | 6.0 | 16.1 | 16.4 |
| Basic (%) | 12.7 | 15.5 | 15.5 | 14.3 | 20.9 | 14.5 | 8.3 | 14.3 | 7.2 | 9.9 | 8.1 |
| Distance < 15 Å around the catalytic serine | |||||||||||
| Lipophilic (%) | 38.9 | 30.4 | 36.2 | 26.4 | 32.7 | 29.2 | 45.4 | 43.7 | 33.5 | 43.8 | 30.2 |
| Non-polar (%) | 48.2 | 38.8 | 43.6 | 38.9 | 43.7 | 38.8 | 56.9 | 53.4 | 41.5 | 51.5 | 37.3 |
| Neutral (%) | 42.1 | 49.6 | 43.5 | 49.3 | 43.3 | 46.4 | 47.0 | 38.2 | 52.9 | 36.2 | 44.5 |
| Acidic (%) | 8.2 | 7.2 | 11.4 | 14.2 | 9.7 | 10.3 | 5.4 | 3.8 | 4.7 | 6.3 | 12.7 |
| Basic (%) | 10.0 | 12.2 | 8.2 | 9.5 | 13.4 | 13.4 | 6.7 | 13.6 | 8.4 | 13.2 | 11.9 |
| Lid b | |||||||||||
| Lipophilic (%) | 39.2 | 33.5 | 38.1 | 37.7 | 36.9 | 37.0 | - | - | - | - | 16.4 |
| Non-polar (%) | 44.7 | 38.1 | 44.4 | 44.7 | 45.5 | 44.1 | 27.6 | ||||
| Neutral (%) | 39.9 | 43.7 | 35.9 | 38.0 | 37.8 | 41.5 | 41.3 | ||||
| Acidic (%) | 11.9 | 7.8 | 10.8 | 14.1 | 12.2 | 8.6 | 24.1 | ||||
| Basic (%) | 8.6 | 15.6 | 14.6 | 9.8 | 12.7 | 12.5 | 1.6 | ||||
| Receptor | SF a | Mean Binding Energy of Cluster (kcal/mol) | Highest Binding Energy in Cluster (kcal/mol) | Distance b | Genetic Runs in Cluster c | Residue(s) in Vicinity with Vinyl Moiety in Cluster |
|---|---|---|---|---|---|---|
| Fae68 | 1 | 6.0457 ± 0.3754 | 6.5580 | 2.823 | 3 | Ile358 |
| AgFae1 | 4.6875 ± 0.2963 | 5.3330 | 3.814 | 15 | Gly108 | |
| AsFaeF | 5.5160 ± 0.1797 | 5.7610 | 2.842 | 3 | Gly108 | |
| AnFaeB | 5.1805 ± 0.1125 | 5.2930 | 2.745 | 2 | Gly106 | |
| FoFaeC | 2 | 5.6489 ± 0.3975 | 6.4670 | 4.188 | 8 | Gly122 |
| AwFaeG | 5.6604 ± 0.4224 | 6.3690 | 2.953 | 12 | Gly111 | |
| C1FaeB1 | 6 | 4.9831 ± 0.2644 | 5.3670 | 2.955 | 8 | Pro57 |
| 4.7887 ± 0.2200 | 5.0730 | 3.968 | 3 | Leu223 | ||
| C1FaeB2 | 5.2065 ± 0.7067 | 6.1220 | 3.969 | 10 | Leu221 | |
| 4.6880 ± 0.0000 | 4.6880 | 2.567 | 1 | Pro56 | ||
| Fae7262 | 4.8974 ± 0.1512 | 5.1510 | 4.387 | 5 | Pro56 | |
| AsFaeE | 4.7585 ± 0.1881 | 4.9000 | 2.978 | 4 | Leu59 | |
| AnFaeA | 7 | 5.8980 ± 0.6567 | 6.7200 | 5.767 | 4 | Thr68 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonopoulou, I.; Dilokpimol, A.; Iancu, L.; Mäkelä, M.R.; Varriale, S.; Cerullo, G.; Hüttner, S.; Uthoff, S.; Jütten, P.; Piechot, A.; et al. The Synthetic Potential of Fungal Feruloyl Esterases: A Correlation with Current Classification Systems and Predicted Structural Properties. Catalysts 2018, 8, 242. https://doi.org/10.3390/catal8060242
Antonopoulou I, Dilokpimol A, Iancu L, Mäkelä MR, Varriale S, Cerullo G, Hüttner S, Uthoff S, Jütten P, Piechot A, et al. The Synthetic Potential of Fungal Feruloyl Esterases: A Correlation with Current Classification Systems and Predicted Structural Properties. Catalysts. 2018; 8(6):242. https://doi.org/10.3390/catal8060242
Chicago/Turabian StyleAntonopoulou, Io, Adiphol Dilokpimol, Laura Iancu, Miia R. Mäkelä, Simona Varriale, Gabriella Cerullo, Silvia Hüttner, Stefan Uthoff, Peter Jütten, Alexander Piechot, and et al. 2018. "The Synthetic Potential of Fungal Feruloyl Esterases: A Correlation with Current Classification Systems and Predicted Structural Properties" Catalysts 8, no. 6: 242. https://doi.org/10.3390/catal8060242
APA StyleAntonopoulou, I., Dilokpimol, A., Iancu, L., Mäkelä, M. R., Varriale, S., Cerullo, G., Hüttner, S., Uthoff, S., Jütten, P., Piechot, A., Steinbüchel, A., Olsson, L., Faraco, V., Hildén, K. S., De Vries, R. P., Rova, U., & Christakopoulos, P. (2018). The Synthetic Potential of Fungal Feruloyl Esterases: A Correlation with Current Classification Systems and Predicted Structural Properties. Catalysts, 8(6), 242. https://doi.org/10.3390/catal8060242