Clog and Release, and Reverse Motions of DNA in a Nanopore


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
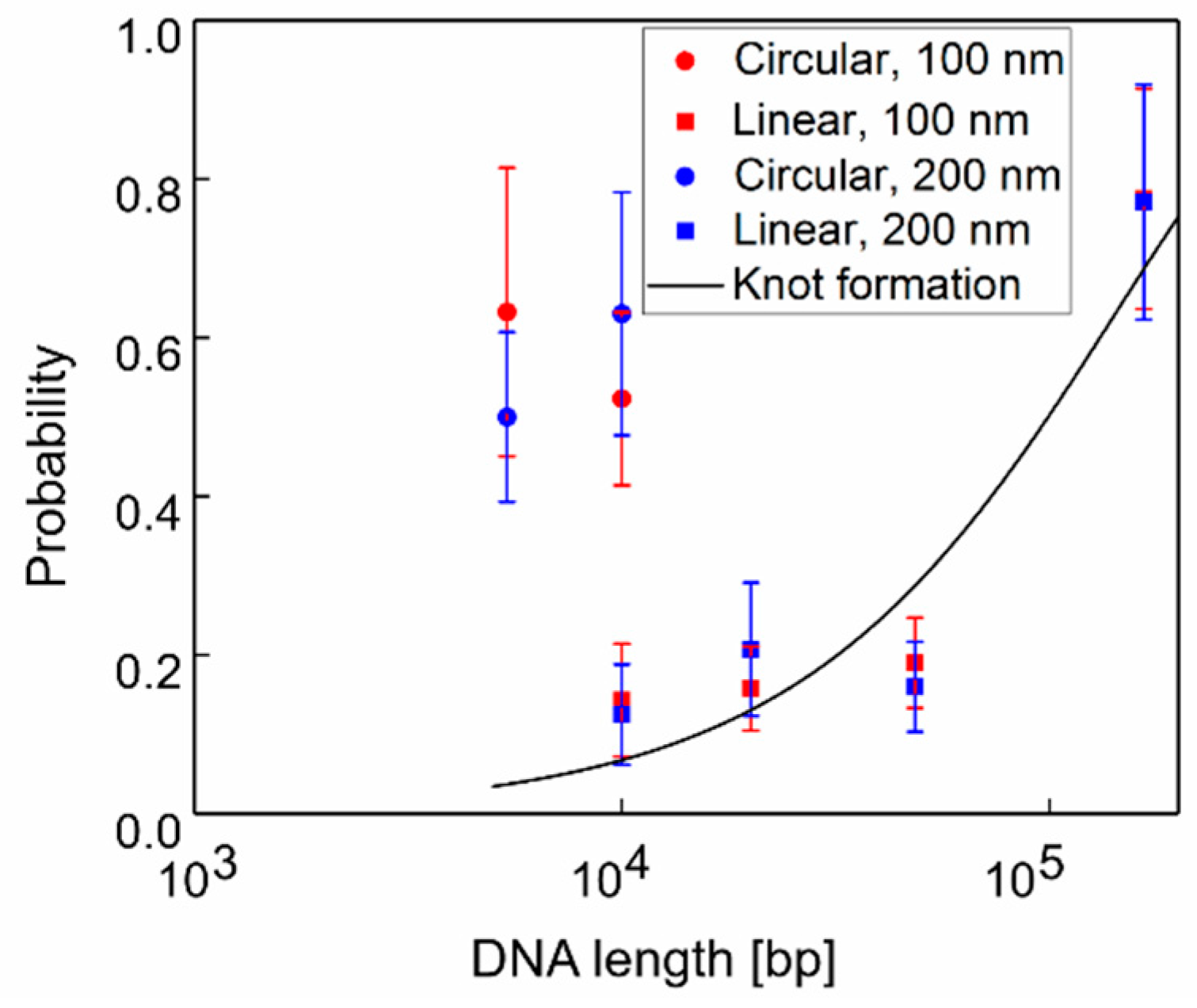
3.1. Clog Probabilities of Various DNA Length and Configurations
3.2. Release of T4 DNA to the cis Side after Clogging
3.3. U Turn of Circular DNA at Nanopore
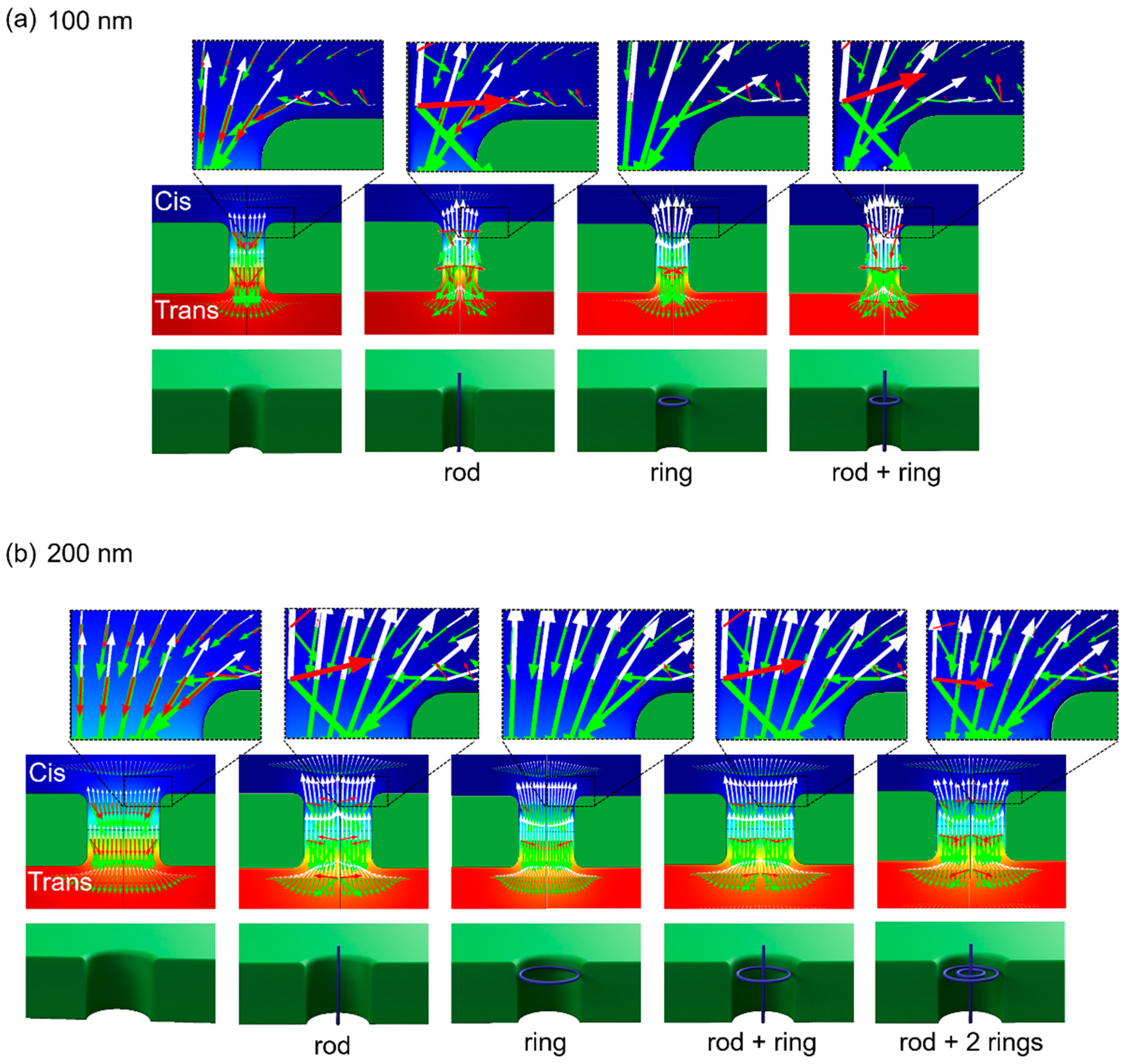
3.4. FEM-Based Numerical Estimation of DNA Motions Near Nanopore
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dekker, C. Solid-state nanopores. Nat. Nanotechnol. 2007, 2, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Branton, D.; Deamer, D.W.; Marziali, A.; Bayley, H.; Benner, S.A.; Butler, T.; Di Ventra, M.; Garaj, S.; Hibbs, A.; Huang, X.; et al. The potential and challenges of nanopore sequencing. Nat. Biotechnol. 2008, 26, 1146–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deamer, D.; Akeson, M.; Branton, D. Three decades of nanopore sequencing. Nat. Biotechnol. 2016, 34, 518–524. [Google Scholar] [CrossRef]
- Wanunu, M. Nanopores: A journey towards DNA sequencing. Phys. Life Rev. 2012, 9, 125–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyser, U.F. Controlling molecular transport through nanopores. J. R. Soc. Interface R. Soc. 2011, 8, 1369–1378. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Park, K.B.; Kim, H.J.; Yu, J.S.; Chae, H.; Kim, H.M.; Kim, K.B. Recent Progress in Solid-State Nanopores. Adv. Mater 2018, 30, e1704680. [Google Scholar] [CrossRef] [PubMed]
- Ando, G.; Hyun, C.; Li, J.; Mitsui, T. Directly observing the motion of DNA molecules near solid-state nanopores. ACS Nano 2012, 6, 10090–10097. [Google Scholar] [CrossRef]
- Fologea, D.; Ledden, B.; McNabb, D.S.; Li, J. Electrical characterization of protein molecules by a solid-state nanopore. Appl. Phys. Lett. 2007, 91, 539011–539013. [Google Scholar] [CrossRef]
- Li, J.; Gershow, M.; Stein, D.; Brandin, E.; Golovchenko, J.A. DNA molecules and configurations in a solid-state nanopore microscope. Nat. Mater. 2003, 2, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Storm, A.J.; Storm, C.; Chen, J.H.; Zandbergen, H.; Joanny, J.F.; Dekker, C. Fast DNA translocation through a solid-state nanopore. Nano Lett. 2005, 5, 1193–1197. [Google Scholar] [CrossRef]
- Larkin, J.; Henley, R.Y.; Muthukumar, M.; Rosenstein, J.K.; Wanunu, M. High-bandwidth protein analysis using solid-state nanopores. Biophys. J. 2014, 106, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Talaga, D.S.; Li, J. Single-molecule protein unfolding in solid state nanopores. J. Am. Chem. Soc. 2009, 131, 9287–9297. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, M.; Rahong, S.; Iizumi, Y.; Okazaki, T.; Taniguchi, M.; Kawai, T. Single-molecule sensing electrode embedded in-plane nanopore. Sci. Rep. 2011, 1, 46. [Google Scholar] [CrossRef] [PubMed]
- Garaj, S.; Hubbard, W.; Reina, A.; Kong, J.; Branton, D.; Golovchenko, J.A. Graphene as a subnanometre trans-electrode membrane. Nature 2010, 467, 190–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusko, E.C.; Johnson, J.M.; Majd, S.; Prangkio, P.; Rollings, R.C.; Li, J.; Yang, J.; Mayer, M. Controlling protein translocation through nanopores with bio-inspired fluid walls. Nat. Nanotechnol. 2011, 6, 253–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, A.P.; Schurmann, G.; Mondin, G.; Bitterli, R.A.; Hegelbach, N.G.; de Rooij, N.F.; Staufer, U. Sensing protein molecules using nanofabricated pores. Appl. Phys. Lett. 2006, 88, 093901. [Google Scholar] [CrossRef]
- McMullen, A.; de Haan, H.W.; Tang, J.X.; Stein, D. Stiff filamentous virus translocations through solid-state nanopores. Nat. Commun. 2014, 5, 4171. [Google Scholar] [CrossRef] [Green Version]
- Arima, A.; Tsutsui, M.; Harlisa, I.H.; Yoshida, T.; Tanaka, M.; Yokota, K.; Tonomura, W.; Taniguchi, M.; Okochi, M.; Washio, T.; et al. Selective detections of single-viruses using solid-state nanopores. Sci. Rep. 2018, 8, 16305. [Google Scholar] [CrossRef]
- Ryuzaki, S.; Tsutsui, M.; He, Y.; Yokota, K.; Arima, A.; Morikawa, T.; Taniguchi, M.; Kawai, T. Rapid structural analysis of nanomaterials in aqueous solutions. Nanotechnology 2017, 28, 155501. [Google Scholar] [CrossRef] [Green Version]
- Bell, N.A.W.; Chen, K.; Ghosal, S.; Ricci, M.; Keyser, U.F. Asymmetric dynamics of DNA entering and exiting a strongly confining nanopore. Nat. Commun. 2017, 8, 380. [Google Scholar] [CrossRef]
- Squires, T.M.; Quake, S.R. Microfluidics: Fluid physics at the nanoliter scale. Rev. Mod. Phys. 2005, 77, 977–1026. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.T.; Muthukumar, M. Polymer capture by electro-osmotic flow of oppositely charged nanopores. J. Chem. Phys. 2007, 126, 164903. [Google Scholar] [CrossRef] [PubMed]
- van der Heyden, F.H.; Bonthuis, D.J.; Stein, D.; Meyer, C.; Dekker, C. Electrokinetic energy conversion efficiency in nanofluidic channels. Nano Lett. 2006, 6, 2232–2237. [Google Scholar] [CrossRef] [PubMed]
- Laohakunakorn, N.; Ghosal, S.; Otto, O.; Misiunas, K.; Keyser, U.F. DNA interactions in crowded nanopores. Nano Lett. 2013, 13, 2798–2802. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Gu, J.; Brandin, E.; Kim, Y.R.; Wang, Q.; Branton, D. Probing Single DNA Molecule Transport Using Fabricated Nanopores. Nano Lett. 2004, 4, 2293–2298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihovilovic, M.; Hagerty, N.; Stein, D. Statistics of DNA capture by a solid-state nanopore. Phys. Rev. Lett. 2013, 110, 028102. [Google Scholar] [CrossRef] [PubMed]
- Plesa, C.; Verschueren, D.; Pud, S.; van der Torre, J.; Ruitenberg, J.W.; Witteveen, M.J.; Jonsson, M.P.; Grosberg, A.Y.; Rabin, Y.; Dekker, C. Direct observation of DNA knots using a solid-state nanopore. Nat. Nanotechnol. 2016, 11, 1093–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, A.; Di Ventra, M.; Micheletti, C. Topological jamming of spontaneously knotted polyelectrolyte chains driven through a nanopore. Phys. Rev. Lett. 2012, 109, 118301. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Kato, Y.; Ishida, K.; Hyun, C.; Li, J.; Mitsui, T. DNA motion induced by electrokinetic flow near an Au coated nanopore surface as voltage controlled gate. Nanotechnology 2015, 26, 065502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Sakashita, N.; Ishida, K.; Mitsui, T. Gate-Voltage-Controlled Threading DNA into Transistor Nanopores. J. Phys. Chem. B 2018, 122, 827–833. [Google Scholar] [CrossRef]
- Gunther, K.; Mertig, M.; Seidel, R. Mechanical and structural properties of YOYO-1 complexed DNA. Nucleic Acids Res. 2010, 38, 6526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, F.; van Kan, J.A.; van der Maarel, J.R. Effects of electrostatic screening on the conformation of single DNA molecules confined in a nanochannel. J. Chem. Phys. 2008, 128, 225109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundukad, B.; Yan, J.; Doyle, P.S. Effect of YOYO-1 on the mechanical properties of DNA. Soft Matter 2014, 10, 9721–9728. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Ghosal, S.; Hu, G. Hydrodynamic flow in the vicinity of a nanopore induced by an applied voltage. Nanotechnology 2013, 24, 245202. [Google Scholar] [CrossRef] [Green Version]
- Constantin, D.; Siwy, Z.S. Poisson-Nernst-Planck model of ion current rectification through a nanofluidic diode. Phys. Rev. E Stat. Nonline Soft Matter Phys. 2007, 76, 041202. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Tsutsui, M.; Fan, C.; Taniguchi, M.; Kawai, T. Controlling DNA translocation through gate modulation of nanopore wall surface charges. ACS Nano 2011, 5, 5509–5518. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Balducci, A.; Doyle, P.S. Ionic effects on the equilibrium dynamics of DNA confined in nanoslits. Nano Lett. 2008, 8, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, K.D. DNA electrophoresis in microfabricated devices. Rev. Mod. Phys. 2010, 82, 2903–2947. [Google Scholar] [CrossRef]
- Gershow, M.; Golovchenko, J.A. Recapturing and trapping single molecules with a solid-state nanopore. Nat. Nanotechnol. 2007, 2, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Nakane, J.; Akeson, M.; Marziali, A. Evaluation of nanopores as candidates for electronic analyte detection. Electrophoresis 2002, 23, 2592–2601. [Google Scholar] [CrossRef]
- Dai, L.; Renner, C.B.; Doyle, P.S. The polymer physics of single DNA confined in nanochannels. Adv. Colloid Interface Sci. 2016, 232, 80–100. [Google Scholar] [CrossRef] [PubMed]
- Vlassarev, D.M.; Golovchenko, J.A. Trapping DNA near a solid-state nanopore. Biophys. J. 2012, 103, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Conlisk, A.T. Forces affecting double-stranded DNA translocation through synthetic nanopores. Biomed. Microdevices 2011, 13, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.; Deurvorst, Z.; van der Heyden, F.H.; Koopmans, W.J.; Gabel, A.; Dekker, C. Electrokinetic concentration of DNA polymers in nanofluidic channels. Nano Lett. 2010, 10, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Mitscha-Baude, G.; Buttinger-Kreuzhuber, A.; Tulzer, G.; Heitzinger, C. Adaptive and iterative methods for simulations of nanopores with the PNP–Stokes equations. J. Comput. Phys. 2017, 338, 452–476. [Google Scholar] [CrossRef] [Green Version]
- Buyukdagli, S.; Ala-Nissila, T. Controlling Polymer Translocation and Ion Transport via Charge Correlations. Langmuir 2014, 30, 12907–12915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suma, A.; Micheletti, C. Pore translocation of knotted DNA rings. Proc. Natl. Acad. Sci. USA 2017, 114, E2991–E2997. [Google Scholar] [CrossRef] [PubMed]
- Krueger, E.; Shim, J.; Fathizadeh, A.; Chang, A.N.; Subei, B.; Yocham, K.M.; Davis, P.H.; Graugnard, E.; Khalili-Araghi, F.; Bashir, R.; et al. Modeling and Analysis of Intercalant Effects on Circular DNA Conformation. ACS Nano 2016, 10, 8910–8917. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubota, T.; Lloyd, K.; Sakashita, N.; Minato, S.; Ishida, K.; Mitsui, T. Clog and Release, and Reverse Motions of DNA in a Nanopore. Polymers 2019, 11, 84. https://doi.org/10.3390/polym11010084
Kubota T, Lloyd K, Sakashita N, Minato S, Ishida K, Mitsui T. Clog and Release, and Reverse Motions of DNA in a Nanopore. Polymers. 2019; 11(1):84. https://doi.org/10.3390/polym11010084
Chicago/Turabian StyleKubota, Tomoya, Kento Lloyd, Naoto Sakashita, Seiya Minato, Kentaro Ishida, and Toshiyuki Mitsui. 2019. "Clog and Release, and Reverse Motions of DNA in a Nanopore" Polymers 11, no. 1: 84. https://doi.org/10.3390/polym11010084
APA StyleKubota, T., Lloyd, K., Sakashita, N., Minato, S., Ishida, K., & Mitsui, T. (2019). Clog and Release, and Reverse Motions of DNA in a Nanopore. Polymers, 11(1), 84. https://doi.org/10.3390/polym11010084